
Control of Brown Rot Produced by Monilinia fructicola in Peaches Using a Full-Spectrum Extract of Zuccagnia punctata Cav.

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Extract Preparation
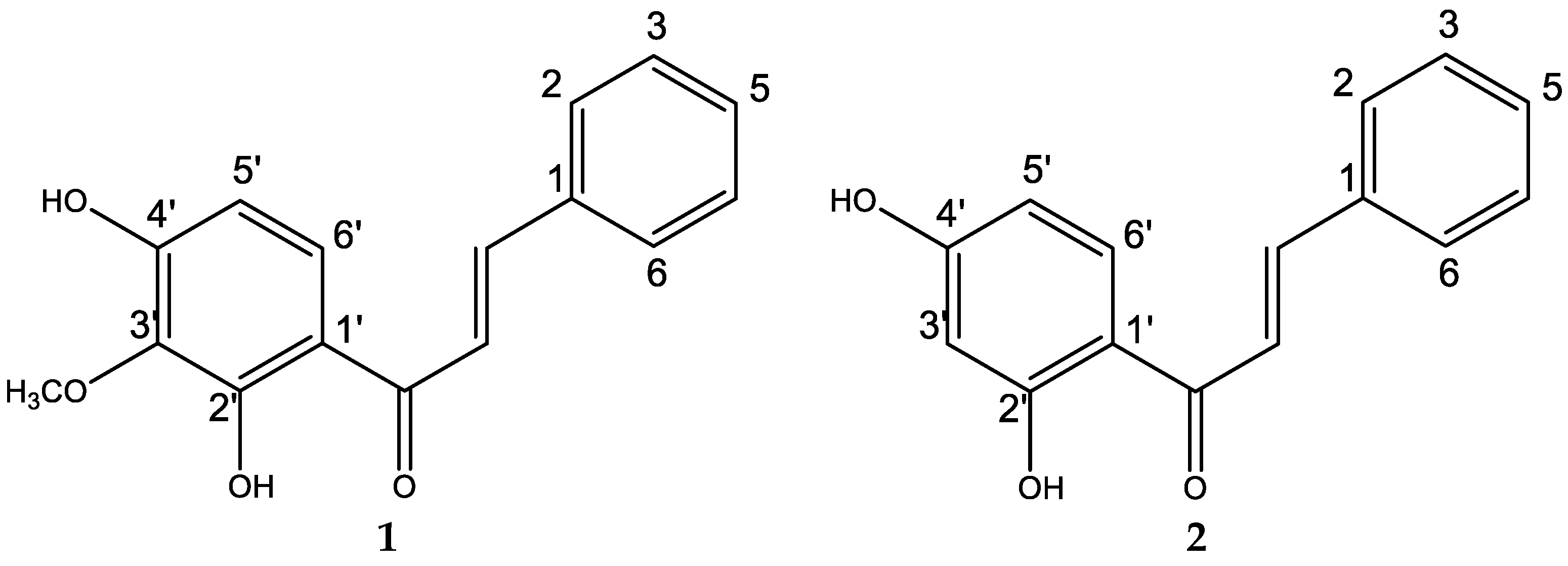
2.2. Isolation and Chemical Characterization of Natural Compounds
2.3. Antifungal Studies
2.3.1. Microorganisms and Media
2.3.2. Molecular Characterization
2.3.3. In Vitro Susceptibility Tests
2.3.4. Ex Vivo Antifungal Assay on Wounded Fruits
Peaches Collection and Preparation
Test on Peaches: Inoculation and Treatment Applications
Test Development and Evaluation
Statistical Analysis
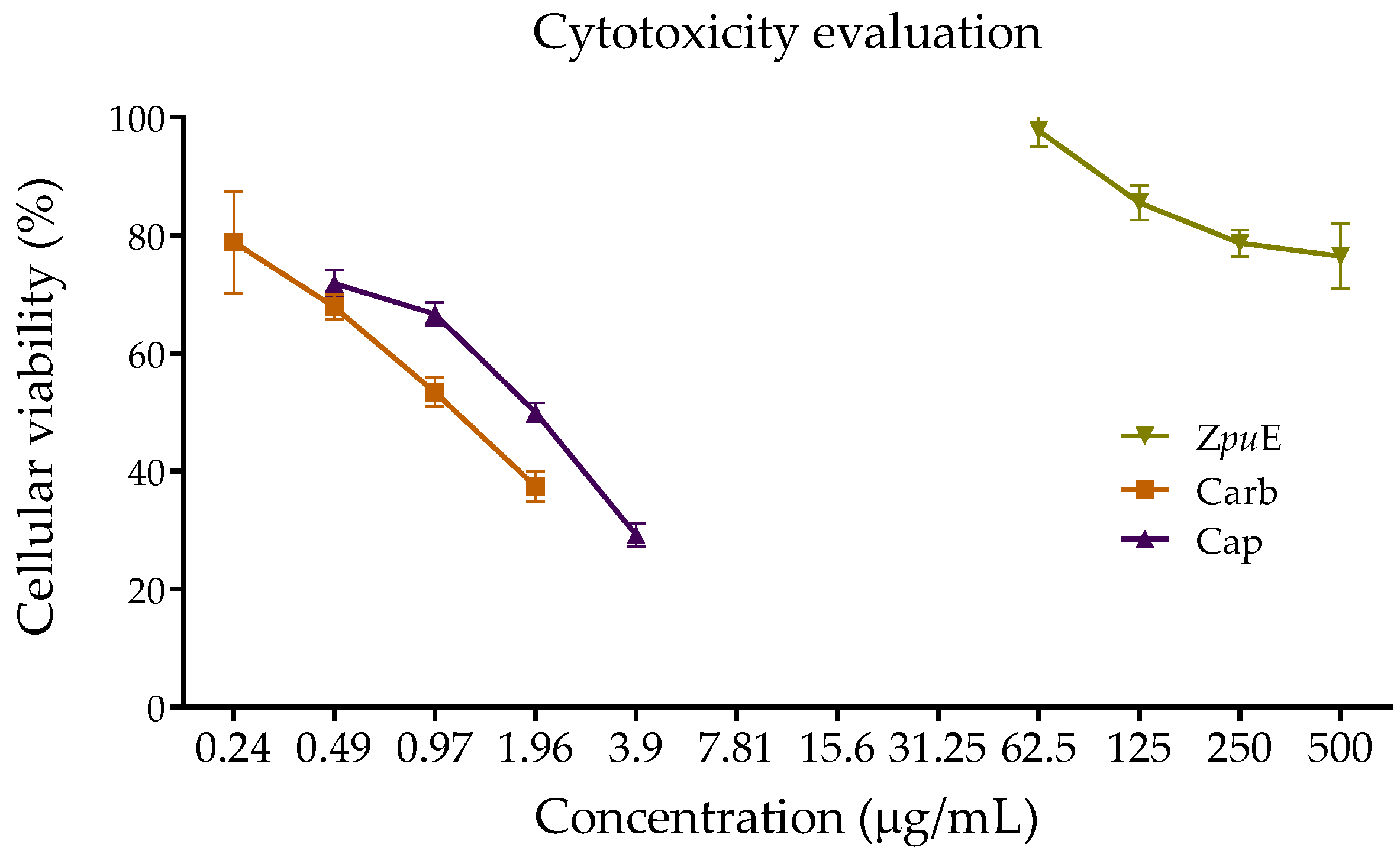
2.4. Cell Viability Assay
3. Results
3.1. Fungal Isolation and Characterization
3.2. In Vitro Antifungal Activity of ZpuE and Compounds 1 and 2 against the Selected Phytopathogenic Fungi
3.3. Detection of Bioactive Compounds by Bioautography and Identification of the Main Active Constituents in the Extract
3.4. Effect of Ethanolic Extract of Z. punctata for the Control of Peach Brown Rot on Fresh Fruits
3.5. Cytotoxicity Evaluation of Ethanolic Extract of Z. punctata Compared with Commercial Fungicides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Freimoser, F.M.; Hilber-Bodmer, M.; Brunisholz, R.; Drissner, D. Direct identification of Monilinia brown rot fungi on infected fruits by matrix-assisted laser desorption/ionization (MALDI) mass spectrometry. Chem. Biol. Technol. Agric. 2016, 3, 7. [Google Scholar] [CrossRef]
- Mondino, P. Sintomatología, etiología y características epidemiológicas de la enfermedad. In Manejo de la Podredumbre Morena (Monilinia fructicola y M. laxa) en Huertos Frutales de Uruguay, Chile, Bolivia, Brasil y Argentina; Mitidieri, M., Castillo, J.A., Eds.; CYTED Programa Interamericano de Ciencia y Tecnología para el Desarrollo: Buenos Aires, Argentina, 2014; Volume 2, pp. 35–41. [Google Scholar]
- Dowling, M.E.; Bridges, W.C.; Cox, B.M.; Sroka, T.; Wilson, J.R.; Schnabel, G. Preservation of Monilinia fructicola genotype diversity within fungal cankers. Plant Dis. 2019, 103, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Akhoon, B.A.; Gupta, S.K.; Dhar, M.K. Dissecting the genome, secretome, and effectome repertoires of Monilinia spp.: The causal agent of brown rot disease: A comparative analysis. Postharvest Biol. Technol. 2023, 195, 112120. [Google Scholar] [CrossRef]
- Casals, C.; Segarra, J.; Torres, R.; Teixidó, N.; De Cal, A.; Usall, J. Validation of a warning system to control brown rot in peach and nectarine. Agronomy 2023, 13, 254. [Google Scholar] [CrossRef]
