
Screening and Identification of Potato StSPS1, a Potential Crucial Gene Regulating Seed Potato Vigor

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions and Sampling Procedure
2.2. Identification and Bioinformatics Analysis
2.3. Gene Expression Analysis and Enzyme Activity Determination
2.4. Correlation Analysis
2.5. Statistical Analysis and Mapping
3. Results
3.1. Identification and Characteristics of the StSPS Family
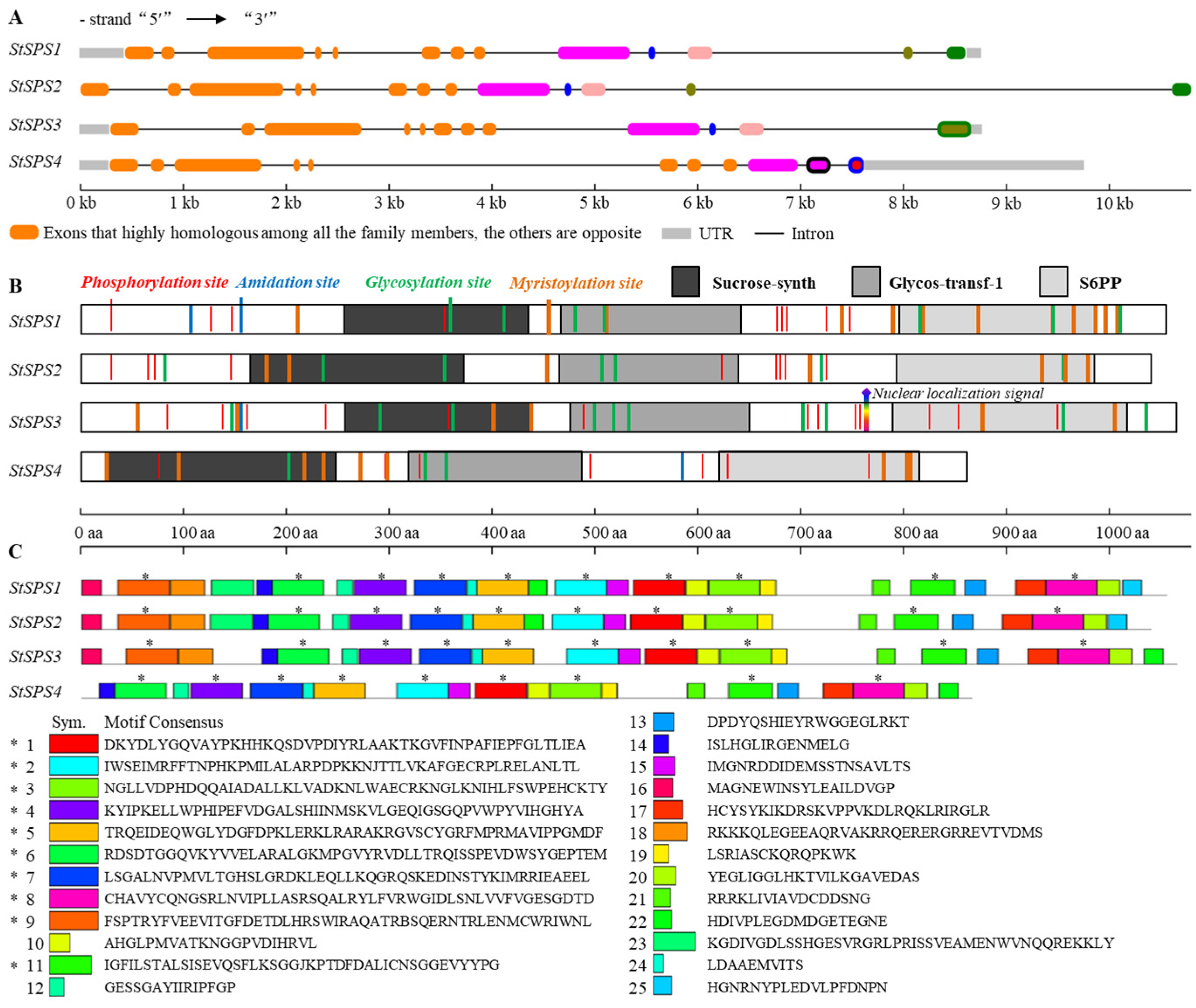
3.2. Gene and Protein Structure Analysis
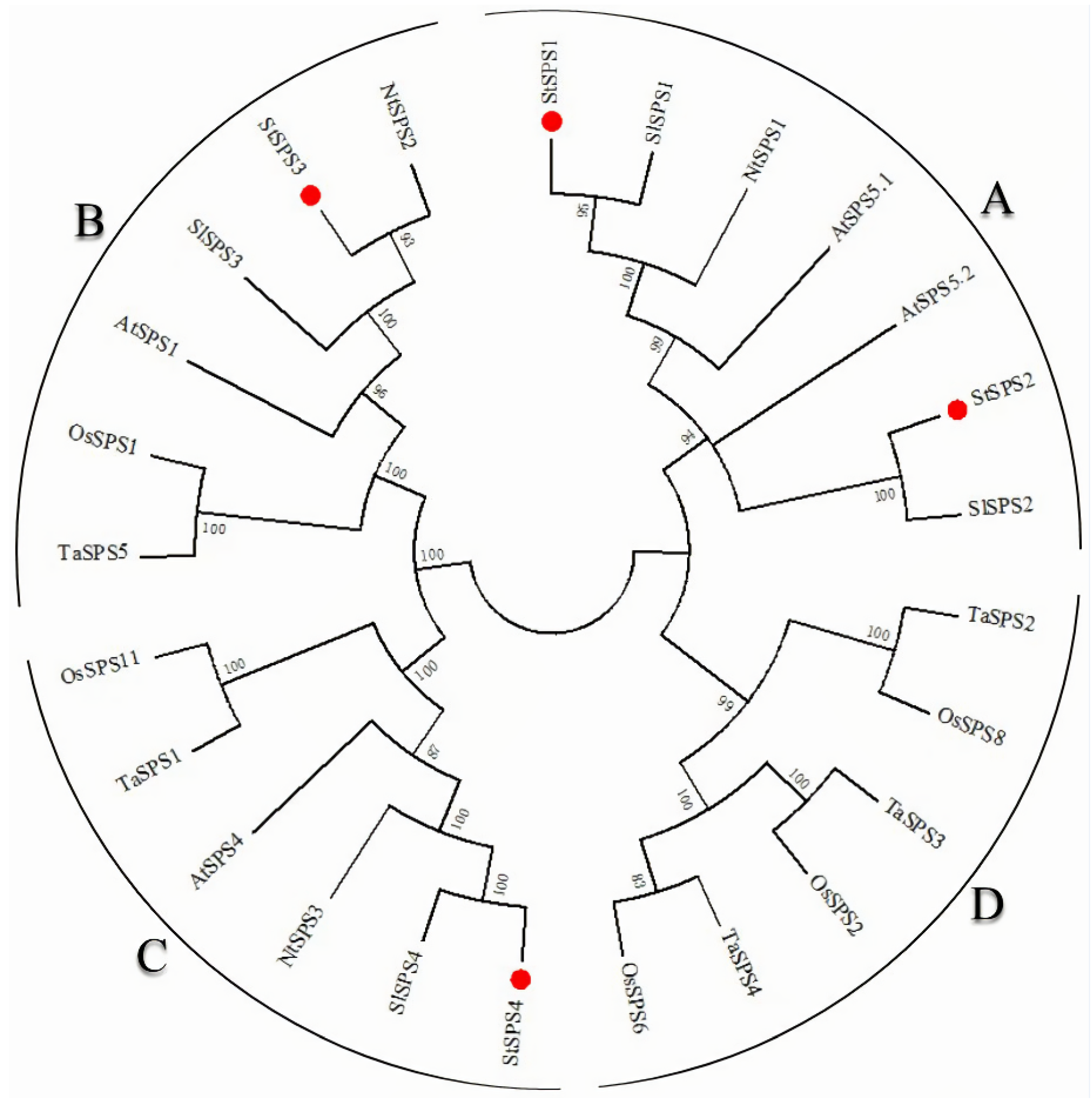
3.3. Phylogenetic Analysis of StSPS Family Members
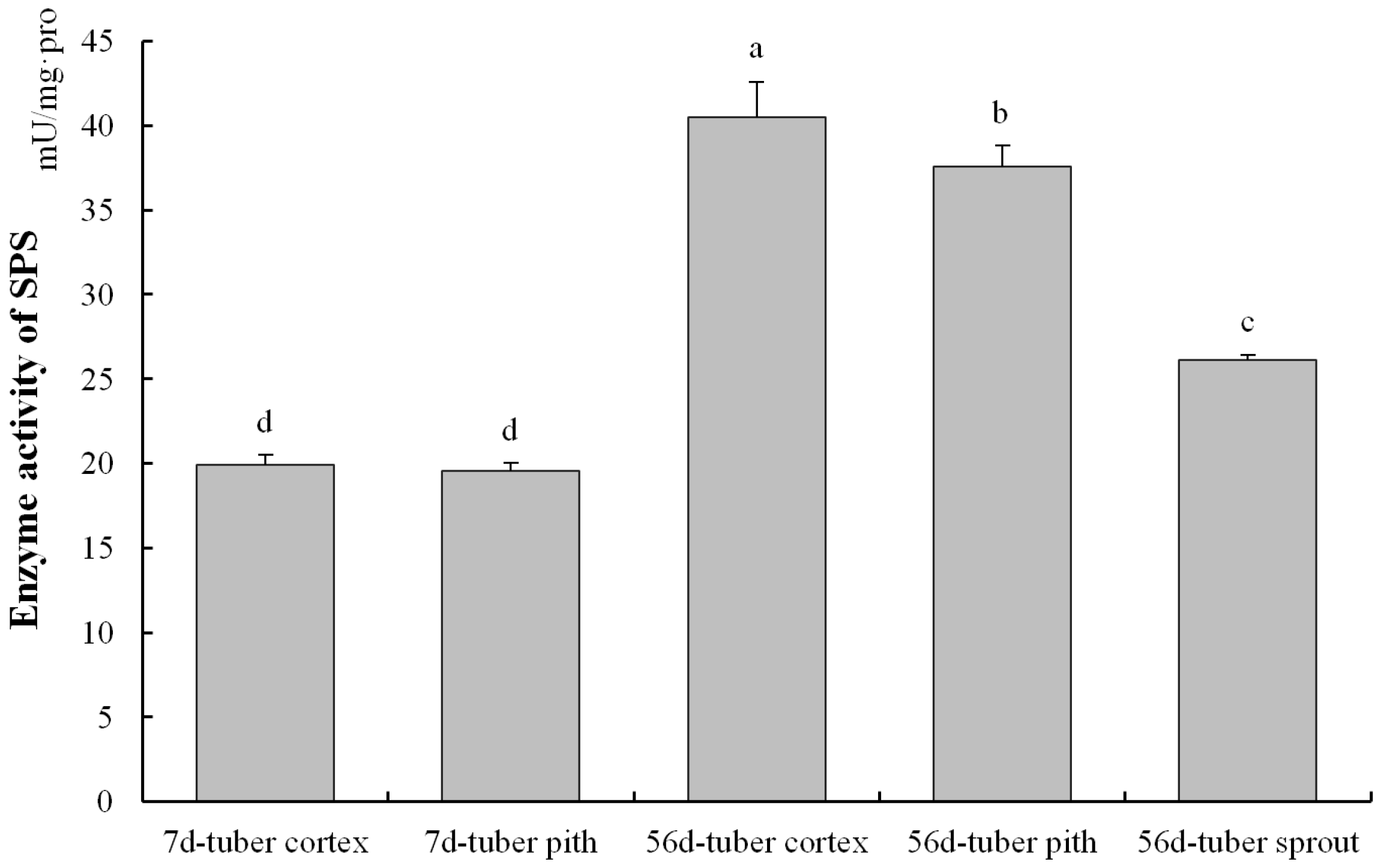
3.4. SPS Enzyme Analysis in Different Tuber Tissues
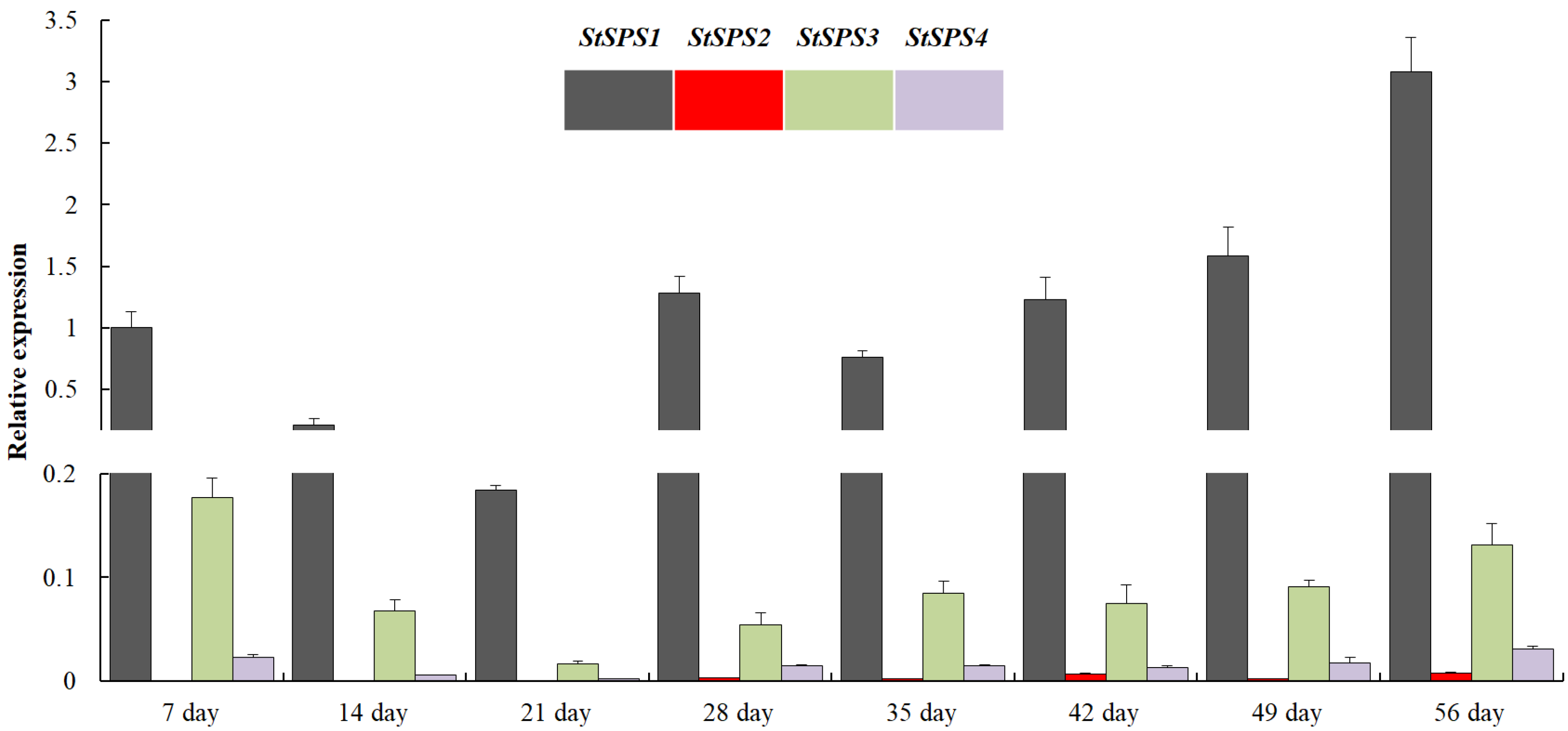
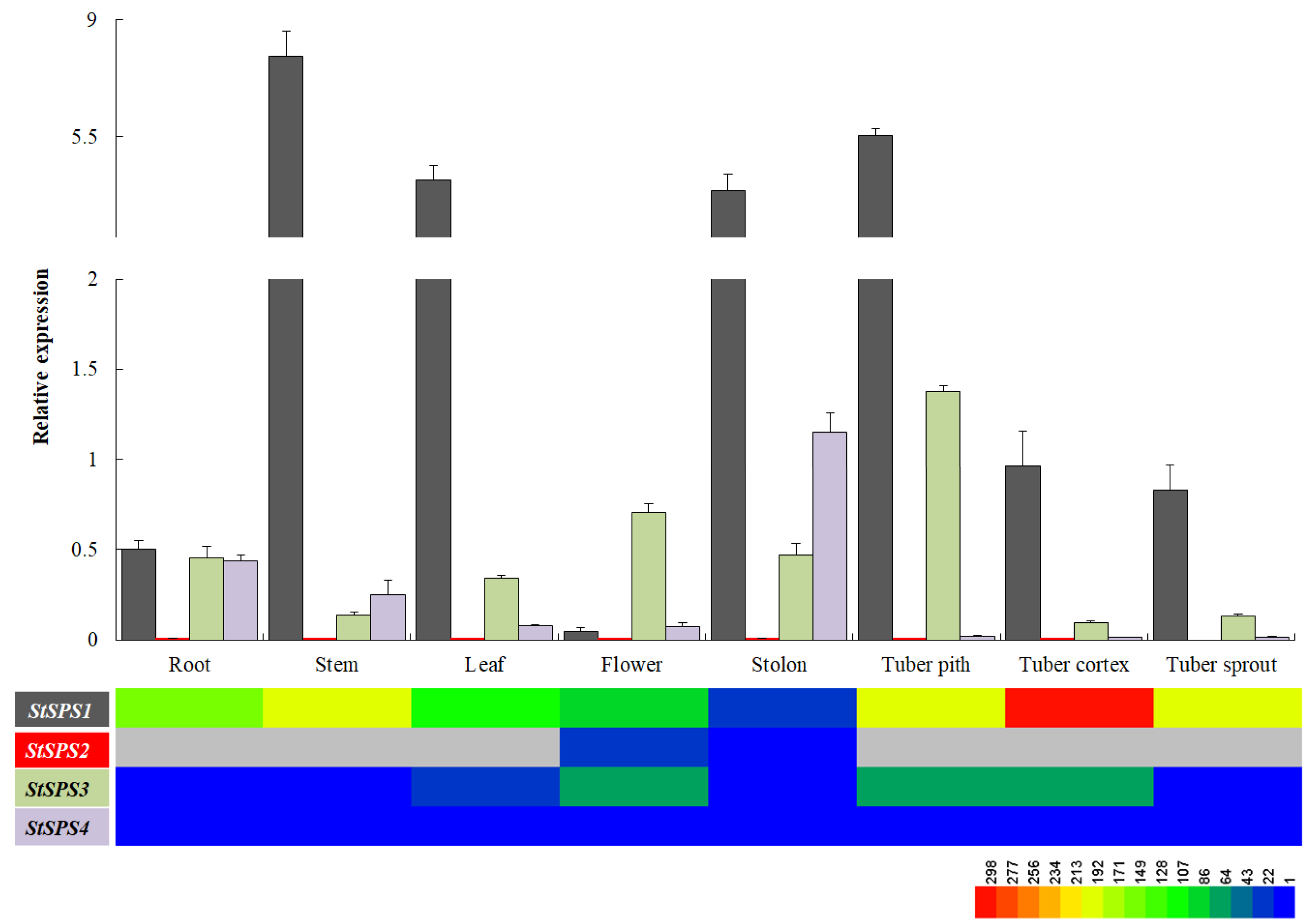
3.5. Expression Analysis of StSPS Family
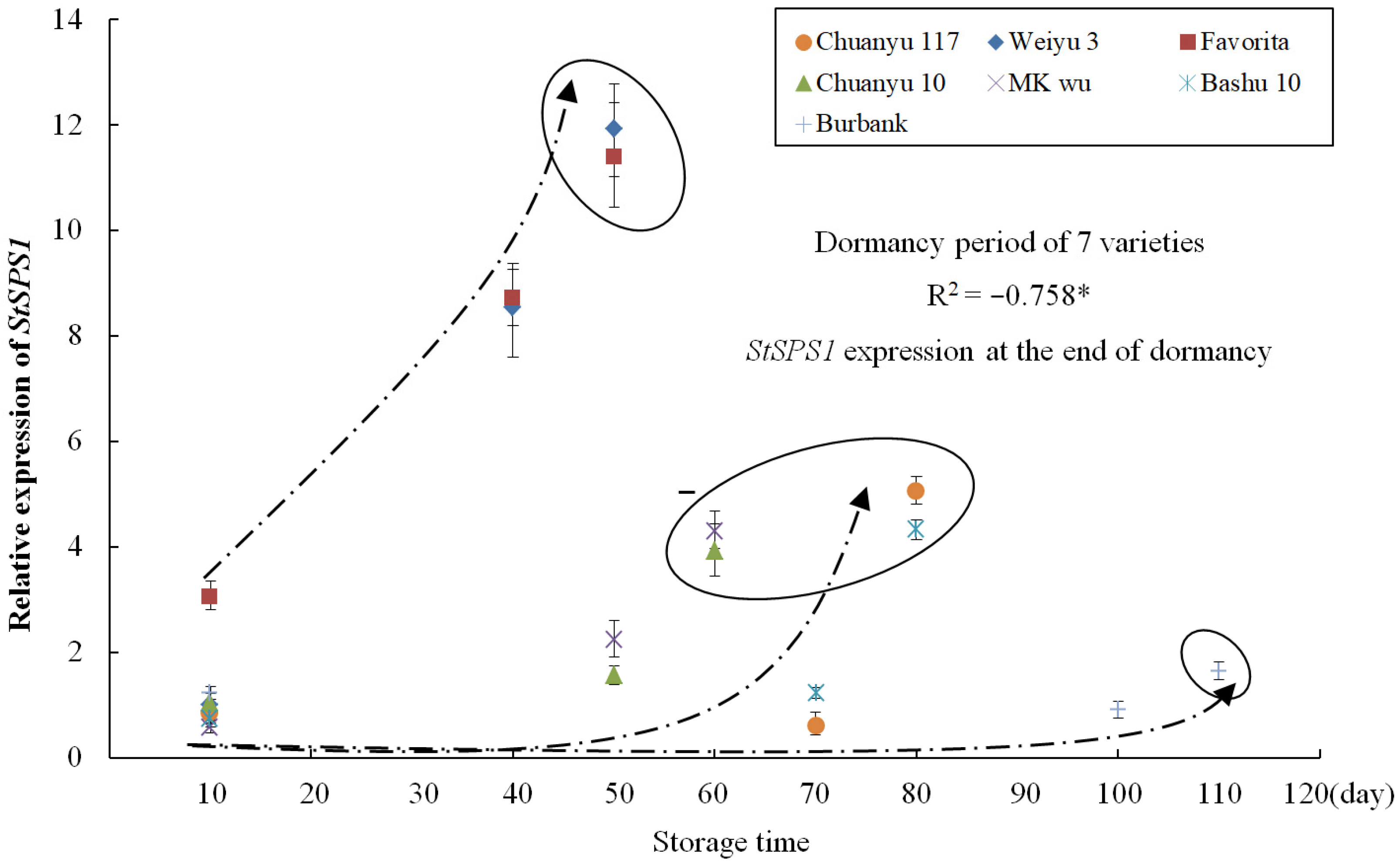
3.6. Correlation Analysis between StSPS1 and Seed Potato Vigor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, N.; Zhang, H.L.; Zhang, R.H.; Xu, Y.K. Current situation and prospect of potato planting in China. Chin. Potato J. 2021, 35, 81–96. [Google Scholar]
- Jansky, S.H.; Jin, L.P.; Xie, K.Y.; Xie, C.H.; Spooner, D.M. Potato production and breeding in China. Potato Res. 2009, 52, 57–65. [Google Scholar] [CrossRef]
- Pu, Y.L.; Wang, D.; Wang, R.B. Vigor and physiological dharacters of potato minitubers under different storage conditions. Acta Bot. Boreali-Occident. Sin. 2008, 28, 336–341. [Google Scholar]
- Caldiz, D.O. Physiological age research during the second half of the twentieth century. Potato Res. 2009, 52, 295–304. [Google Scholar] [CrossRef]
