
Effect of Seaweed-Based Biostimulants on Growth and Development of Hydrangea paniculata under Continuous or Periodic Drought Stress
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Experiment 1—Deficit Irrigation (2019)
2.3. Experiment 2—Repeated Drying and Wetting Cycles (2021)
2.4. Substrate Moisture Content Measurements
2.5. Plant Physiological Responses
2.6. Morphological Parameters
2.7. Statistical Analysis
3. Results
3.1. Effect of Biostimulants under Deficit Irrigation in 2019
3.1.1. Effect of Deficit Irrigation
3.1.2. Effect of Biostimulants
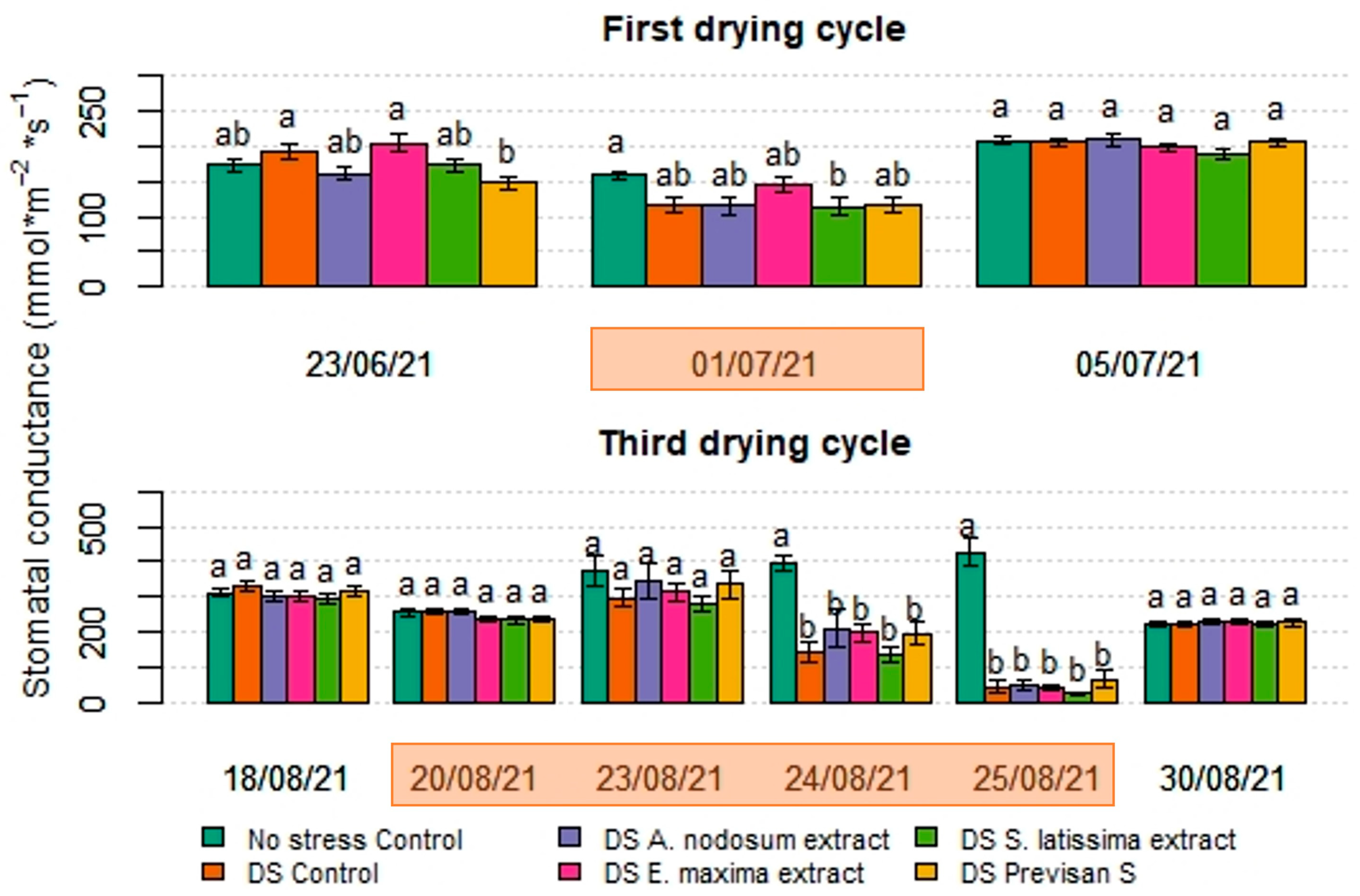
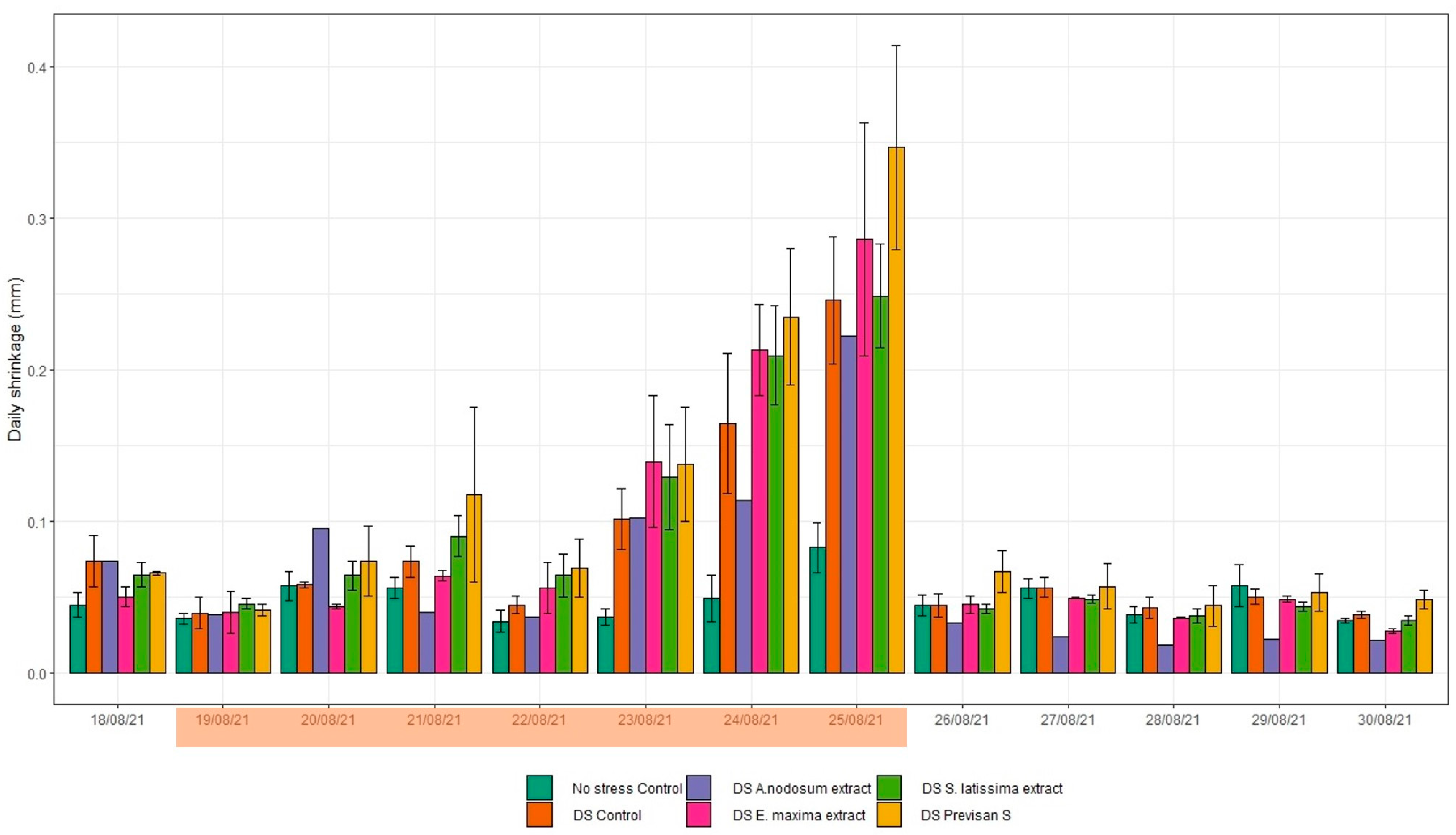
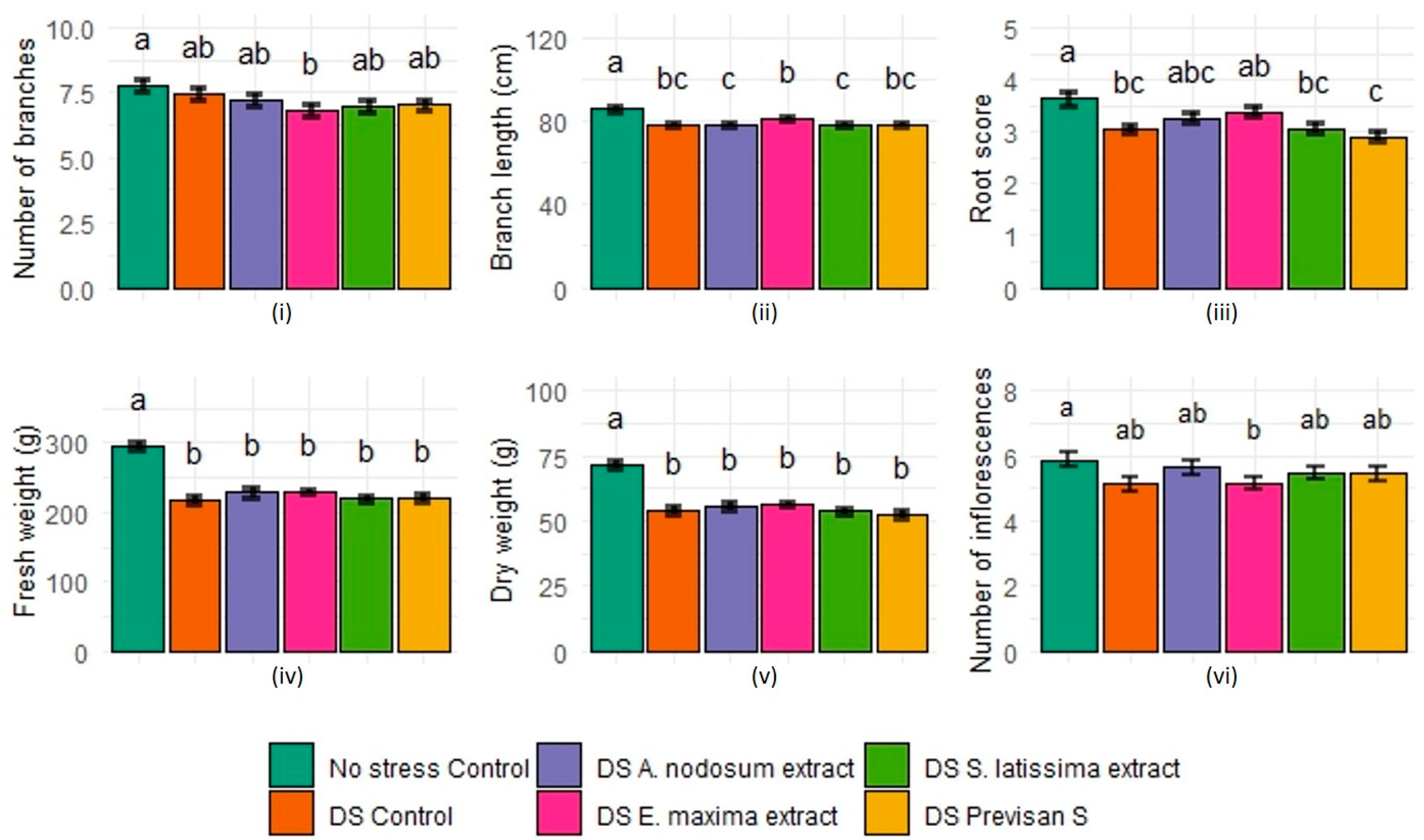
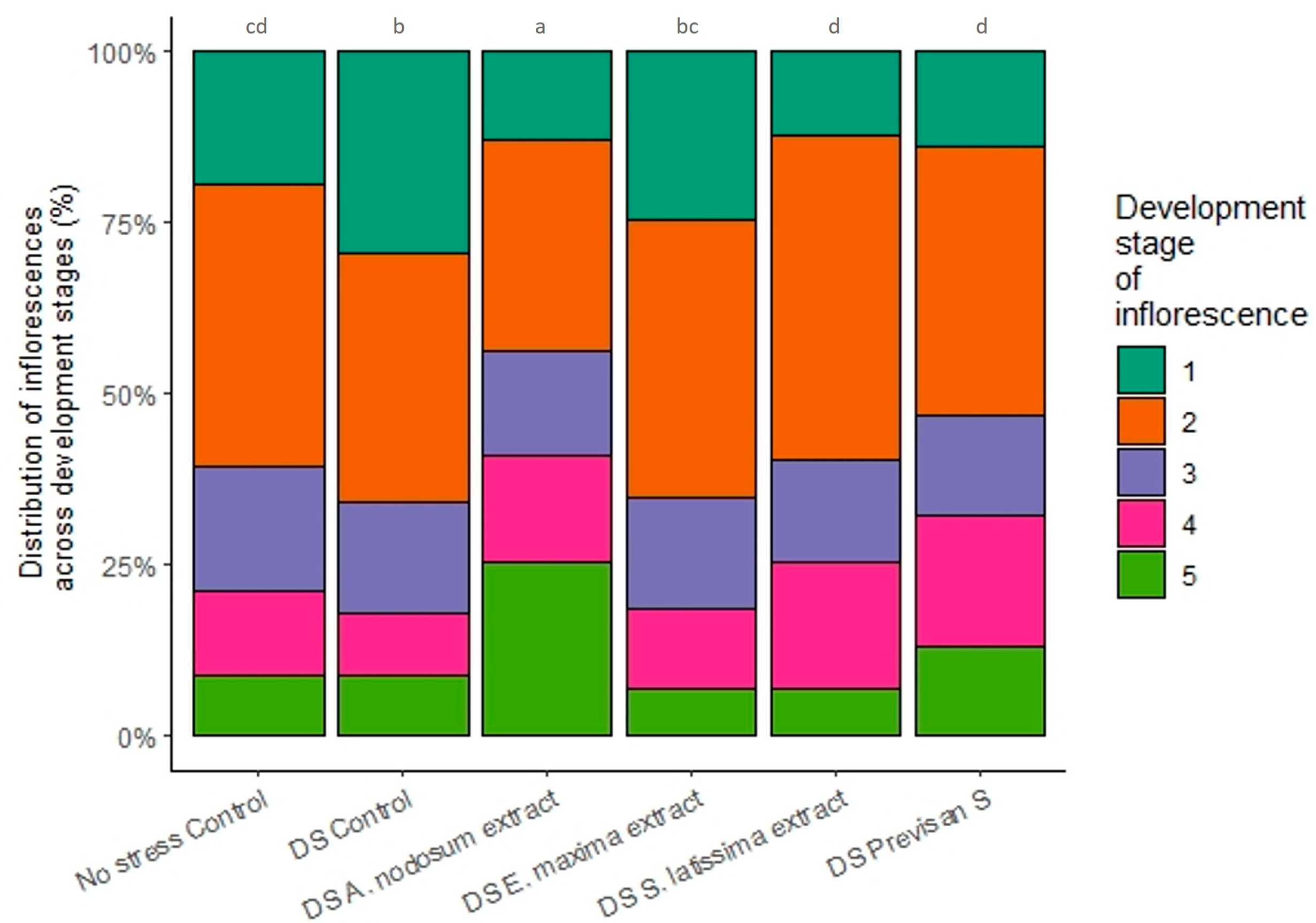
3.2. Effect of Biostimulants under Repeated Drying and Wetting Cycles in 2021
4. Discussion
4.1. Effect of Deficit Irrigation and Repeated Drying Cycles on Hydrangea
4.2. Effects of Biostimulants on Hydrangea Grown under Deficit Irrigation or Repeated Drying Cycles
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Departement Landbouw en Visserij. Landbouwrapport 2018: Uitdagingen Voor de Vlaamse Land- En Tuinbouw; Departement Landbouw en Visserij: Brussel, Belgium, 2018. [Google Scholar]