- Mitidieri, M.; Castillo, J.A. Manejo de la Podredumbre Morena (Monilinia fructicola y M. laxa) en Huertos Frutales de Uruguay, Chile, Bolivia, Brasil y Argentina; CYTED Programa Interamericano de Ciencia y Tecnología para el Desarrollo: Buenos Aires, Argentina, 2014; Volume 2. [Google Scholar]
- Ondejková, N.; Hudecová, M.; Bacigálová, K. First report on Monilinia fructicola in the Slovak Republic. Plant Prot. Sci. 2010, 46, 181–184. [Google Scholar] [CrossRef]
- Oliveira Lino, L.; Pacheco, I.; Mercier, V.; Faoro, F.; Bassi, D.; Bornard, I.; Quilot-Turion, B. Brown rot strikes Prunus fruit: An ancient fight almost always lost. J. Agric. Food Chem. 2016, 64, 4029–4047. [Google Scholar] [CrossRef]
- Villarino, M.; Egüen, B.; Lamarca, N.; Segarra, J.; Usall, J.; Melgarejo, P.; De Cal, A. Occurrence of Monilinia laxa and M. fructigena after introduction of M. fructicola in peach orchards in Spain. Eur. J. Plant Pathol. 2013, 137, 835–845. [Google Scholar] [CrossRef]
- Rodriguez Romera, M.; Díaz Micari, M.; Pizzuolo, P. Primer reporte de Monilinia spp. en frutales de pepita en Argentina. In Libro de Resúmenes 4° Congreso Argentino de Fitopatología, 1st ed.; Asociación Argentina de Fitopatólogos, Ed.; Asociación Civil Argentina de Fitopatólogos: Córdoba, Argentina, 2017; p. 251. [Google Scholar]
- Di Liberto, M.G.; Stegmayer, M.I.; Svetaz, L.A.; Derita, M.G. Evaluation of Argentinean medicinal plants and isolation of their bioactive compounds as an alternative for the control of postharvest fruits phytopathogenic fungi. Rev. Bras. Farmacogn. 2019, 29, 686–688. [Google Scholar] [CrossRef]
- Stegmayer, M.I.; Álvarez, N.H.; Favaro, M.A.; Fernandez, L.N.; Carrizo, M.E.; Reutemann, A.G.; Derita, M.G. Argentinian wild plants as controllers of fruits phytopathogenic fungi: Trends and perspectives. In Wild Plants: The Treasure of Natural Healers; Rai, M., Bhattarai, S., Feitosa, C.M., Eds.; CRC Press: Boca Raton, FL, USA, 2020; Volume 6, pp. 121–137. [Google Scholar]
- Pergomet, J.L.; Di Liberto, M.G.; Derita, M.G.; Bracca, A.B.J.; Kaufman, T.S. Activity of the pterophyllins 2 and 4 against postharvest fruit pathogenic fungi. Comparison with a synthetic analog and related intermediates. Fitoterapia 2018, 125, 98–105. [Google Scholar] [CrossRef]
- Di Liberto, M.G.; Caldo, A.J.; Quiroga, A.D.; Riveira, M.J.; Derita, M.G. Zanthosimuline and related pyranoquinolines as antifungal agents for postharvest fruit disease control. ACS Omega 2020, 5, 7481–7487. [Google Scholar] [CrossRef]