- Zou, X.; Ding, F.; Yu, J.L.; Peng, J.; Deng, M.S.; Wang, Y.; Liu, L.F.; Yu-Han, K.Z.; Chen, N.W.; Wang, X.Y. Suppression mechanism of volatile sprout-inhibitors on potato tuber sprouting. Acta Agron. Sin. 2019, 45, 235–247. [Google Scholar] [CrossRef]
- Si, H.; Zhang, N.; Wang, D. Molecular mechanism and regulation strategy of potato tuber dormancy and germination. Chin. Potato J. 2007, 21, 104–107. [Google Scholar]
- Zhong, L.; Deng, J.C.; Wang, L.J.; Zheng, S.L.; Dou, P.; Wang, X.L.; Yuan, J.C. Effect of plant growth regulators on germination and oxidase activity during storage of potato tubers. Acta Pratacult. Sin. 2021, 26, 147–157. [Google Scholar]
- Viola, R.; Pelloux, J.; van der Ploeg, A.; Gillespit, T.; Marquis, N.; Roberts, A.G.; Hancock, R.D. Symplastic connection is required for bud outgrowth following dormancy in potato (Solanum tuberosum L.) tubers. Plant Cell Environ. 2007, 30, 973–983. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Wang, X.F. Advances on relation-ship between biomembrane and seed vigor. Chin. Bull. Bot. 2004, 6, 641–648. [Google Scholar]
- Leloir, L.F.; Cardini, C.E. The biosynthesis of sucrose phosphate. J. Biol. Chem. 1955, 214, 157. [Google Scholar] [CrossRef]
- Huber, S.C.; Huber, J.L. Role and regulation of sucrose-phosphate synthase in higher plants. Annu. Rev. Plant Biol. 1996, 47, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.W.; Zhong, L.Y.; Feng, Y.; Zhu, W.M.; Zhang, Y.Y. Research advance of sucrose phosphate synthase (SPS) in higher plant. Plant Physiol. J. 2020, 56, 931–938. [Google Scholar]