- Departement Landbouw en Visserij. Landbouwrapport 2020; Departement Landbouw en Visserij: Brussel, Belgium, 2020. [Google Scholar]
- Cook, B.I.; Mankin, J.S.; Anchukaitis, K.J. Climate Change and Drought: From Past to Future. Curr. Clim. Chang. Rep. 2018, 4, 164–179. [Google Scholar] [CrossRef]
- European Union Regulation (EC). No 765/2008 of the European Parliament and of the Council (4) Lays down Rules on the Accredi; European Union Regulation (EC): Brussel, Belgium, 2019. [Google Scholar]
- du Jardin, P. The Science of Plant Biostimulants—A Bibliographic Analysis; Ad hoc Study Report to the European Commission; European Union: Brussel, Belgium, 2012. [Google Scholar]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed Extract Stimuli in Plant Science and Agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed Extracts as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Whapham, C.A.; Blunden, G.; Jenkins, T.; Hankins, S.D. Significance of Betaines in the Increased Chlorophyll Content of Plants Treated with Seaweed Extract. J. Appl. Phycol. 1993, 5, 231. [Google Scholar] [CrossRef]
- Wally, O.S.D.; Critchley, A.T.; Hiltz, D.; Craigie, J.S.; Han, X.; Zaharia, L.I.; Abrams, S.R.; Prithiviraj, B. Regulation of Phytohormone Biosynthesis and Accumulation in Arabidopsis Following Treatment with Commercial Extract from the Marine Macroalga Ascophyllum Nodosum. J. Plant Growth Regul. 2013, 32, 324–339. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Mittler R Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [CrossRef]
- Blunden, G.; Jenkins, T.; Liu, Y.-W. Enhanced Leaf Chlorophyll Levels in Plants Treated with Seaweed Extract. J. Appl. Phycol. 1996, 8, 535–543. [Google Scholar] [CrossRef]
- Zhang, X.; Ervin, E.H. Cytokinin-Containing Seaweed and Humic Acid Extracts Associated with Creeping Bentgrass Leaf Cytokinins and Drought Resistance. Crop Sci. 2004, 44, 1737–1745. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W.; Harlan, J.C. Monitoring the Vernal Advancement and Retrogradation (Greenwave Effect) of Natural Vegetation; Remote Sensing Center, Texas A&M University: College Station, TX, USA, 1974. [Google Scholar]
- Lichtenthaler, H.K. Vegetation Stress: An Introduction to the Stress Concept in Plants. J. Plant Physiol. 1996, 148, 4–14. [Google Scholar] [CrossRef]