- Cortés, I.; Di Liberto, M.G.; Kaufman, T.S.; Derita, M.G.; Bracca, A.B.J. Synthesis and evaluation of aromatic methoxime derivatives against five postharvest phytopathogenic fungi of fruits. Main structure-activity relationships. Food Chem. 2020, 321, 126701. [Google Scholar] [CrossRef] [PubMed]
- Stegmayer, M.I.; Fernandez, L.; Álvarez, N.; Seimandi, G.; Reutemann, A.; Derita, M. In Vitro Antifungal screening of argentine native or naturalized plants against the phytopathogen Monilinia fructicola. Comb. Chem. High Screen. 2022, 24, 1158–1166. [Google Scholar] [CrossRef]
- Cabrera, A.L. Fitogeografía de la República Argentina. Bol. Soc. Argent. Bot. 1971, XIV, 15–16. Available online: https://botanicaargentina.org.ar/boletin-14-1-2/ (accessed on 4 May 2023).
- Burkart, A. Las Leguminosas Argentinas, Silvestres y Cultivadas, 2nd ed.; ACME Agency: Buenos Aires, Argentina, 1952; p. 590. [Google Scholar]
- Ratera, E.L.; Ratera, M.O. Plantas de la Flora Argentina Empleadas en Medicina Popular; Hemisferio Sur: Buenos Aires, Argentina, 1980; p. 189. [Google Scholar]
- Toursarkissian, M. Plantas Medicinales de la Argentina; Sus Nombres Botánicos, Vulgares, Usos y Distribución Geográfica; Hemisferio Sur: Buenos Aires, Argentina, 1980; p. 178. [Google Scholar]
- Quiroga, E.N.; Sampietro, A.R.; Vattuone, M.A. Screening antifungal activities of selected medicinal plants. J. Ethnopharmacol. 2001, 74, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, S.L.; Cortadi, A.; Juárez, M.A.; Petenatti, E.; Tomi, F.; Casanova, J.; van Baren, C.M.; Zacchino, S.; Vila, R. (−)-5,6-Dehydrocamphor from the antifungal essential oil of Zuccagnia punctata. Phytochem. Lett. 2012, 5, 194–199. [Google Scholar] [CrossRef]
- Nuño, G.; Alberto, M.; Zampini, I.; Cuello, S.; Ordoñez, R.; Sayago, J.; Baroni, V.; Wunderlin, D.; Isla, M.I. The effect of Zuccagnia punctata Cav., an Argentina medicinal plant, on virulence factors from Candida species. Nat. Prod. Comm. 2014, 9, 933–936. [Google Scholar] [CrossRef]
- Nuño, G. Aislamiento y Caracterización de Biomoléculas Producidas por Especies Vegetales que Crecen en Ecosistemas Semiáridos. Ph.D. Thesis, Universidad Nacional de Tucumán, Tucumán, Argentina, 2015. [Google Scholar]
- Jimenez, C.M.; Sampietro, D.A.; Sgariglia, M.A.; Soberón, J.R.; Vattuone, M.A. Isolation, identification and usefulness of antifungal compounds from Zuccagnia punctata for control of toxigenic ear rot pathogens. Nat. Prod. Commun. 2014, 9, 1461–1464. [Google Scholar] [CrossRef]