- Lunn, J.E.; Macrae, E. New complexities in the synthesis of sucrose. Curr. Opin. Plant Biol. 2003, 6, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Castleden, C.K.; Aoki, N.; Gillespie, V.; Macrae, E.A.; Quick, W.P.; Bunchner, P.; Foyer, C.H.; Lunn, F.J.E. Evolution and function of the sucrose-phosphate synthase gene families in wheat and other grasses. Plant Physiol. 2004, 135, 1753–1764. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Chen, M.; Yao, Y.L.; Zhang, X.M. Research progress of plant sucrose phosphate synthase (SPS) gene. Mol. Plant Breed. 2022, 1, 12. [Google Scholar]
- Widhi, D.S.; Hirotaka, N.; Etsuko, I.I.; Bambang, S.; Toshiharu, H.; Atsushi, N. Purification and characterization of recombinant sugarcane sucrose phosphate synthase expressed in E. coli and insect Sf9 cells: An importance of the N-terminal domain for an allosteric regulatory property. J. Biochem. 2016, 159, 599–607. [Google Scholar]
- Wang, D.; Zhao, J.; Hu, B.; Li, J.; Qin, Y.; Chen, L.; Qin, Y.; Hu, G. Identification and expression profile analysis of the sucrose phosphate synthase gene family in Litchi chinensis Sonn. PeerJ 2018, 6, e4379. [Google Scholar] [CrossRef]
- Chen, L.Q.; Qu, X.Q.; Hou, B.H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207. [Google Scholar] [CrossRef]
- Hashida, Y.; Hirose, T.; Okamura, M.; Hibara, K.I.; Ohsugi, R.; Aoki, N. A reduction of sucrose phosphate synthase (SPS) activity affects sucrose/starch ratio in leaves but does not inhibit normal plant growth in rice. Plant Sci. 2016, 253, 40–49. [Google Scholar] [CrossRef]
- Bahaji, A.; Baroja, F.E.; Ricarte, B.A.; Sánchez, L.Á.M.; Muñoz, F.; Romero, J.M.; Ruiz, M.T.; Baslam, M.; Almagro, G.; Sesma, M.T.; et al. Characterization of multiple SPS knockout mutants reveals redundant functions of the four Arabidopsis sucrose phosphate synthase isoforms in plant viability, and strongly indicates that enhanced respiration and accelerated starch turnover can alleviate the blockage of sucrose biosynthesis. Plant Sci. 2015, 238, 135–147. [Google Scholar] [PubMed]
