
Morphological and Molecular Characterization of Prevalent Plant-Parasitic Nematodes from Turfgrasses in Guangdong, China

Abstract
:1. Introduction
2. Material and Methods
2.1. Soil Sampling
2.2. Morphological Characterization
2.3. Molecular Characterization
3. Results
3.1. Nematode Species
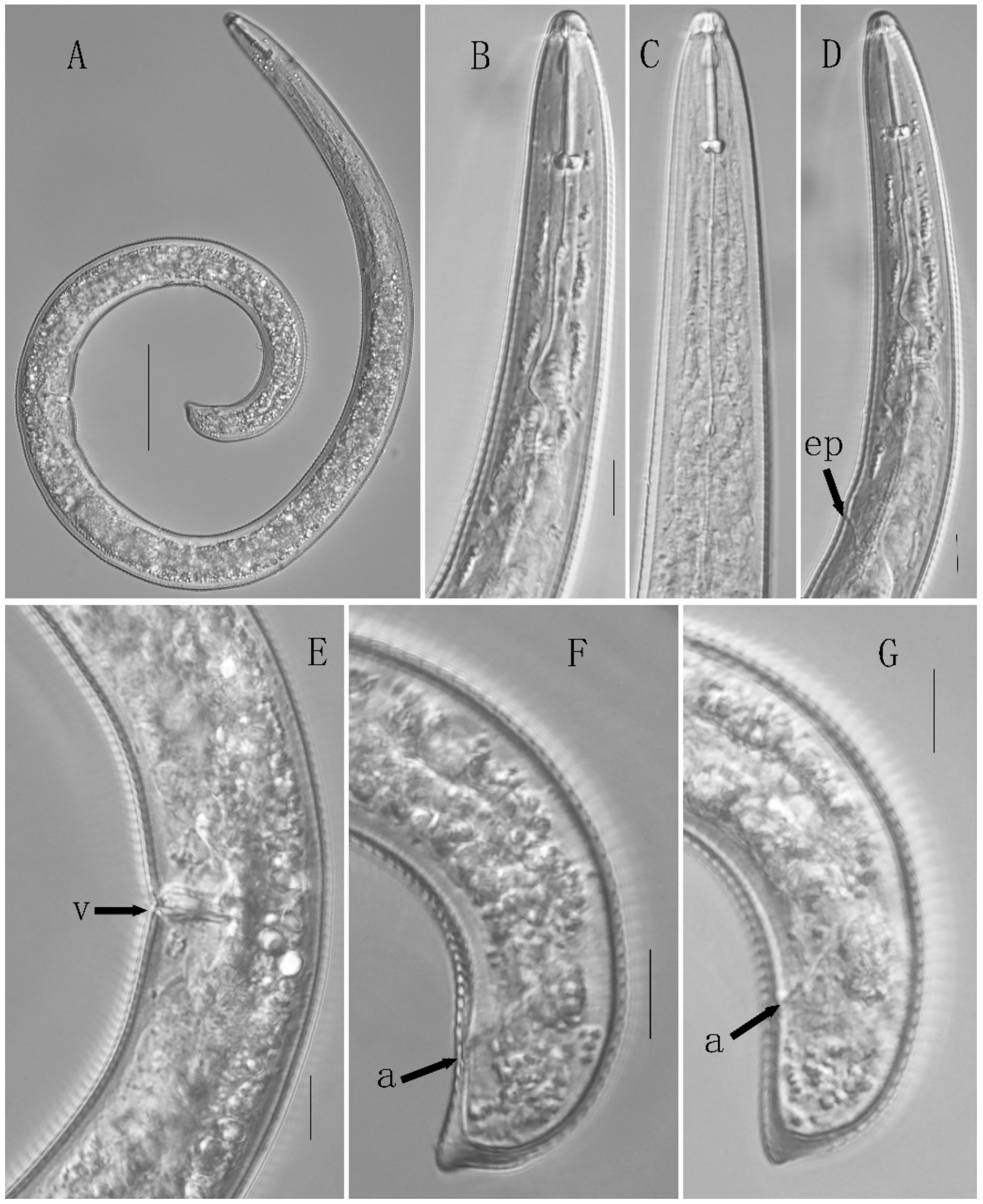
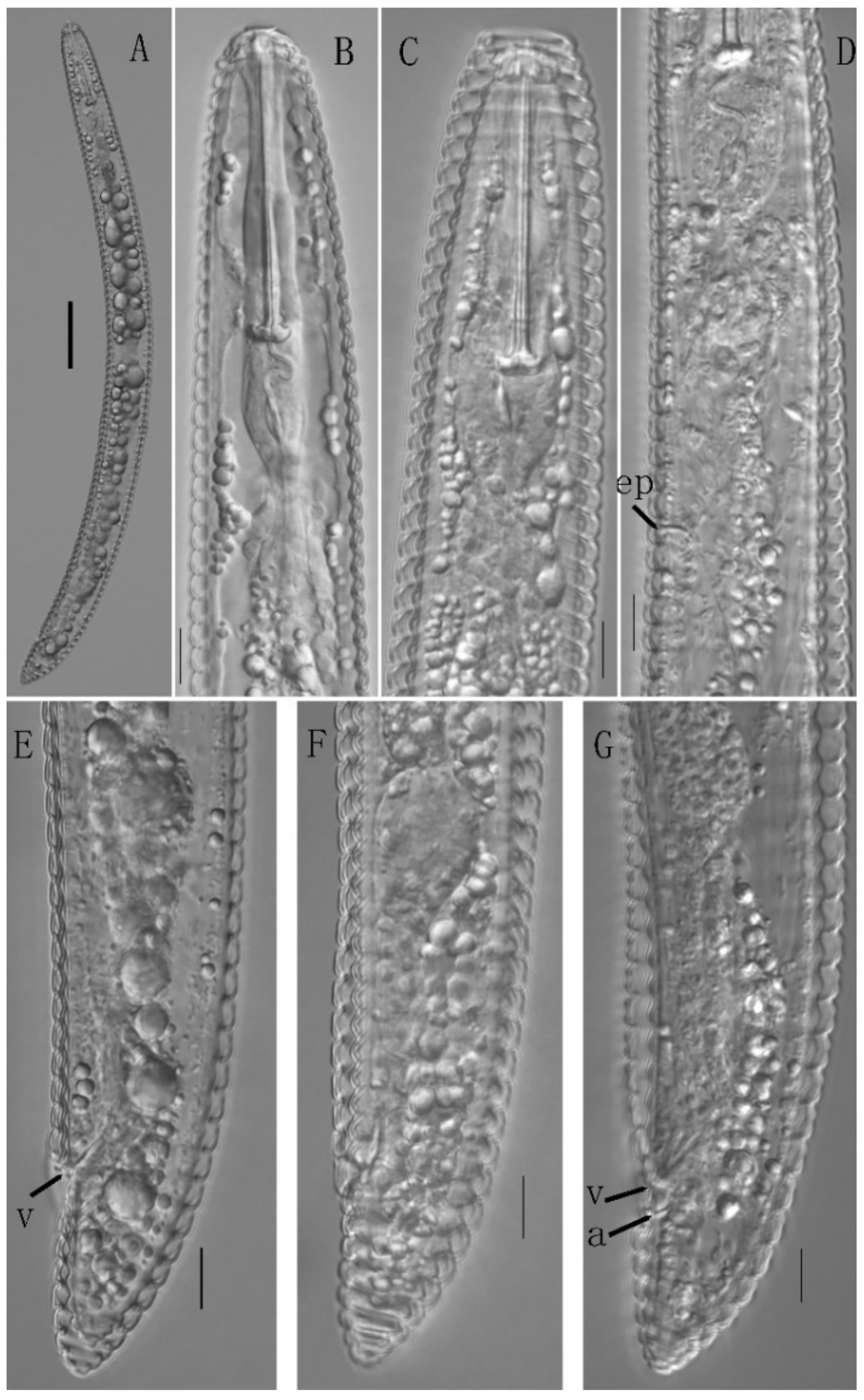