- Svetaz, L.A.; Tapia, A.; López, S.N.; Furlán, R.L.; Petenatti, E.; Pioli, R.; Schmeda-Hirschmann, G.; Zacchino, S.A. Antifungal chalcones and new caffeic acid esters from Zuccagnia punctata acting against soybean infecting fungi. J. Agric. Food Chem. 2004, 52, 3297–3300. [Google Scholar] [CrossRef]
- Svetaz, L.A.; Agüero, M.B.; Alvarez, S.; Luna, L.; Feresin, G.; Derita, M.G.; Tapia, A.; Zacchino, S.A. Antifungal activity of Zuccagnia punctata Cav.: Evidence for the mechanism of action. Planta Med. 2007, 73, 1074–1080. [Google Scholar] [CrossRef]
- Svetaz, L.; Zuljan, F.; Derita, M.; Petenatti, E.; Tamayo, G.; Cáceres, A.; Cechinel Filho, V.; Giménez, A.; Pinzón, R.; Zacchino, S.A.; et al. Value of the ethnomedical information for the discovery of plants with antifungal properties. A survey among seven Latin American countries. J. Ethnopharmacol. 2010, 127, 137–158. [Google Scholar] [CrossRef]
- Agüero, M.B.; Gonzalez, M.; Lima, B.; Svetaz, L.A.; Sánchez, M.; Zacchino, S.A.; Feresin, G.E.; Schmeda Hirschmann, G.; Palermo, J.; Wunderlin, D.; et al. Argentinean propolis from Zuccagnia punctata Cav. (Caesalpinieae) exudates: Phytochemical characterization and antifungal activity. J. Agric. Food Chem. 2010, 58, 194–201. [Google Scholar] [CrossRef]
- Butassi, E.; Svetaz, L.A.; Sortino, M.A.; Quiroga, A.D.; Carvalho, V.S.D.; Cortés, J.C.C.; Ribas, J.C.; Zacchino, S.A. Approaches to the mechanism of antifungal activity of Zuccagnia punctata-Larrea nitida bi-herbal combination. Phytomedicine 2019, 54, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Butassi, E.; Svetaz, L.A.; Ivancovich, J.J.; Feresin, G.E.; Tapia, A.; Zacchino, S.A. Synergistic mutual potentiation of antifungal activity of Zuccagnia punctata Cav. and Larrea nitida Cav. extracts in clinical isolates of Candida albicans and Candida glabrata. Phytomedicine 2015, 22, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.A.; Gomez-Mascaraque, L.; Arias, M.; Zampini, I.C.; Sayago, J.E.; Pino Ramos, L.L.; Schmeda-Hirschmann, G.; Lopez-Rubio, A.; Isla, M.I. Electrosprayed chitosan microcapsules as delivery vehicles for vaginal phytoformulations. Carbohydr. Polym. 2018, 201, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Isla, M.I.; Moreno, A.; Nuño, G.; Carabajal, A.; Alberto, M.R.; Zampini, C. Zuccagnia punctata Cav.: A review of its traditional uses, phytochemistry, pharmacology and toxicology. Nat. Prod. Commun. 2016, 11, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Chieli, E.; Romiti, N.; Zampini, I.C.; Garrido, G.; Isla, M.I. Effects of Zuccagnia punctata extracts and their flavonoids on the function and expression of ABCB1/P-glycoprotein multidrug transporter. J. Ethnopharmacol. 2012, 144, 797–801. [Google Scholar] [CrossRef]