- Hirose, T.; Hashida, Y.; Aoki, N.; Okamura, M.; Yonekura, M.; Ohto, C.; Terao, T.; Ohsugi, R. Analysis of gene-disruption mutants of a sucrose phosphate synthase gene in rice, OsSPS1, shows the importance of sucrose synthesis in pollen germination. Plant Sci. 2014, 225, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Reimholz, R.; Deiting, U.; Sonnewald, U.; Stitt, M. Decreased expression of sucrose phosphate synthase strongly inhibits the water stress-induced synthesis of sucrose in growing potato tubers. Plant J. 1999, 19, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, K.; Hirotsu, N.; Kashiwagi, T.; Madoka, Y.; Nagasuga, K.; Ono, K.; Ohsugi, R. Overexpression of a maize SPS gene improves yield characters of potato under field conditions. Plant Prod. Sci. 2008, 11, 104–107. [Google Scholar] [CrossRef]
- Li, L.Q.; Zou, X.; Deng, M.S.; Peng, J.; Huang, X.L.; Lu, X.; Fang, C.C.; Wang, X.Y. Comparative morphology, transcription, and proteomics study revealing the key molecular mechanism of camphor on the potato tuber sprouting effect. Int. J. Mol. Sci. 2017, 18, 2280. [Google Scholar] [CrossRef]
- Sudhir, K.; Glen, S.; Koichiro, T. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Li, L.Q.; Deng, M.S.; Lyu, C.C.; Zhang, J.; Peng, J.; Cai, C.C.; Yang, S.M.; Lu, L.M.; Ni, S.; Liu, F.; et al. Quantitative phosphoproteomics analysis reveals that protein modification and sugar metabolism contribute to sprouting in potato after BR treatment. Food Chem. 2020, 325, 126875. [Google Scholar] [CrossRef]