- Carter, G.A. Ratios of Leaf Reflectances in Narrow Wavebands as Indicators of Plant Stress. Int. J. Remote Sens. 1994, 15, 697–703. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical Properties and Nondestructive Estimation of Anthocyanin Content in Plant Leaves. Photochem. Photobiol. 2001, 74, 38–45. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence-a Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Auguie, B.; Antonov, A. GridExtra: Miscellaneous Functions for “Grid” Graphics. R Package Version 2.3. 2017. Available online: https://cran.r-project.org/web/packages/gridExtra/index.html (accessed on 23 January 2023).
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes. R Package Sersion 1.1-3. 2022. Available online: https://cran.r-project.org/web/packages/RColorBrewer/RColorBrewer.pdf (accessed on 23 January 2023).
- Wickham, H.; François, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. R-Package Version 1.1.1. 2022. Available online: https://cran.r-project.org/web/packages/dplyr/dplyr.pdf (accessed on 23 January 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation. R-Package Version 2.4.26. 2022. Available online: https://cran.r-project.org/web/packages/rcompanion/index.html (accessed on 23 January 2023).
- Ogle, D.H.; Doll, J.C.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis. R Package Version 0.9.4. 2022. Available online: https://CRAN.R-project.org/package=FSA (accessed on 23 January 2023).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression. R Package Version 3.1-2. 2019. Available online: https://cran.r-project.org/web/packages/car/index.html (accessed on 23 January 2023).
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-5. 2021. Available online: https://cran.r-project.org/web/packages/agricolae/agricolae.pdf (accessed on 23 January 2023).
- Wickham, H.; Bryan, J. Readxl: Read Excel Files. R Package. R Package Version 1.4.2. 2022. Available online: https://cran.r-project.org/web/packages/readxl/index.html (accessed on 23 January 2023).
- Wickham, H. Forcats: Tools for Working with Categorical Variables (Factors). R Package Version 1.0.0. 2021. Available online: https://cran.r-project.org/web/packages/forcats/index.html (accessed on 23 January 2023).