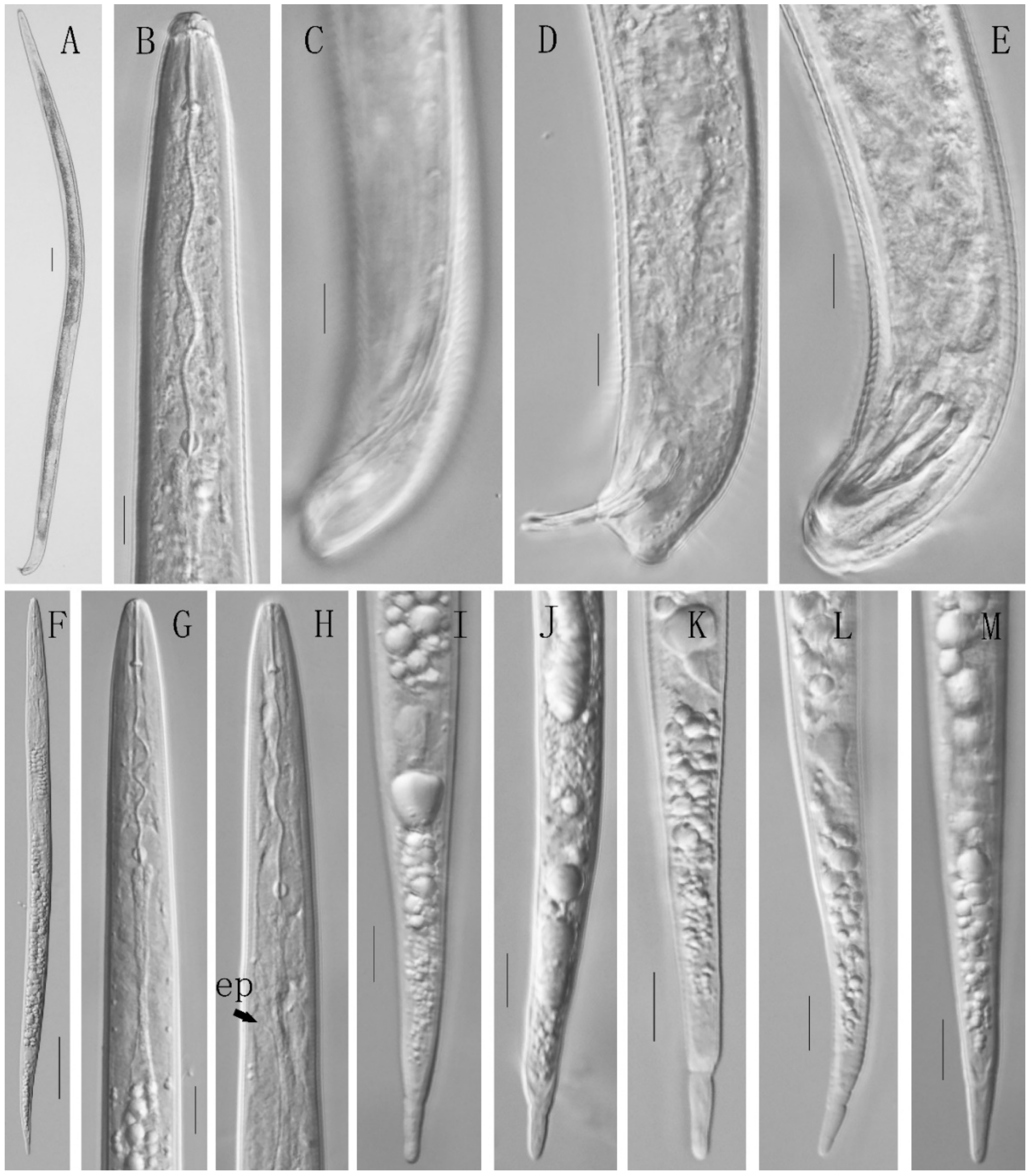
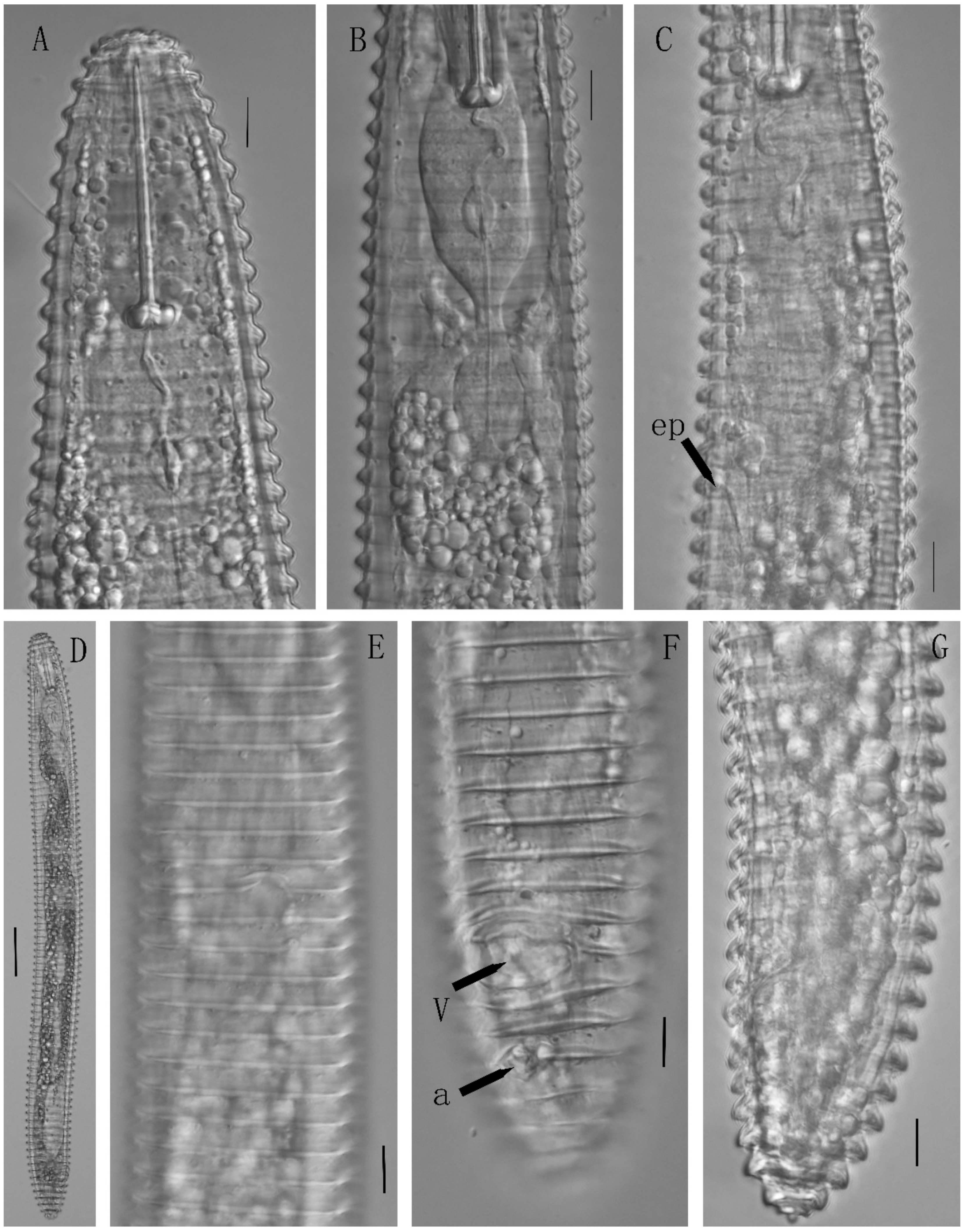
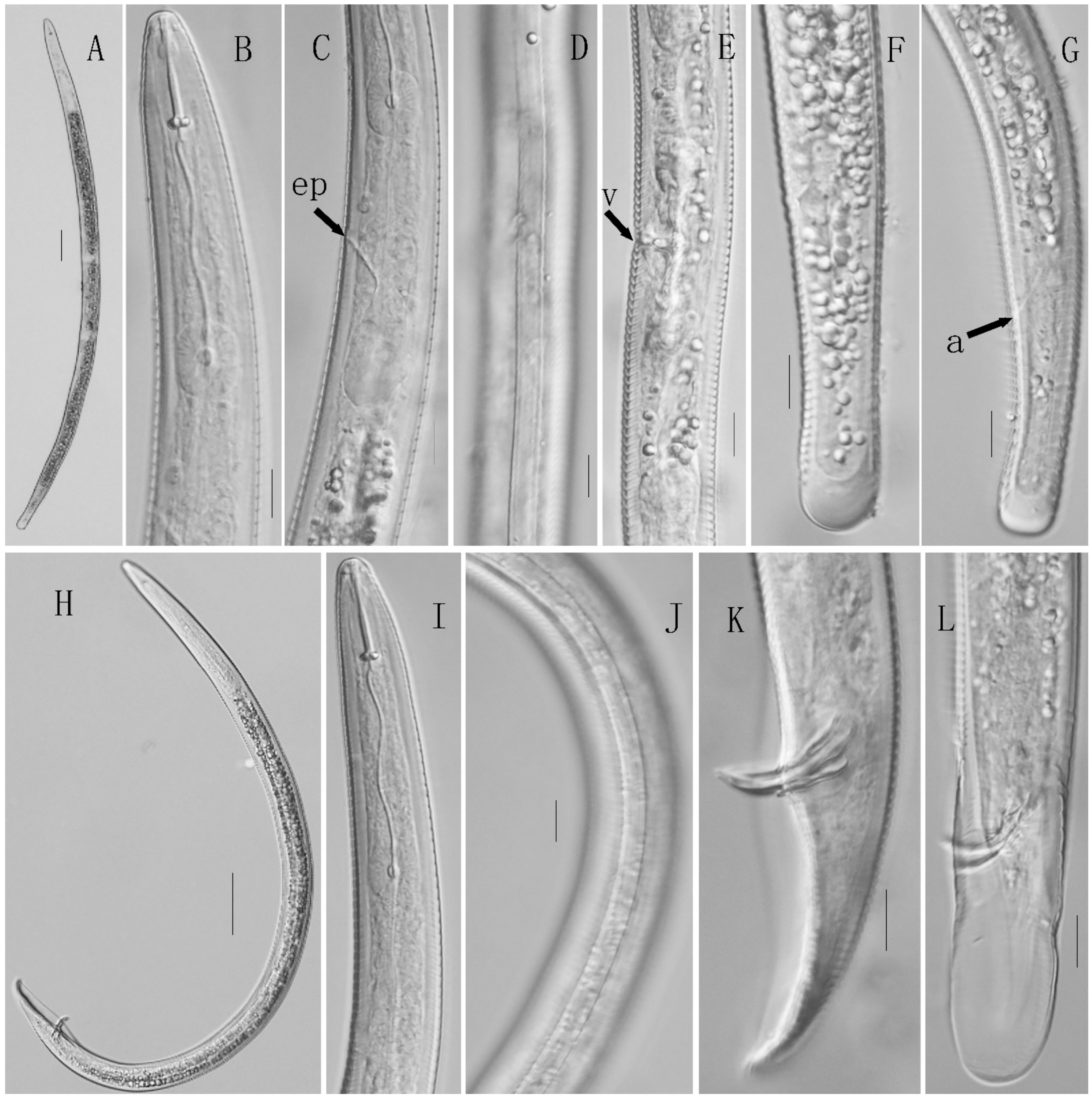
3.2. Morphological Description
3.2.1. Description of Helicotylenchus dihystera
3.2.2. Description of Hemicriconemoides rosae
3.2.3. Description of Meloidogyne graminis
3.2.4. Description of Microcinema xenoplax
3.2.5. Description of Tylenchorhynchus leviterminalis