- Deng, M.S.; Peng, J.; Zhang, J.; Ran, S.; Cai, C.C.; Yu, L.P.; Ni, S.; Huang, X.L.; Li, L.Q.; Wang, X.Y. The cysteine-rich peptide snakin-2 negatively regulates tubers sprouting through modulating lignin biosynthesis and H2O2 accumulation in potato. Int. J. Mol. Sci. 2021, 22, 2287. [Google Scholar] [CrossRef]
- Langenkämper, G.; Fung, R.W.M.; Newcomb, R.D.; Atkinson, R.G.; Gardner, R.C.; Macrae, E.A. Sucrose phosphate synthase genes in plants belong to three different families. J. Mol. Evol. 2002, 54, 322–332. [Google Scholar] [CrossRef]
- Duan, Y.; Yang, L.; Zhu, H.; Zhou, J.; Gong, H. Structure and expression analysis of sucrose phosphate synthase, sucrose synthase and invertase gene families in solanum lycopersicum. Int. J. Mol. Sci. 2021, 22, 4698. [Google Scholar] [CrossRef] [PubMed]
- Sonnewald, U. Control of potato tuber sprouting. Trends Plant Sci. 2001, 6, 333–335. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, K.; Ono, K.; Kashiwagi, T. Identification of a new gene controlling plant height in rice using the candidate-gene strategy. Planta 2004, 218, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Hashida, Y.; Aoki, N.; Kawanishi, H.; Okamura, M.; Ebitani, T.; Hirose, T.; Yamagishi, T.; Ohsugi, R. A near isogenic line of rice carrying chromosome segments containing OsSPS1 of Kasalath in the genetic background of Koshihikari produces anincreased spikelet number per panicle. Field Crops Res. 2013, 149, 56–62. [Google Scholar] [CrossRef]