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W. H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Gitelson, A.A.; Merzlyak, M.N.; Lichtenthaler, H.K. Detection of Red Edge Position and Chlorophyll Content by Reflectance Measurements near 700 Nm. J. Plant Physiol. 1996, 148, 501–508. [Google Scholar] [CrossRef]
- Marenco, R.A.; Antezana-Vera, S.A.; Nascimento, H.C.S. Relationship between Specific Leaf Area, Leaf Thickness, Leaf Water Content and SPAD-502 Readings in Six Amazonian Tree Species. Photosynthetica 2009, 47, 184–190. [Google Scholar] [CrossRef]
- Martínez, D.E.; Guiamet, J.J. Distortion of the SPAD 502 Chlorophyll Meter Readings by Changes in Irradiance and Leaf Water Status. Agronomie 2004, 24, 41–46. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, R.; Mori, T.; Saito, K. Alternation of Flavonoid Accumulation under Drought Stress in Arabidopsis thaliana. Plant Signal. Behav. 2014, 9, e29518. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of Oxidative and Drought Tolerance in Arabidopsis by Overaccumulation of Antioxidant Flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R. Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 9783642326530. [Google Scholar]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and Metabolic Limitations to Photosynthesis under Drought and Salinity in C3 Plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, Y.G.; Guo, K.; Zheng, Y.R.; Li, G.Q.; Yu, L.F.; Yang, R. Influence of Drought Intensity on the Response of Six Woody Karst Species Subjected to Successive Cycles of Drought and Rewatering. Physiol. Plant. 2010, 139, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.C. Measurement and Influence of Environmental and Plant Factors on Stomatal Conductance in the Field. Agric. For. Meteorol. 1991, 54, 137–154. [Google Scholar] [CrossRef]
- Basiri Jahromi, N.; Fulcher, A.; Walker, F.; Altland, J. Photosynthesis, Growth, and Water Use of Hydrangea Paniculata ‘Silver Dollar’ Using a Physiological-Based or a Substrate Physical Properties-Based Irrigation Schedule and a Biochar Substrate Amendment. Irrig. Sci. 2020, 38, 263–274. [Google Scholar] [CrossRef]
- Kim, J. Water Relations of Bedding Plants: Water Use, Drought Physiology, and Gene Expression. Ph.D. Thesis, University of Georgia, Athens, Greece, 2011. [Google Scholar]
- Flexas, J.; Medrano, H. Drought-Inhibition of Photosynthesis in C3 Plants: Stomatal and Non-Stomatal Limitations Revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Barradas, V.L.; Jones, H.G. Responses of CO2 Assimilation to Changes in Irradiance: Laboratory and Field Data and a Model for Beans (Phaseolus vulgaris L.). J. Exp. Bot. 1996, 47, 639–645. [Google Scholar] [CrossRef]
- Turner, N.C.; Schulze, E.-D.; Gollan, T. The Responses of Stomata and Leaf Gas Exchange to Vapour Pressure Deficits and Soil Water Content. Oecologia 1984, 63, 338–342. [Google Scholar] [CrossRef]
- Steppe, K.; Sterck, F.; Deslauriers, A. Diel Growth Dynamics in Tree Stems: Linking Anatomy and Ecophysiology. Trends Plant Sci. 2015, 20, 335–343. [Google Scholar] [CrossRef]
- de Swaef, T.; de Schepper, V.; Vandegehuchte, M.W.; Steppe, K. Stem Diameter Variations as a Versatile Research Tool in Ecophysiology. Tree Physiol. 2015, 35, 1047–1061. [Google Scholar] [CrossRef] [PubMed]
- Cameron, R.W.F.; Harrison-Murray, R.S.; Atkinson, C.J.; Judd, H.L. Regulated Deficit Irrigation—A Means to Control Growth in Woody Ornamentals. J. Hortic. Sci. Biotechnol. 2006, 81, 435–443. [Google Scholar] [CrossRef]
- Cameron, R.; Harrison-Murray, R.; Fordham, M.; Wilkinson, S.; Davies, W.; Atkinson, C.; Else, M. Regulated Irrigation of Woody Ornamentals to Improve Plant Quality and Precondition against Drought Stress. Ann. Appl. Biol. 2008, 153, 49–61. [Google Scholar] [CrossRef]