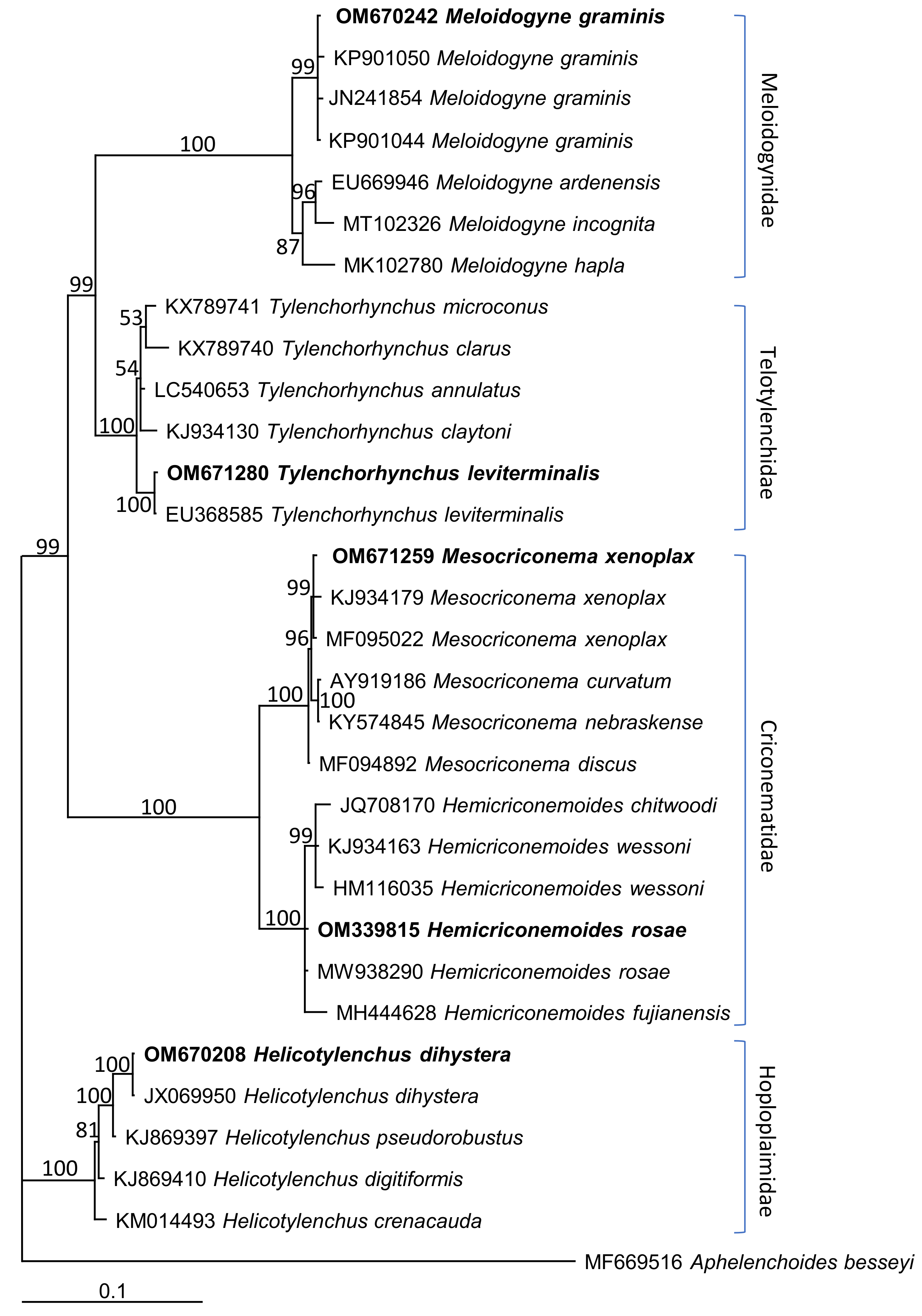
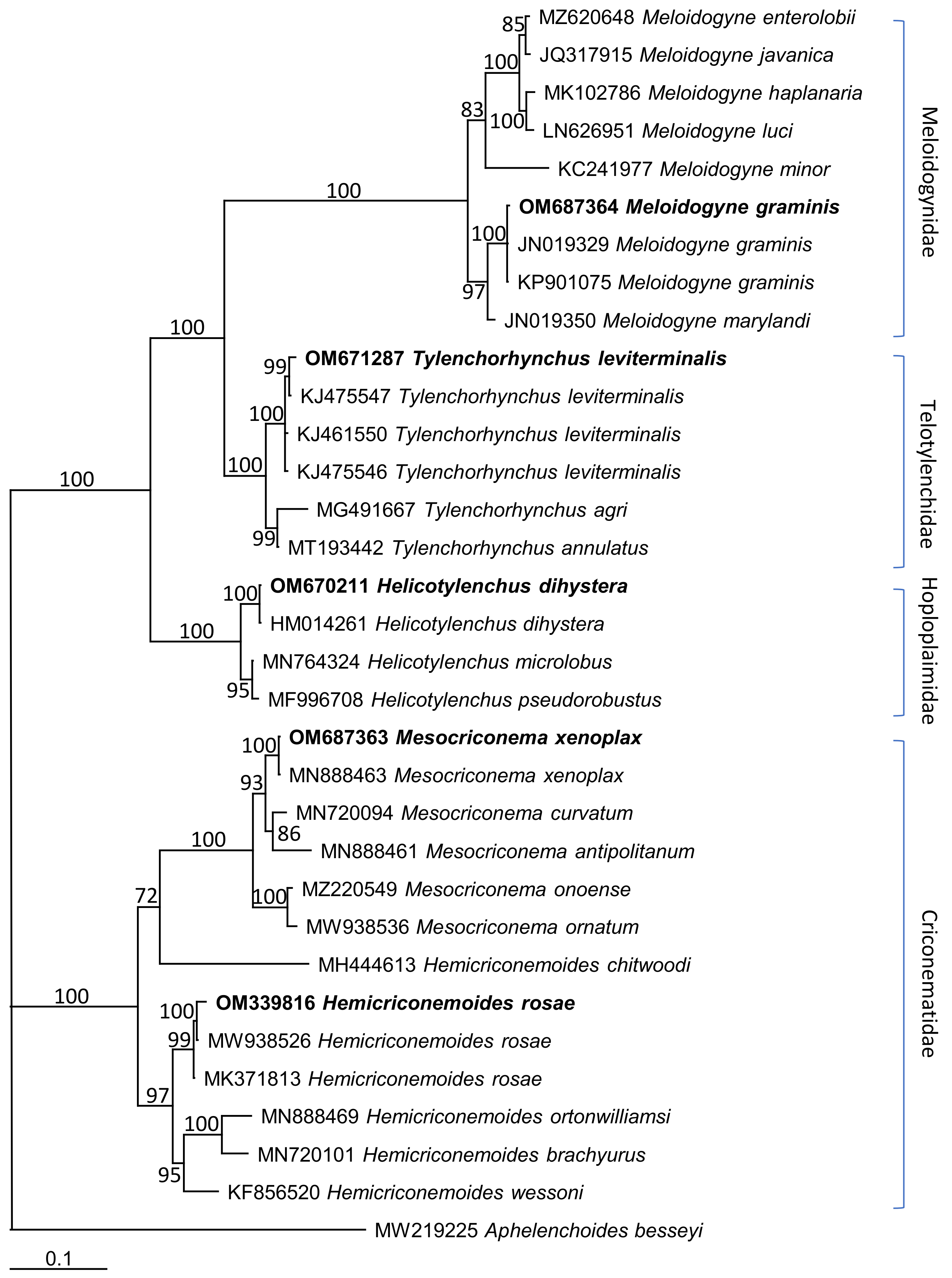
3.3. Molecular Characterization and Phylogenetic Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinese National Bureau of Statistics. The Top Ten Provinces in the Country in Terms of Urban Green Area Rankings. Available online: https://www.ixigua.com/6921648720102556164?wid_try=1 (accessed on 25 January 2021).