- Bruneau, J.M.; Worrell, A.C.; Cambou, B.; Lando, D.; Voelker, T.A. Sucrose phosphate synthase, a key enzyme for sucrose biosynthesis in plants: Protein purification from corn leaves and immunological detection. Plant Physiol. 1991, 96, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Sonnewald, U.; Quick, W.P.; Macrae, E.; Krause, K.P.; Stitt, M. Purification, cloning and expression of spinach leaf sucrose-phosphate synthase in Escherichia Coli. Planta 1993, 189, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.P.; Zhang, X.T.; Chen, L.P.; Zhao, Q.; Zhang, Q.; Hua, X.T.; Wang, Z.C.; Tang, H.B.; Yu, Q.Y.; Zhang, M.Q.; et al. Comparative analysis of sucrose phosphate synthase (SPS) gene family between Saccharum officinarum and Saccharum spontaneum. BMC Plant Biol. 2020, 20, 422. [Google Scholar] [CrossRef]
- Reimholz, R.; Geiger, M.; Haake, V.; Deiting, U.; Krause, K.P.; Sonnewald, U.; Stitt, M. Potato plants contain multiple forms of sucrose phosphate synthase, which differ in their tissue distributions, their levels during development, and their responses to low temperature. Plant Cell Environ. 1997, 20, 291–305. [Google Scholar] [CrossRef]
- Solís, G.M.G.; Argüello, A.G.; López, B.J.; Ruiz, H.R.L.F.; López, M.J.E.; Sánchez, C.L.; Carreón, A.Y.; Martínez, T.M. Arabidopsis thaliana sucrose phosphate synthase(sps) genes are expressed differentially in organs and tissues, and their transcription is regulated by osmotic stress. Gene Expr. Patterns 2017, 25, 92–101. [Google Scholar] [CrossRef]
- Okamura, M.; Aoki, N.; Hirose, T.; Yonekura, O.C.; Ohsugi, R. Tissue specificity and diurnal change in gene expression of the sucrose phosphate synthase gene family in rice. Plant Sci. 2011, 181, 159–166. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer | Purpose |
|---|---|---|