- Wada, K.C.; Takeno, K. Stress-Induced Flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef]
- Irani, H.; ValizadehKaji, B.; Naeini, M.R. Biostimulant-Induced Drought Tolerance in Grapevine Is Associated with Physiological and Biochemical Changes. Chem. Biol. Technol. Agric. 2021, 8, 1–13. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A Commercial Extract of Brown Macroalga (Ascophyllum Nodosum) Affects Yield and the Nutritional Quality of Spinach In Vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Effect on Yield, Total Phenolic, Total Flavonoid and Total Isothiocyanate Content of Two Broccoli Cultivars (Brassica oleraceae Var Italica) Following the Application of a Commercal Brown Seaweed Extract. Agric. Food Sci. 2014, 23, 28–37. [Google Scholar] [CrossRef]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Enhancement of Phenolic and Flavonoid Compounds in Cabbage (Brassica Oleraceae) Following Application of Commercial Seaweed Extracts of the Brown Seaweed (Ascophyllum Nodosum). Agric. Food Sci. 2013, 22, 288–295. [Google Scholar] [CrossRef]
- Elansary, H.O.; Norrie, J.; Ali, H.M.; Salem, M.Z.M.; Mahmoud, E.A.; Yessoufou, K. Enhancement of Calibrachoa Growth, Secondary Metabolites and Bioactivity Using Seaweed Extracts. BMC Complement. Altern. Med. 2016, 16, 341. [Google Scholar] [CrossRef]
- Xu, C.; Leskovar, D.I. Effects of A. Nodosum Seaweed Extracts on Spinach Growth, Physiology and Nutrition Value under Drought Stress. Sci. Hortic. 2015, 183, 39–47. [Google Scholar] [CrossRef]
- Jones, R.J.; Mansfield, T.A. Suppression of Stomatal Opening in Leaves Treated with Abscisic Acid. J. Exp. Bot. 1970, 21, 714–719. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Agliassa, C.; Mannino, G.; Vigliante, I.; Contartese, V.; Secchi, F.; Bertea, C.M. A Biostimulant Based on Seaweed (Ascophyllum Nodosum and Laminaria Digitata) and Yeast Extracts Mitigateswater Stress Effects on Tomato (Solanum Lycopersicum l.). Agriculture 2021, 11, 557. [Google Scholar] [CrossRef]
- Kałuzewicz, A.; Krzesiński, W.; Spizewski, T.; Zaworska, A. Effect of Biostimulants on Several Physiological Characteristics and Chlorophyll Content in Broccoli under Drought Stress and Re-Watering. Not. Bot. Horti Agrobot. Cluj Napoca 2017, 45, 197–202. [Google Scholar] [CrossRef]
- Top, S.; Vandoorne, B.; Pauwels, E.; Perneel, M.; Van Labeke, M.-C.; Steppe, K. Plant Sensors Untangle the Water-Use and Growth Effects of Selected Seaweed-Derived Biostimulants on Drought-Stressed Tomato Plants (Solanum lycopersicum). J. Plant Growth Regul. 2023. [Google Scholar] [CrossRef]
- Sumangala, K.; Srikrishnah, S.; Sutharsan, S. Roses Growth and Flowering Responding to Concentration and Frequency of Seaweed (Sargassum crassifolium L.) Liquid Extract Application. Curr. Agric. Res. J. 2019, 7, 236–244. [Google Scholar] [CrossRef]
- Krajnc, A.U.; Ivanuš, A.; Kristl, J.; Šušek, A. Seaweed Extract Elicits the Metabolic Responses in Leaves and Enhances Growth of Pelargonium Cuttings. Eur. J. Hortic. Sci. 2012, 77, 170–181. [Google Scholar]
- Pohl, A.; Grabowska, A.; Kalisz, A.; Ekara, A.S. Biostimulant Application Enhances Fruit Setting in Eggplant—An Insight into the Biology of Flowering. Agronomy 2019, 9, 482. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial Year | Biostimulant | Application Dose | Application Frequency |
|---|---|---|---|
| 2019 | Ascophyllum nodosum extract (Phylgreen, Tradecorp) | 1.5 L ha−1 | Every 15 days |
| Soliera chordalis extract (SeaMelPure, Olmix) | 2 L ha−1 | One application two weeks after planting | |