- De la Rocha, N.; María, A.O.; Gianello, J.C.; Pelzer, L. Cytoprotective effects of chalcones from Zuccagnia punctata and melatonin on gastroduodenal tract in rats. Pharmacol. Res. 2003, 48, 97–99. [Google Scholar] [CrossRef]
- Morán-Vieyra, F.; Boggetti, H.J.; Zampini, I.C.; Ordoñez, R.M.; Isla, M.I.; Alvarez, R.M.S.; De Rosso, V.; Mercadante, A.Z.; Borsarelli, C.D. Singlet oxygen quenching and radical scavenging capacities of structurally related flavonoids present in Zuccagnia punctata Cav. Free Radic. Res. 2009, 43, 553–564. [Google Scholar] [CrossRef]
- Zampini, I.C.; Vattuone, M.A.; Isla, M.I. Antibacterial activity of Zuccagnia punctata Cav. ethanolic extracts. J. Ethnopharmacol. 2005, 102, 450–456. [Google Scholar] [CrossRef]
- Zampini, I.C.; Villarini, M.; Moretti, M.; Dominici, L.; Isla, M.I. Evaluation of genotoxic and antigenotoxic effects of hydroalcoholic extracts of Zuccagnia punctata Cav. J. Ethnopharmacol. 2008, 115, 330–335. [Google Scholar] [CrossRef]
- Zampini, I.C.; Villena, J.; Salva, S.; Herrera, M.; Isla, M.I.; Alvarez, S. Potentiality of standardized extract and isolated flavonoids from Zuccagnia punctata for the treatment of respiratory infections by Streptococcus pneumoniae: In vitro and in vivo studies. J. Ethnopharmacol. 2012, 140, 287–292. [Google Scholar] [CrossRef]
- Chahar, F.C.; Alvarez, P.E.; Zampini, C.; Isla, M.I.; Brandán, S.A. Experimental and DFT studies on 2′,4′-dihydroxychalcone, a product isolated from Zuccagnia punctata Cav. (Fabaceae) medicinal plant. J. Mol. Struct. 2020, 1201, 127221. [Google Scholar] [CrossRef]
- Wollenweber, E.; Seigler, D.S. Flavonoids from the exudate of Acacia neovernicosa. Phytochemistry 1982, 21, 1063–1066. [Google Scholar] [CrossRef]
- CLSI (Clinical and Laboratory Standards Institute). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; CLSI Standard M38; CLSI: Berwyn, PA, USA, 2017; pp. 1–35. [Google Scholar]
- Gupta, V.K.; Tuohy, M.G.; Gaur, R. Methods for high-quality DNA extraction from fungi. In Laboratory Protocols in Fungal Biology; Gupta, V., Tuohy, M., Ayyachamy, M., Turner, K., O’Donovan, A., Eds.; Springer: New York, NY, USA, 2012; pp. 403–406. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA Genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. Available online: https://msafungi.org/wp-content/uploads/2019/03/February-2013-Inoculum.pdf (accessed on 20 June 2022).