| StSPS1 | qStSPS1-F: CAAGCTCAGTGGATGGCAAA | qRT-PCR |
| qStSPS1-R: TGACTGACCCATGCTTCCAT | ||
| StSPS2 | qStSPS2-F: AAGGGGGTTTTCATCAATCC | |
| qStSPS2-R: GCTTCAAAAGTGCATCAGCA | ||
| StSPS3 | qStSPS3-F: AGGAGTCAATTGCCATGGTC | |
| qStSPS3-R: GAAAACGCATGACCTCAGAC | ||
| StSPS4 | qStSPS4-F: ATCTGATGCTTTTGGCTGCT | |
| qStSPS4-R: TAAGGCCACACTGCTTTTCC | ||
| EF1αL | qEF1αL-F: CTTGTACACCACGCTAAGGAG | |
| qEF1αL-R: GTCAATGCAAACCATTCCTTG |
| Gene | PGSC Number | Chromosome Locus | Coding Sequence | EBI Number |
|---|---|---|---|---|
| StSPS1 | Soltu.DM.07G003160.1 | 7: 3688426–3697173 | 3165/1054 | M1CPB7/Q43845 |
| StSPS2 | Soltu.DM.08G010240.1 | 8: 30404288–30393496 | 3120/1039 | M1CXH8 |
| StSPS3 | Soltu.DM.09G029230.1 | 9: 65340125–65331377 | 3195/1064 | M1CI66 |
| StSPS4 | Soltu.DM.11G017190.1 | 11: 33423533–33413792 | 2598/865 | M1BN62/M1BN65 |
| Gene | MW (kD) | PIs | Gravy | Instability Index | +/− Residues | Localization |
|---|---|---|---|---|---|---|
| StSPS1 | 118.43 | 6.02 | −0.428 | 46.83 | 133/147 | Nucleus |
| StSPS2 | 116.68 | 6.19 | −0.374 | 45.72 | 132/144 | Nucleus |
| StSPS3 | 119.57 | 6.15 | −0.496 | 47.38 | 140/152 | Nucleus |
| StSPS4 | 96.48 | 6.45 | −0.239 | 45.67 | 101/107 | Chloroplast |
| Species | Gene Name | Number in Original Literature | Literature |
|---|---|---|---|
| Arabidopsis thaliana | AtSPS1/4/5.1/5.2 | AAF40445/CAB39764 | [30] |
| AAK64015/CAC03459 | |||
| Solanum lycopersicum | SlSPS1/2/3/4 | Solyc07g007790/Solyc08g042000 | [31] |
| Solyc09g092130/Solyc09g092130 | |||
| Nicotiana tabacum | NtSPS1/2/3 | AAF06792/ABA64521/ABA64520 | [16] |
| Oryza sativa | OsSPS1/2/6/8/11 | Rice SPS1/Rice SPS2/Rice SPS6 | [15] |
| Rice SPS8/Rice SPS11 | |||