| Ecklonia maxima extract (Kelpak, Kelp Products International) | 2.5 L ha−1 | First application: 7–10 days after planting; repeated at 14–21 days intervals up to four applications | |
| 2021 | Ascophyllum nodosum extract combined with product based on plant-based amino acids (Phylgreen + Delfan Plus V, Tradecorp) | Phylgreen: 0.5 mL L−1 Delfan Plus V: 2 mL L−1 | Phylgreen: Every 15 days Delfan Plus V: during stress |
| Ecklonia maxima extract (Kelpak, Kelp Products International) | 2.5 L ha−1 | First application: 7–10 days after planting; repeated at 14–21 days intervals up to four applications | |
| Saccharina latissima extract (experimental product) | 3 mL L−1 | Every two weeks | |
| Previsan S (Agriton) | 30 mL L−1 | Every two weeks |
| Reflectance Index | Formula | Reference |
|---|---|---|
| NDVI (Normalized Difference Vegetation Index) | Ref. [17] | |
| Lic1 (Lichtenthaler Index 1) | Ref. [18] | |
| Ctr2 (Carter Index 2) | Ref. [19] | |
| ARI1 (Anthocyanin Reflectance Index 1) | Ref. [20] |
| Parameter | Standard Irrigation (100%) | Deficit Irrigation (80%) (DS Control) | Statistics | ||||
|---|---|---|---|---|---|---|---|
| Substrate-related parameters | |||||||
| Volumetric moisture content [vol%] | 26.6 | ± | 1.1 a | 20.9 | ± | 1.2 b | p = 0.0003 1 |
| EC 3 [mS.m−1] | 221.3 | ± | 7.1 a | 178.6 | ± | 6.0 b | p = 0.0005 1 |
| Plant-related parameters | |||||||
| Chlorophyll index [-] | 22.25 | ± | 0.50 b | 25.00 | ± | 0.49 a | p < 0.0001 1 |
| Flavonol index [-] | 0.675 | ± | 0.019 a | 0.712 | ± | 0.026 a | p = 0.2394 1 |
| REIP 4 [nm] | 702.8 | ± | 0.9 b | 704.4 | ± | 0.8 a | p = 0.0464 2 |
| NDVI 5 [-] | 0.505 | ± | 0.005 a | 0.506 | ± | 0.004 a | p = 0.8599 1 |
| Ctr2 6 [-] | 0.438 | ± | 0.006 a | 0.432 | ± | 0.004 a | p = 0.9867 2 |
| Lic1 7 [-] | 0.573 | ± | 0.005 a | 0.576 | ± | 0.004 a | p = 0.7523 2 |
| ARI1 8 [-] | 0.296 | ± | 0.012 a | 0.251 | ± | 0.014 b | p = 0.0136 1 |
| FV/FM [-] | 0.794 | ± | 0.003 a | 0.784 | ± | 0.008 a | p= 0.0685 1 |
| Stomatal conductance [mmol m−2 s−1] | 239.8 | ± | 20.2 a | 132.1 | ± | 11.8 b | p = 0.0011 2 |
| Branch length [cm] | 63.1 | ± | 1.5 a | 31.5 | ± | 0.9 b | p < 0.0001 2 |
| Number of branches [-] | 9.8 | ± | 0.6 b | 11.6 | ± | 0.5 a | p = 0.0167 1 |
| Fresh weight [g] | 89.6 | ± | 4.3 a | 51.2 | ± | 2.5 b | p < 0.0001 1 |
| Dry weight [g] | 29.2 | ± | 1.3 a | 14.9 | ± | 0.7 b | p < 0.0001 2 |
| Water content [%] [(FW-DW)/FW × 100] | 67.3 | ± | 0.3 b | 70.7 | ± | 0.3 a | p < 0.0001 2 |
| Root development score [-] | 3.42 | ± | 0.10 a | 2.63 | ± | 0.2 b | p = 0.0002 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Clercq, P.; Pauwels, E.; Top, S.; Steppe, K.; Van Labeke, M.-C. Effect of Seaweed-Based Biostimulants on Growth and Development of Hydrangea paniculata under Continuous or Periodic Drought Stress. Horticulturae 2023, 9, 509. https://doi.org/10.3390/horticulturae9040509
De Clercq P, Pauwels E, Top S, Steppe K, Van Labeke M-C. Effect of Seaweed-Based Biostimulants on Growth and Development of Hydrangea paniculata under Continuous or Periodic Drought Stress. Horticulturae. 2023; 9(4):509. https://doi.org/10.3390/horticulturae9040509
Chicago/Turabian StyleDe Clercq, Paulien, Els Pauwels, Seppe Top, Kathy Steppe, and Marie-Christine Van Labeke. 2023. "Effect of Seaweed-Based Biostimulants on Growth and Development of Hydrangea paniculata under Continuous or Periodic Drought Stress" Horticulturae 9, no. 4: 509. https://doi.org/10.3390/horticulturae9040509
APA StyleDe Clercq, P., Pauwels, E., Top, S., Steppe, K., & Van Labeke, M.-C. (2023). Effect of Seaweed-Based Biostimulants on Growth and Development of Hydrangea paniculata under Continuous or Periodic Drought Stress. Horticulturae, 9(4), 509. https://doi.org/10.3390/horticulturae9040509