- Hong’e, M. Golf Courses Proliferate in China, Defying Gov’t Ban. Available online: https://lawrencesolomon.files.wordpress.com/2011/10/prolif.pdf (accessed on 22 May 2011).
- Walker, N.R.; Goad, C.L.; Zhang, H.; Martin, D.L. Factors associated with populations of plant-parasitic nematodes in bentgrass putting greens in Oklahoma. Plant Dis. 2002, 86, 764–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, Y.K.; Starr, J.L. Nematodes in Texas Golf Course. Available online: https://cdn-ext.agnet.tamu.edu/wp-content/uploads/2018/09/E-294-nematodes-in-texas-golf-courses.pdf (accessed on 30 September 2018).
- Crow, W. Plant parasitic nematodes on golf course turf. Pest Manag. 2005, 12, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Mwamula, A.O.; Lee, D.W. Occurrence of plant-parasitic nematodes of turfgrass in Korea. Plant Pathol. J. 2021, 37, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Arce, O.; Palacios-Espinosa, A.; Carillo-Fasio, J.; Hernandez-Montiel, L.G.; Hernández-Rubio, J.S.; Romero, M. Diversity and distribution of plant-parasitic nematodes on golf courses in localities of Baja California Sur, Mexico. Rev. Fac. Agron. La Univ. Del Zulia 2021, 38, 651–660. [Google Scholar]
- Vandenbossche, B.; Viaene, N.; De Sutter, N.; Maes, M.; Kareeen, G.; Bert, W. Diversity and incidence of plant-parasitic nematodes in Belgian turf grass. Nematology 2011, 13, 245–256. [Google Scholar] [CrossRef]
- Yu, Q.; Potter, J.W.; Gilby, G. Plant-parasitic nematodes associated with turfgrass in golf courses in southern Ontario. Can. J. Plant Pathol. 1998, 20, 304–307. [Google Scholar] [CrossRef]
- Zeng, Y.; Ye, W.; Tredway, L.; Martin, S.; Martin, M. Taxonomy and morphology of plant-parasitic nematodes associated with turfgrasses in North and South Carolina, USA. Zootaxa 2012, 3452, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Ye, W.; Martin, S.; Martin, M.; Tredway, L. Diversity and occurrence of nematodes associated with turfgrasses in North and South Carolina, USA. J. Nematol. 2012, 44, 345–355. [Google Scholar]
- Zeng, Y.; Ye, W.; Kerns, J.; Tredway, L.; Martin, S.; Martin, M. Molecular characterization and phylogenetic relationships of plant-parasitic nematodes associated with turfgrasses in North Carolina and South Carolina, United States. Plant Dis. 2015, 99, 982–993. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, K.; Hu, M.; Wang, H.; Tang, Z.; Shao, X.; Liao, J. Identification of Meloidogyne graminis on golf greens. Acta Pratacul. Sin. 2011, 20, 253–256. (In Chinese) [Google Scholar]
- Jenkins, W.R. A rapid centrifugal-flotation technique for separating nematodes from soil. Plant Dis. Rep. 1964, 48, 692. [Google Scholar]
- Hooper, D.J. Handling, fixing, staining and mounting nematodes. In Laboratory Methods for Work with Plant and Soil Nematodes, 5th ed.; Her Majesty’s Stationery Office: London, UK, 1970; pp. 39–54. [Google Scholar]
- Ye, W. Applying Microsoft Works spread sheet in statistics for morphometric data of nematode identification. Afro-Asian J. Nematol. 1996, 6, 203–211. [Google Scholar]
- Williams, B.D.; Schrank, B.; Huynh, C.; Shownkeen, R.; Waterston, R.H. A genetic mapping system in Caenorhabditis elegans based on polymorphic sequence—Tagged sites. Genetics 1992, 131, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Mullin, P.G.; Harris, T.S.; Powers, T.O. Phylogenetic relationships of Nygolaimina and Dorylaimina (Nematoda: Dorylaimida) inferred from small subunit ribosomal DNA sequences. Nematology 2005, 7, 59–79. [Google Scholar] [CrossRef]
- Nunn, G.B. Nematode Molecular Evolution. Ph.D. Dissertation, University of Nottingham, Nottingham, UK, 1992. [Google Scholar]
- Ye, W.; Giblin-Davis, R.M.; Braasch, H.; Morris, K.; Thomas, W.K. Phylogenetic relationships among Bursaphelenchus species (Nematoda: Parasitaphelenchidae) inferred from nuclear ribosomal and mitochondrial DNA sequence data. Mol. Phylogenetics Evol. 2007, 43, 1185–1197. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetic analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. Mr Bayes: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 1754–1755. [Google Scholar] [CrossRef] [Green Version]
- Larget, B.; Simon, D.L. Markov chain Monte Carlo algorithms for the Bayesian analysis of phylogenetic trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef]
- CABI. Helicotylenchus dihystera. [Distribution map]. In Distribution Maps of Plant Diseases (April), 1st ed.; CABI: Wallingford, UK, 2010. [Google Scholar]
- Kinloch, R.A. Florida field crops as hosts of Helicotylenchus dihystera. Nematropica 1971, 1, 38–39. [Google Scholar]
- Siddiqi, M.R. Helicotylenchus dihystera; Commonwealth Institute of Helminthology Descriptions of Plant-Parasitic Nematodes, Commonwealth Agricultural Bureaux: St. Albans, UK, 1972. [Google Scholar]
- Lucas, L.T.; Blake, C.T.; Barker, K.R. Nematodes associated with bentgrass and bermudagrass golf greens in North Carolina. Plant Dis. Rep. 1974, 58, 822–824. [Google Scholar]
- Sumner, D.R. Nematodes in bluegrass. Plant Dis. Rep. 1967, 51, 457–460. [Google Scholar]
- Rathour, K.S.; Sharma, S.; Singh, K.; Ganguly, S. Three new species of Hemicriconemoides (Nematoda: Tylenchida: Criconematina) associated with ornamental plants in Bareilly district of Uttar Pradesh, India. Indian J. Nematol. 2003, 33, 160–166. [Google Scholar]
- Khan, M.; Phani, V.; Chauhan, K.; Somvanshi, V.S.; Pervez, R.; Walia, R.K. Redescription and molecular characterisation of Hemicriconemoides rosae Rathour, Sharma, Singh & Ganguly, 2003 from rhizosphere of sugarcane in India. Nematology 2019, 21, 767–778. [Google Scholar]
- Sledge, E.B. Preliminary report on a Meloidogyne sp. parasite of grass in Florida. Plant Dis. Rep. 1962, 46, 52–54. [Google Scholar]