| Triticum aestivum | TaSPS1/2/3/4/5 | Wheat SPS1a/Wheat SPS2a/Wheat SPS3a | |
| Wheat SPS4a/Wheat SPS5a |
| Gene | Number | RPKM Value in Transcriptome | Protein Expression in Proteome | |
|---|---|---|---|---|
| Dormancy | Sprouting | Sprouting | ||
| StSPS1 | PGSC0003DMT400071807 | 124.72 | 111.79 | 9139.23 |
| M1CPB7 | ||||
| StSPS2 | PGSC0003DMT400076855 | NA | 0.12 | N/A |
| M1CXH8 | ||||
| StSPS3 | PGSC0003DMT400067951 | 14.15 | 5.12 | 1305.77 |
| M1CI66 | ||||
| StSPS4 | PGSC0003DMT400049042 | 2.86 | 1.60 | N/A |
| M1BN62 | ||||
| PGSC0003DMT400049047 | 2.65 | 2.27 | N/A | |
| M1BN65 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, C.; Liu, S.; Liu, J.; Wen, H.; Li, L.; Wang, Q.; Li, L.; Wang, X. Screening and Identification of Potato StSPS1, a Potential Crucial Gene Regulating Seed Potato Vigor. Horticulturae 2023, 9, 511. https://doi.org/10.3390/horticulturae9040511
Cai C, Liu S, Liu J, Wen H, Li L, Wang Q, Li L, Wang X. Screening and Identification of Potato StSPS1, a Potential Crucial Gene Regulating Seed Potato Vigor. Horticulturae. 2023; 9(4):511. https://doi.org/10.3390/horticulturae9040511
Chicago/Turabian StyleCai, Chengcheng, Shifeng Liu, Jie Liu, He Wen, Luopin Li, Qiang Wang, Liqin Li, and Xiyao Wang. 2023. "Screening and Identification of Potato StSPS1, a Potential Crucial Gene Regulating Seed Potato Vigor" Horticulturae 9, no. 4: 511. https://doi.org/10.3390/horticulturae9040511
APA StyleCai, C., Liu, S., Liu, J., Wen, H., Li, L., Wang, Q., Li, L., & Wang, X. (2023). Screening and Identification of Potato StSPS1, a Potential Crucial Gene Regulating Seed Potato Vigor. Horticulturae, 9(4), 511. https://doi.org/10.3390/horticulturae9040511