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Zaretskaya, I. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Rahalison, L.; Hamburger, M.; Hostettmann, K.; Monod, M.; Frenk, E. A bioautographic agar overlay method for the detection of antifungal compounds from higher plants. Phytochem. Anal. 1991, 2, 199–203. [Google Scholar] [CrossRef]
- Saxena, G.; Farmer, S.; Towers, G.H.N.; Hancock, R.E.W. Use of specific dyes in the detection of antimicrobial compounds from crude plant extracts using a thin layer chromatography agar overlay technique. Phytochem. Anal. 1995, 6, 125–129. [Google Scholar] [CrossRef]
- Egüen, B.; Melgarejo, P.; De Cal, A. Sensitivity of Monilinia fructicola from Spanish peach orchards to thiophanate-methyl, iprodione, and cyproconazole: Fitness analysis and competitiveness. Eur. J. Plant Pathol. 2015, 141, 789–801. [Google Scholar] [CrossRef]
- Eckert, J.W.; Brown, G.E. Evaluation of postharvest treatments for citrus fruits. In Methods for Evaluating Pesticides for Control of Plant Pathogens; Hickey, K.D., Ed.; American Phytopathological Society Press: St. Paul, MN, USA, 1986; pp. 92–97. [Google Scholar]
- Ferretti, A.C.; Tonucci, F.M.; Hidalgo, F.; Almada, E.; Larocca, M.C.; Favre, C. AMPK and PKA Interaction in the Regulation of Survival of Liver Cancer Cells Subjected to Glucose Starvation. Oncotarget 2016, 7, 17815–17828. [Google Scholar] [CrossRef]
- Martini, C.; Mari, M. Monilinia fructicola, Monilinia laxa (Monilinia rot, brown rot). In Postharvest Decay. Control Strategies; Bautista-Baños, S., Ed.; Academic Press: Waltham, MA, USA, 2014; pp. 233–265. [Google Scholar]
- Holetz, F.B.; Pessini, G.L.; Sanches, N.R.; Cortez, D.A.G.; Nakamura, C.V.; Dias Filho, B.P. Screening of some plants used in the Brazilian folk medicine for the treatment of infectious diseases. Memórias Inst. Oswaldo Cruz 2002, 97, 1027–1031. [Google Scholar] [CrossRef]
- Deresa, E.M.; Diriba, T.F. Phytochemicals as alternative fungicides for controlling plant diseases: A comprehensive review of their efficacy, commercial representatives, advantages, challenges for adoption, and possible solutions. Heliyon 2023, 9, e13810. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Philogène, B.J.R. Past and current prospects for the use of botanicals and plant allelochemicals in integrated pest management. Pharmaceut. Biol. 2008, 46, 41–52. [Google Scholar] [CrossRef]
- Jiménez-Reyes, M.F.; Carrasco, H.; Olea, A.F.; Silva-Moreno, E. Natural compounds: A sustainable alternative to the phytopathogens control. J. Chil. Chem. Soc. 2019, 64, 4459–4465. [Google Scholar] [CrossRef]