- Dickson, O.J. Some observations on Hypsoperine graminis in Kansas. Plant Dis. Rep. 1966, 50, 396–398. [Google Scholar]
- Grisham, M.P.; Dale, J.L.; Riggs, R.D. Meloidogyne graminis and Meloidogyne spp. on zoysia; infection, reproduction, disease development, and control. Phytopathology 1974, 64, 1485–1489. [Google Scholar] [CrossRef]
- McClure, M.A.; Nischwitz, C.; Skantar, A.M.; Schmitt, M.E.; Subbotin, S.A. Root-knot nematodes in golf greens of the western United States. Plant Dis. 2012, 96, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Perichi, G. First report of Meloidogyne graminis (Nematoda: Tylenchida) in Venezuela. Fitopatol. Venez. 2006, 19, 17–18. [Google Scholar]
- Oliveira, S.A.; Oliveira, C.M.G.; Maleita, C.M.N.; Silva, M.F.A.; Abrantes, I.M.O.; Wilcken, S.R.S. First report of Meloidogyne graminis on golf courses turfgrass in Brazil. PLoS ONE 2018, 13, e0192397. [Google Scholar]
- Sturhan, D. Outdoor occurrence of Meloidogyne species in western Germany. Nachr. Dtsch. Pflanzenschutzd. 1976, 288, 113–117. [Google Scholar]
- Wesemael, W.; Viaene, N.; Moens, M. Root-knot nematodes (Meloidogyne spp.) in Europe. Nematology 2011, 13, 3–16. [Google Scholar] [CrossRef]
- Kaul, V.K.; Chhabra, H.K. A new record of Meloidogyne graminis on Raya and occurrence of Meloidogyne spp. in Ludhiana, Punjab, India. J. Oilseeds Res. 1988, 5, 200–202. [Google Scholar]
- Murray, J.J.; Poole, T.E.; Ostazeski, S.A. Techniques for determining reproduction of Meloidogyne graminis on zoysiagrass and bermudagrass. Plant Dis. 1986, 70, 559–560. [Google Scholar] [CrossRef] [Green Version]
- Sledge, E.B.; Golden, A.M. Hypsoperine graminis (Nematode: Heteroderidae), a new genus and species of plant parasitic nematode. Proc. Helminthol. Soc. Wash. 1964, 31, 83–88. [Google Scholar]
- Raski, D.J. On the morphology of Criconemoides Taylor 1936, with descriptions of six new species. Proc. Helminthol. Soc. Wash. 1952, 19, 85–99. [Google Scholar]
- Nyczepir, A.P.; Bertrand, P.F.; Miller, R.W.; Motsinger, R.E. Incidence of Criconemella spp. and peach orchard histories in short life and non-short life sites in Georgia and South Carolina. Plant Dis. 1985, 69, 874–877. [Google Scholar] [CrossRef]
- Aballay, E.; Persson, P.; Martensson, A. Plant-parasitic nematodes in Chilean vineyards. Nematropica 2009, 39, 85–97. [Google Scholar]
- De Abrantes, I.M.O.; Vieira dos Santos, M.C.; da Conceição, I.L.P.M.; de Santos, M.S.N.A.; Vovlas, N. Root-knot and other plant-parasitic nematodes associated with fig trees in Portugal. Nematol. Medit. 2008, 36, 131–136. [Google Scholar]
- Karanastasi, E.; Handoo, Z.A.; Tzortzakakis, E.A. First report of Mesocriconema xenoplax (Nematoda: Criconematidae) in Greece and first record of Viburnum sp. as a possible host for this ring nematode. Helminthologia 2008, 45, 103–105. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, E. Studies on some Criconematoidea (Nematoda) from South Africa with a description of Ogma rhombosquamatum (Mehta & Raski, 1971) Andrassy, 1979. Phytophylactica 1980, 12, 15–23. [Google Scholar]
- Stirling, G.R. Distribution of plant parasitic nematodes in South Australian vineyards. Aust. J. Exp. Agric. Animal Hus. 1976, 16, 588–591. [Google Scholar] [CrossRef]
- Loof, P.A.A.; Wouts, W.M.; Yeates, G.W. Criconematidae (Nematoda: Tylenchida) from the New Zealand region: Genera Mesocriconema, Criconema, Discocriconemella, and Hemicriconemoides. N. Z. J. Zool. 1997, 24, 123–151. [Google Scholar] [CrossRef]
- Xie, Z.; Yang, Q.; Cheng, J.; Zhang, S. Eight species of nematodes parasitized at the roots of rice. J. Fujian Agric. Forest. Univ. (Nat. Sci. Ed.) 2007, 36, 20–24. [Google Scholar]
- Gupta, N.K.; Gupta, A.K. On some plant parasitic nematodes of the genus Macroposthonia De Man, 1880 (Medinematidae: Criconematoidea) from India. Rev. Iber. Parasitol. 1981, 41, 25–41. [Google Scholar]
- Orton Williams, K.J. Macroposthonia xenoplax; Commonwealth Institute of Helminthology Descriptions of Plant-Parasitic Nematodes, Commonwealth Agricultural Bureaux: St. Albans, UK, 1972. [Google Scholar]
- Deimi, A.M.; Chitambar, J.J.; Maafi, Z.T. Nematodes associated with flowering ornamental plants in Mahallat, Iran. Nematol. Medit. 2008, 36, 115–123. [Google Scholar]
- Nyczepir, A.P. Host suitability of an endophyte-friendly tall fescue grass to Mesocriconema xenoplax and Pratylenchus vulnus. Nematropica 2011, 41, 45–51. [Google Scholar]
- Siddiqi, M.R.; Mukherjee, B.; Dasgupta, M.K. Tylenchorhynchus microconus n. sp., T. crassicaudatus leviterminalis n. subsp. and T. coffeae Siddiqi & Basir, 1959 (Nematoda: Tylenchida). Syst. Parasitol. 1982, 4, 257–262. [Google Scholar]
- Tanha Maafi, Z.; Amani, M.; Ebrahimi, N. Plant parasitic nematodes of banana plantations in Systan and Baluchestan province. In Proceedings of the 17th Iranian Plant Protection Congress, Karadj, Iran, 2–5 September 2006; 330p. [Google Scholar]
- Karani, S.H.M.; Kashi, L.; Ghaderi, R.; Karegar, A. Five Species of Tylenchidae and Dolichodoridae (Nematoda: Tylenchoidea) from Iran. J. Agric. Sci. Technol. 2015, 15, 227–240. [Google Scholar]
- Chen, D.Y.; Ni, H.F.; Yen, J.H.; Tsay, T.T. Identification of stunt nematode Tylenchorhynchus annulatus and a new recorded Tylenchorhynchus leviterminalis (Nematoda: Belonolaimidae) in Taiwan. Plant Pathol. Bull. 2006, 15, 251–262, (In Chinese with English Abstract). [Google Scholar]
- Vovlas, N.; Cheng, H. Morpho-anatomy of Tylenchorhynchus leviterminalis from the People’s Republic of China. Nematol. Medit. 1988, 16, 149–152. [Google Scholar]
- Talavera, M.; Watanabe, T.; Mizukubo, T. Description of Tylenchorhynchus shimizui n. sp. from Paraguay and notes on T. leviterminalis Siddiqi, Mukherjee & Dasgupta from Japan (Nematoda: Tylenchida: Telotylenchidae). Syst. Parasitol. 2002, 51, 171–177. [Google Scholar] [PubMed]
- Mitkowski, N.A. First report of Subanguina radicicola, the root gall nematode infecting Poa annua putting greens in Washington State. Plant Dis. 2007, 91, 905. [Google Scholar] [CrossRef]
- Karssen, G.; Bolk, R.J.; Vanaelst, A.C.; Vanden Beld, I.; Kox, L.F.F.; Korthals, G.; Molendijk, L.; Zijlstra, C.; Van Hoof, R.; Cook, R. Description of Meloidogyne minor n. sp. (Nematoda: Meloidogynidae), a root-knot nematode associated with yellow patch disease in golf courses. Nematology 2004, 6, 59–72. [Google Scholar] [CrossRef]
- Simard, L.; Bélair, G.; Miller, S. First report of Longidorus breviannulatus associated with damage on creeping bentgrass golf greens in Québec, Canada. Plant Dis. 2009, 93, 846–847. [Google Scholar] [CrossRef] [PubMed]
- Echeverría, M.M.; Chaves, E.J. Identification of Meloidogyne naasi Franklin, 1965 from Argentina. Nematologica 1998, 44, 219–220. [Google Scholar]
- Magnusson, C.; Hammeraas, B. Nematoder i grasbaner. Växtskyddsnotiser 1997, 61, 121–132. [Google Scholar]
- Knuth, P. Schaden in einem Sportsrasen durch das Würzengallenälchen Meloidogyne naasi in Baden-Württemberg. Nachr. Des Dtsch. Pflanzenschutzd. 1998, 12, 305–307. [Google Scholar]
- Oka, Y.; Karssen, G.; Mor, M. First report of the root-knot nematode Meloidogyne marylandi on turfgrasses in Israel. Plant Dis. 2004, 88, 309. [Google Scholar] [CrossRef]
- Laughlin, C.W.; Vargas, J.M., Jr. Pathogenic potential of Tylenchorhynchus dubius on selected turfgrass. J. Nematol. 1972, 4, 277–280. [Google Scholar]
- Geraert, E. The Criconematidae of the World. In Identification of the Family Criconematidae (Nematoda); Academia Press: Ghent, Belgium, 2010; 615p. [Google Scholar]
- Zhao, X. Identification and Analysis of Molecular Phylogeny of Criconematid Species in the Southern China. Master’s Thesis, Zhongkai University of Agriculture and Engineering, Guangzhou, China, 2021. (In Chinese). [Google Scholar]
- Cobb, N.A. New nematode genera found inhabiting fresh-water and non-brackish soils. J. Wash. Acad. Sci. 1913, 3, 432–445. [Google Scholar] [CrossRef]
- Handoo, Z.A. A Key and diagnostic compendium to the species of the genus Tylenchorhynchus Cobb, 1913 (Nematoda: Belonolaimidae). J. Nematol. 2000, 32, 20–34. [Google Scholar] [PubMed]
- Uzma, I.; Nasira, K.; Firoza, K.; Shahina, F. Review of the genus Helicotylenchus Steiner, 1945 (Nematoda: Hoplolaimidae) with updated diagnostic compendium. Pakistan. J. Nematol. 2015, 33, 115–160. [Google Scholar]
- Fortuner, R.; Maggenti, A.R.; Whittaker, L.M. Morphometrical variability in Helicotylenchus Steiner, 1945. 4: Study of field populations of H. pseudorobustus and related species. Rev. Nematol. 1984, 7, 121–135. [Google Scholar]
- Subbotin, S.A.; Inserra, R.N.; Marias, M.; Mullin, P.; Powers, T.O.; Roberts, P.A.; Van Den Berg, E.; Yates, G.W.; Baldwin, J.G. Diversity and phylogenetic relationships within the spiral nematodes of Helicotylenchus Steiner, 1945 (Tylenchida: Hoplolaimidae) as inferred from analysis of the D2-D3 expansion segments of 28S rRNA gene sequences. Nematology 2011, 13, 333–345. [Google Scholar]
- Brzeski, M.W.; Loof, P.A.A.; Choi, Y.E. Compendium of the genus Mesocriconema Andrássy, 1965 (Nematoda: Criconematidae). Nematology 2002, 4, 341–360. [Google Scholar]
- Powers, T.O.; Bernard, E.C.; Harris, T.; Higgins, R.; Olson, M.; Lodema, M.; Mullin, P.; Sutton, L.; Powers, K.S. COI haplotype groups in Mesocriconema (Nematoda: Criconematidae) and their morphospecies associations. Zootaxa 2014, 3827, 101–146. [Google Scholar] [CrossRef] [Green Version]
- Powers, T.O.; Mullin, P.; Higgins, R.; Harris, T.; Powers, K.S. Description of Mesocriconema ericaceum n. sp. (Nematoda: Criconematidae) and notes on other nematode species discovered in an ericaceous heath bald community in Great Smoky Mountains National Park, USA. Nematology 2016, 18, 879–903. [Google Scholar] [CrossRef] [Green Version]
- Pinkerton, J.N.; Vasconcelos, M.C.; Sampaio, T.L.; Shaffer, R.G. Reaction of grape rootstocks to ring nematode Mesocriconema xenoplax. Am. J. Enol. Vitic. 2005, 56, 377–385. [Google Scholar]
- Zeng, Y.; Roberts, J. Interactions of Root-Knot Nematodes (Meloidogyne spp.) with Mini-Ring Disease (Rhizoctonia zeae); Pee Dee Research and Education Center, Clemson University: Florence, SC, USA, 2022; manuscript in preparation. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locations | Turf Species | Locations | Turf Species |
|---|---|---|---|
| Zhuhai Cuihu Golf | Cynodon dactylon (L.) Pers. | Sand River Golf | Cynodon dactylon |
| Zhuhai Jinwan Golf | Cynodon dactylon | Guangzhou Martyrs’ Cemetery | Zoysia tenuifolia Willd. ex Trin. |
| Guangzhou Xiancun International Golf | Cynodon dactylon | Sun Yat-sen University | Eleusine indica (L.) Gaertn. Sporobolus indicus (L.) R. Br. |
| Guangzhou Lihu Golf | Cynodon dactylon | Guangzhou Baiyun Mountain | Panicum repens L. Zoysia tenuifolia |
| Guangzhou Fengshen Golf | Cynodon dactylon | Guangzhou Haizhuhu Park | Zoysia tenuifolia |
| Guangzhou Jiulonghu Golf | Cynodon dactylon | Guangzhou Liwanhu Park | Panicum repens |
| Guangzhou Nanhu Golf | Cynodon dactylon | South China Botanical Garden | Cynodon dactylon |
| Guangzhou Luhu Golf | Cynodon dactylon | Guangzhou Liuhuahu Park | Eleusine indica |
| Guangzhou Nansha Golf | Cynodon dactylon | Guangzhou People Park | Panicum repens |
| Guangzhou Lianhuashan Golf | Cynodon dactylon | Guangzhou Tianhe Park | Sporobolus indicus |
| Shunde Junan Golf | Cynodon dactylon | Guangzhou Shamian Park | Panicum repens |
| Nanhai Taoyuan Golf | Cynodon dactylon | Guangzhou Culture Park | Eleusine indica Sporobolus indicus |
| Dongguan Zhongxin Golf | Cynodon dactylon | Guangzhou Xiaogang Park | Panicum repens Zoysia tenuifolia |