- Ammar, H.A.; El-Helaly, S.H.; Abdel-Rahman, T.A. Efficacy of some plant extracts and fungicides to control Alternaria black spots caused by Alternaria alternata on postharvest mango fruits in Egypt. Menoufia J. Plant Prot. 2022, 7, 29–40. [Google Scholar] [CrossRef]
- Mendy, T.K.; Misran, A.; Mahmud, T.M.M.; Ismail, S.I. Antifungal properties of Aloe vera through in vitro and in vivo screening against postharvest pathogens of papaya fruit. Sci. Hortic. 2019, 257, 108767. [Google Scholar] [CrossRef]
- Hasan, A.M.; Marzani, Q.A.; Abdulrahman, D.N. Management of fungal postharvest diseases of apple and pear using plant extracts. J. Duhok Univ. 2022, 25, 66–78. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. Application of plant extracts to control postharvest gray mold and susceptibility of apple fruits to B. cinerea from different plant hosts. Foods 2020, 9, 1430. [Google Scholar] [CrossRef]
- Di Liberto, M.G.; Seimandi, G.M.; Fernández, L.N.; Ruiz, V.E.; Svetaz, L.A.; Derita, M.G. Botanical control of citrus green mold and peach brown rot on fruits assays using a Persicaria acuminata phytochemically characterized extract. Plants 2021, 10, 425–439. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | MICs (µg/mL) | MFCs (µg/mL) |
|---|---|---|
| ZpuE | 250 | 250 |
| 1 | 62.5 | 125 |
| 2 | 125 | 125 |
| Cbz | 0.97 | 0.97 |
| Cap | 1.9 | 1.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Liberto, M.G.; Stegmayer, M.I.; Fernández, L.N.; Quiroga, A.D.; Svetaz, L.A.; Derita, M.G. Control of Brown Rot Produced by Monilinia fructicola in Peaches Using a Full-Spectrum Extract of Zuccagnia punctata Cav. Horticulturae 2023, 9, 1141. https://doi.org/10.3390/horticulturae9101141
Di Liberto MG, Stegmayer MI, Fernández LN, Quiroga AD, Svetaz LA, Derita MG. Control of Brown Rot Produced by Monilinia fructicola in Peaches Using a Full-Spectrum Extract of Zuccagnia punctata Cav. Horticulturae. 2023; 9(10):1141. https://doi.org/10.3390/horticulturae9101141
Chicago/Turabian StyleDi Liberto, Melina G., María Inés Stegmayer, Laura N. Fernández, Ariel D. Quiroga, Laura A. Svetaz, and Marcos G. Derita. 2023. "Control of Brown Rot Produced by Monilinia fructicola in Peaches Using a Full-Spectrum Extract of Zuccagnia punctata Cav." Horticulturae 9, no. 10: 1141. https://doi.org/10.3390/horticulturae9101141
APA StyleDi Liberto, M. G., Stegmayer, M. I., Fernández, L. N., Quiroga, A. D., Svetaz, L. A., & Derita, M. G. (2023). Control of Brown Rot Produced by Monilinia fructicola in Peaches Using a Full-Spectrum Extract of Zuccagnia punctata Cav. Horticulturae, 9(10), 1141. https://doi.org/10.3390/horticulturae9101141