| Dongguan Changan Golf | Cynodon dactylon | Guangzhou Yingzhou Ecological Park | Cynodon dactylon |
| Shenzhen Juhao Golf | Cynodon dactylon | Guangzhou Yuntai Park | Zoysia tenuifolia |
| Shenzhen Longgang Golf | Cynodon dactylon | ||
| China Zhongshan Hot Spring Golf | Cynodon dactylon | ||
| Huizhou Taojing Golf | Cynodon dactylon |
| Nematode Species | Accession Numbers | |
|---|---|---|
| 18S | 28S | |
| Helicotylenchus dihystera (Cobb) Sher | OM670208 | OM670211 |
| Hemicriconemoides rosae Rathour, Sharma, Singh & Ganguly | OM339815 | OM339816 |
| Meloidogyne graminis | OM670242 | OM687364 |
| Mesocriconema xenoplax (Raski) Loof & De Grisse | OM671259 | OM687363 |
| Tylenchorhynchus leviterminalis Siddiqi, Mukherjee & Dasgupta | OM671280 | OM671287 |
| Parasitic Nematodes | Locations | Detection Rates * (%) |
|---|---|---|
| Helicotylenchus dihystera | Zhuhai Cuihu Golf, Guangzhou Lihu Golf, South China Botanical Garden, Guangzhou Haizhuhu Park, Xiaogang Park, Yingzhou Ecological Park | 18.2 |
| Hemicriconemoides rosae | Guangzhou Lihu Golf, Culture Park, People Park, Sand River Golf, Shenzhen Juhao Golf, Dongguan Zhongxin Golf | 18.2 |
| Meloidogyne graminis | Zhuhai Cuihu Golf, Guangzhou Lihu Golf, Shunde Junan Golf, Shenzhen Juhao Golf, Dongguan Zhongxin Golf, Changan Golf, Huizhou Taojing Golf, Longgang Golf, China Zhongshan Hot Spring Golf | 27.3 |
| Mesocriconema xenoplax | Zhuhai Cuihu Golf, Zhuhai Jinwan Golf, Guangzhou Xiancun International Golf, Lihu Golf, Fengshen Golf, Jiulonghu Golf, Nanhu Golf, Luhu Golf, Nansha Golf, Lianhuashan Golf, Shunde Junan Golf, Nanhai Taoyuan Golf, Dongguan Zhongxin Golf, Dongguan Changan Golf, Shenzhen Juhao Golf, Longgang Golf, China Zhongshan Hot Spring Golf, Huizhou Taojing Golf, Tianhe Park, Guangzhou Baiyun Mountain Park | 60.6 |
| Tylenchorynchus leviterminalis | Liuhuahu Park, Liwanhu Park, Zhuhai Cuihu Golf, Guangzhou Jiulonghu Golf, Yuntai Park, Shamian Park, South China Botanical Garden | 21.2 |
| Character | Guangzhou Lihu Golf | Zhuhai Cuihu Golf | Guangzhou Haizhuhu Park | South China Botanical Garden | Guangzhou Xiaogang Park | Zeng et al. [10] |
|---|---|---|---|---|---|---|
| H. dihystera | H. dihystera | H. dihystera | H. dihystera | H. dihystera | H. dihystera | |
| n | 10 ♀♀ | 10 ♀♀ | 10 ♀♀ | 10 ♀♀ | 10 ♀♀ | 15 ♀♀ |
| L | 650.0 ± 32.1 (610.0–699.0) | 649.9 ± 28.9 (621.2–695.8) | 682.4 ±89.7 (562.3–828.2) | 648.6 ± 66.8 (527.2–762.8) | 663.5 ± 36.6 (583.5–662.6) | 649.8 ± 27.9 (620.2–693.3) |
| a | 23.8 ± 2.9 (21.3–26.0) | 22.9 ± 1.6 (19.2–24.3) | 24.3 ± 2.4 (21.9–30.8) | 25.7 ± 2.1 (21.5–28.8) | 24.5 ± 3.5 (20.0–29.3) | 21.9 ± 1.4 (19.8–23.6) |
| b | 4.5 ± 0.6 (3.8–5.3) | 4.5 ± 0.4 (4.0–5.0) | 4.4 ± 0.8 (4.1–6.7) | 5.1± 0.8 (4.8 -6.8) | 5.0 ± 0.6 (4.3–6.9) | 4.9 ± 0.2 (4.7–5.2) |
| c | 38.3 ± 4.8 (35.3–46.5) | 37.8 ± 1.9 (34.2–44.6) | 36.7 ± 6.3 (29.9–46.9) | 38.5 ± 4.3 (33.5–46.6) | 31.9 ±3.8 (27.5–36.3) | 36.1 ± 0.9 (35.2–37.2) |
| C′ | 1.1 ± 0.1 (1.0–1.2) | 1.1 ± 0.1 (1.0–1.2) | 1.4 ± 0.2 (1.2–1.6) | 1.2 ± 0.2 (1.0–1.4) | 1.2 ±0.1 (1.0–1.3) | 1.1 ± 0.1 (1.0–1.2) |
| V | 62.6 ± 0.9 (61.0–63.5) | 61.9 ± 1.5 (60.8–63.5) | 64.5 ± 1.0 (63.0–65.5) | 63.5 ± 1.2 (61.3–65.9) | 63.1 ± 2.5 (59.6–67.2) | 61.7 ± 1.7 (58.8–63.2) |
| Stylet length | 24.2 ± 1.5 (23.0–26.0) | 23.6 ± 0.5 (22.5–24.0) | 24.5 ± 1.2 (23.2–26.4) | 24.0 ± 1.0 (23.0 – 25.8) | 24.6 ±2.8 (19.9–26.8) | 23.7 ± 0.4 (23.0–24.0) |
| Excretory pore from anterior end | 106.7 ± 3.1 (102.0–109.8) | 108.0 ± 6.7 (101.5–116.5) | 111.0 ± 1.0 (110.0–112.0) | 105.3 ± 12.3 (93.8–136.2) | 109.3 ± 6.3 (101.9–116.5) | 109.0 ± 5.7 (101.0–116.6) |
| Character | Bermuda Grass Population |
|---|---|
| n | 10 ♀♀ |
| L | 477.7 ± 22.6 (434.3–509.1) |
| a | 14.8 ± 1.4 (11.2–16.2) |
| b | 4.8 ± 0.3 (4.5–5.4) |
| c | 25.0 ± 7.8 (19.9–38.7) |
| c′ | 1.0 ± 0.2 (0.8–1.2) |
| V | 93.5 ± 0.4 (92.8–94.3) |
| Body diameter | 32.6 ± 4.1 (28.5–43.5) |
| Stylet length | 52.2 ± 1.4 (49.9–54.8) |
| Pharynx length | 100.5 ± 4.9 (91.5–107.7) |
| Anal body diameter | 19.9 ± 2.5 (16.2–22.7) |
| Tail length | 20.5 ± 5.3 (12.6–25.5) |
| Excretory pore from anterior end | 135.0 ± 9.0 (125.4–144.6) |
| R | 103.8 ± 3.9 (97.0–110.0) |
| Rs | 13.6 ± 1.0 (12.0–15.0) |
| Roes | 23.8 ± 1.6 (21.0–26.0) |
| Rex | 30.0 ± 1.1 (28.0–31.0) |
| Rv | 8.8 ± 0.8 (8.0–10.0) |
| Ran | 5.6 ± 1.1 (4.0–7.0) |
| Rvan | 3.2 ± 1.3 (2.0–5.0) |
| Character | Zhuhai Cuihu Golf | Guangzhou Lihu Golf | Shunde Junan Golf | Zeng et al. [10] |
|---|---|---|---|---|
| M. graminis | M. graminis | M. graminis | M. graminis | |
| n | 15 | 15 | 15 | 15 |
| L | 410.6 ±19.8 (393.4–440.0) | 392.5 ±16.9 (378.0–422.0) | 401.9 ± 13.4 (374.6–430.6) | 392.4 ± 16.8 (368.2–420.2) |
| a | 28.0 ± 1.3 (26.5–30.0) | 26.9 ± 1.7 (22.0 -29.5) | 27.0 ± 1.1 (26.5–30.0) | 24.9 ± 1.8 (22.7–27.9) |
| b | 4.2 ± 0.3 (3.8–4.5) | 4.3 ± 0.2 (4.0–4.7) | 4.2 ± 0.4 (4.0 – 4.6) | 4.3 ± 0.2 (4.1–4.7) |
| c | 6.7 ± 0.3 (6.3–7.8) | 6.6 ± 0.5 (6.0–7.0) | 6.7 ± 0.4 (6.0–7.5) | 6.5 ± 0.4 (6.0–6.9) |
| c′ | 5.7 ± 0.3 (5.5–6.4) | 5.5 ± 0.5 (4.3–6.5) | 5.6 ± 0.3 (4.5 -6.5) | 5.5 ± 0.6 (4.8–6.4) |
| Body width | 14.5 ± 0.6 (13.0–16.2) | 15.0 ± 1.2 (13.5–17.0) | 15.0 ± 0.4 (14.5–16.0) | 15.9 ± 1.3 (13.9–17.7) |
| Stylet length | 11.8 ± 0.4 (11.0–12.5) | 11.5 ± 0.2 (11.0–12.0) | 11.0 ± 0.5 (10.0–11.8) | 11.7 ± 0.4 (11.0–12.0) |
| Body diam. at stylet basal knob | 9.5 ± 0.5 (9.0–10.0) | 9.4 ± 0.3 (9.0–10.0) | 9.6 ± 0.5 (9.0–11.0) | 9.7 ± 0.3 (9.3–10.0) |
| Pharynx length (Head to metacarpus base) | 96.0 ± 7.0 (90.0–108.5) | 90.5 ± 5.0 (81.5–96.0) | 91.8 ± 4.5 (84.0–96.0) | 90.7 ± 5.2 (81.3–96.2) |
| Anal body width | 10.8± 0.5 (9.5–11.5) | 11.0± 0.8 (10.0–12.5) | 10.6 ± 0.4 (8.5–11.8) | 11.2 ± 0.9 (10.0–12.6) |
| Tail length | 62.0 ± 5.0 (56.0–69.0) | 61.0 ± 3.5 (54.0–64.5) | 61.5 ± 1.6 (58.0–67.0) | 61.0 ± 3.8 (53.9–64.5) |
| Hyaline tail part | 11.5 ± 1.0 (10.5–13.5) | 11.2 ± 0.6 (10.0–12.5) | 11.5 ± 1.9 (10.0–12.5) | 11.5 ± 0.8 (10.2–12.6) |
| Lip width | 5.6 ± 0.5 (5.0–6.5) | 5.5 ± 0.2 (5.0–5.7) | 5.5 ± 0.6 (5.0–5.8) | 5.4 ± 0.2 (5.1–5.6) |
| Lip height | 2.5 ± 0.2 (2.3–2.8) | 2.2 ± 0.1 (2.2–2.3) | 2.4 ± 0.2 (2.2–2.6) | 2.2 ± 0.1 (2.2–2.3) |
| Metacorpus length | 10.0 ± 0.3 (9.5–10.5) | 10.0 ± 0.5 (9.0–10.7) | 10.0 ± 0.2 (9.5–10.5) | 10.0 ± 0.7 (9.1–10.8) |
| Metacorpus width | 7.6 ± 0.5 (7.0–8.5) | 7.7 ± 0.3 (7.0–8.5) | 7.5 ± 0.2 (7.0–8.0) | 7.8 ± 0.5 (7.1–8.3) |
| LipL/W | 2.2 ± 0.2 (2.0- 2.5) | 2.4 ± 0.1 (2.2–2.5) | 2.3 ± 0.3 (2.1–2.5) | 2.4 ± 0.1 (2.3–2.5) |
| Metacorpus L/W | 1.3 ± 0.1 (1.3–1.4) | 1.3 ± 0.0 (1.2–1.3) | 1.3 ± 0.1 (1.3–1.4) | 1.3 ± 0.0 (1.2–1.3 |
| H% tail | 23.0 ± 3.5 (18.0–29.0) | 20.5 ± 2.0 (17.5–22.9) | 21.0 ± 2.0 (18.5–25.5) | 18.9 ± 2.0 (16.7–21.8) |
| Character | Guangzhou Lihu Golf | Zhuhai Cuihu Golf | Guangzhou Baiyun Mountain Park | Guangzhou Tianhe Park | Zeng et al. [10] |
|---|---|---|---|---|---|
| M. xenoplax | M. xenoplax | M. xenoplax | M. xenoplax | M. xenoplax | |
| n | 10 ♀♀ | 10 ♀♀ | 10 ♀♀ | 10 ♀♀ | 15 ♀♀ |
| L | 553.9 ± 28.0 (502.9–603.5) | 542.6 ± 49.5 (455.8–622.0) | 444.4 ±51.2 (371.3–578.2) | 386.6 ± 53.8 (304.2–503.8) | 532.8 ± 44.7 (472.7–632.2) |
| a | 13.0 ± 0.6 (12.4–13.6) | 13.2 ± 0.7 (12.5–13.5) | 9.8 ± 1.4 (8.9–12.8) | 9.7 ± 1.1 (8.5–12.6) | 12.6 ± 0.9 (11.3–14.1) |
| b | 4.5 ± 0.2 (4.0–5.2) | 4.6 ± 0.2 (4.3–5.0) | 4.0 ± 0.4 (3.3–4.9) | 4.2± 0.5 (3.3–4.8) | 4.7 ± 0.3 (4.2–5.2) |
| c | 31.0 ± 4.8 (25.5–41.0) | 30.5 ± 3.5 (24.5–36.3) | 23.2 ± 2.6 (20.8–25.3) | 21.5 ± 2.1 (18.0–23.9) | 29.1 ± 3.4 (23.9–35.3) |
| c’ | 0.8 ± 0.1 (0.7–0.9) | 0.7 ± 0.1 (0.6–0.8) | 0.7 ± 0.2 (0.6–0.9) | 0.7 ± 0.1 (0.6–0.8) | 0.8 ± 0.1 (0.7–0.9) |
| V | 92.5 ± 0.5 (91.5–93.7) | 92.3 ± 0.4 (91.5–93.5) | 93.7 ± 0.6 (91.9–94.6) | 92.5 ± 0.7 (91.5–94.6) | 92.3 ± 0.4 (91.7–92.9) |
| Stylet length | 56.5 ± 1.9 (56.5–61.5) | 55.3 ± 1.5 (52.5–58.8) | 54.0 ± 2.3 (52.8–57.6) | 52.0 ± 2.1 (60.0–80.0) | 55.3 ± 1.6 (52.3–58.2) |
| Pharynx length (Head to metacarpus base) | 119.3 ± 7.4 (105.4–132.0) | 118.0 ± 6.0 (108.5–128.9) | 110.5 ± 9.5 (98.8–132.6) | 95.0 ± 5.0 (87.8–180.6) | 114.2 ± 5.1 (109.6–122.9) |
| Excretory pore from anterior end | 136.7 ± 7.5 (130.6–150.0) | 133.8 ± 9.2 (128.5–141.6) | 130.5 ± 4.7 (126.3–141.4) | 132.8 ± 5.5 (127.9–140.0) | 127.7 ± 5.0 (120.6–133.8) |
| R | 100.8 ± 3.3 (87.0–108.0) | 101.9 ± 3.1 (97.0–109.0) | 87.0 ± 3.0 (84.0–92.0) | 96.0 ± 13.0 (82.0–110.0) | 102.8 ± 4.2 (96.0–112.0) |
| Rs | 13.0 ± 0.5 (12.0–14.0) | 13.5 ± 0.4 (13.0–14.5) | 13.0 ± 1.0 (12.0–14.0) | 15.0 ± 1.0 (14.0–16.0) | 13.4 ± 0.5 (13.0–14.0) |
| Roes | 25.0 ± 1.5 (21.0–27.0) | 24.5 ± 0.9 (21.0–25.0) | 23.0 ± 2.0 (21.0–26.0) | 26.0 ± 3.0 (22.0–29.0) | 25.5 ± 0.9 (23.0–26.0) |
| Rex | 29.0 ± 0.7 (26.0–33.0) | 29.0 ± 1.5 (25.0–32.0) | 24.0 ± 2.0 (22.0–26.0) | 27.0 ± 2.0 (25.0–30.0) | 28.2 ± 1.3 (25.0–30.0) |
| Rv | 8.0 ± 0.9 (6.0–10.0) | 8.5 ± 0.7 (7.0–11.0) | 7.0 ± 1.0 (6.0–8.0) | 7.0 ± 1.0 (6.0–8.0) | 8.6 ± 0.7 (8.0–10.0) |
| Ran | 4.5 ± 1.0 (4.0–7.0) | 4.8 ± 0.5 (4.0–8.0) | 4.0 ± 1.0 (3.0–5.0) | 6.0 ± 1.0 (5.0–7.0) | 4.3 ± 0.5 (4.0–5.0) |
| Rvan | 4.0 ± 0.8 (2.0–5.0) | 4.3 ± 0.5 (3.0–5.0) | 2.5 ± 0.2 (2.0–3.0) | 2.3 ± 0.5 (2.0–3.0) | 4.3 ± 0.6 (3.0–5.0) |
| Character | Liuhuahu Park | Liwanhu Park | ||
|---|---|---|---|---|
| T. leviterminalis | T. leviterminalis | |||
| n | 10 ♀♀ | 10 ♂♂ | 10 ♀♀ | 10 ♂♂ |
| L | 642.4 ± 17.7 (620.3–661.2) | 628.6 ± 32.2 (560.2–662.2) | 667.5 ± 36.6 (606.5–722.0) | 687.5 ± 36.0 (620.5–791.0) |
| a | 29.3 ± 1.4 (27.9–30.0) | 29.7 ± 2.8 (27.5–34.8) | 27.5 ± 2.5 (25.0–31.0) | 29.5 ± 2.6 (26.5–34.0) |
| b | 4.5 ± 0.3 (4.1–5.0) | 4.0 ± 0.8 (3.0–5.1) | 4.8 ± 0.2 (4.4–5.3) | 5.5 ± 0.6 (4.9–6.5) |
| c | 14.0 ± 1.1 (12.9–16.0) | 13.5 ± 0.9 (12.5–14.6) | 15.0 ± 0.8 (13.5–16.3) | 17.0 ± 2.2 (13.1–21.3) |
| c′ | 3.5 ± 0.5 (2.8–4.3) | 2.8 ± 0.2 (2.3–3.2) | 2.6 ±0.3 (2.1–3.2) | 2.2 ± 0.3 (1.6–3.1) |
| V | 55.5 ± 0.5 (54.0–56.5) | - - | 54.5 ± 1.5 (51.0–56.2) | - - |
| Stylet length | 22.5 ± 0.5 (21.0–23.0) | 21.0 ± 6.0 (20.2–21.8) | 20.6 ± 0.8 (19.0–21.8) | 19.6 ± 0.8 (18.8–21.6) |
| MB | 48.3 ± 1.8 (46.4–51.3) | 47.5 ± 2.2 (45.1–50.1) | 48.6 ± 1.6 (46.8–50.4) | 50.6 ± 3.2 (47.1–53.8) |
| Excretory pore from anterior end | 101.0 ± 6.0 (95.0–110.5) | - - | 109.4 ± 6.6 (102.0–125.5) | - - |
| Tail annules | 16.0 ± 3.0 (12.0–20.0) | - - | 17.0 ± 2.0 (15.0–21.0) | - - |
| T | - - | 56.0 ± 3.5 (52.0–60.0) | - - | 50.5 ± 5.5 (44.0–57.5) |
| Spicule length | - - | 23.3 ± 0.8 (21.5–24.6) | - - | 22.3 ± 1.8 (20.5–25.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.; Chen, X.; Ni, Y.; Zhao, C.; Kerns, J.; Tredway, L.; Roberts, J. Morphological and Molecular Characterization of Prevalent Plant-Parasitic Nematodes from Turfgrasses in Guangdong, China. Horticulturae 2022, 8, 611. https://doi.org/10.3390/horticulturae8070611
Zeng Y, Chen X, Ni Y, Zhao C, Kerns J, Tredway L, Roberts J. Morphological and Molecular Characterization of Prevalent Plant-Parasitic Nematodes from Turfgrasses in Guangdong, China. Horticulturae. 2022; 8(7):611. https://doi.org/10.3390/horticulturae8070611
Chicago/Turabian StyleZeng, Yongsan, Xiaoguan Chen, Yibei Ni, Chen Zhao, James Kerns, Lane Tredway, and Joseph Roberts. 2022. "Morphological and Molecular Characterization of Prevalent Plant-Parasitic Nematodes from Turfgrasses in Guangdong, China" Horticulturae 8, no. 7: 611. https://doi.org/10.3390/horticulturae8070611
APA StyleZeng, Y., Chen, X., Ni, Y., Zhao, C., Kerns, J., Tredway, L., & Roberts, J. (2022). Morphological and Molecular Characterization of Prevalent Plant-Parasitic Nematodes from Turfgrasses in Guangdong, China. Horticulturae, 8(7), 611. https://doi.org/10.3390/horticulturae8070611