
Effect of Wounding Intensity on Edible Quality by Regulating Physiological and ROS Metabolism in Fresh-Cut Pumpkins


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Treatment
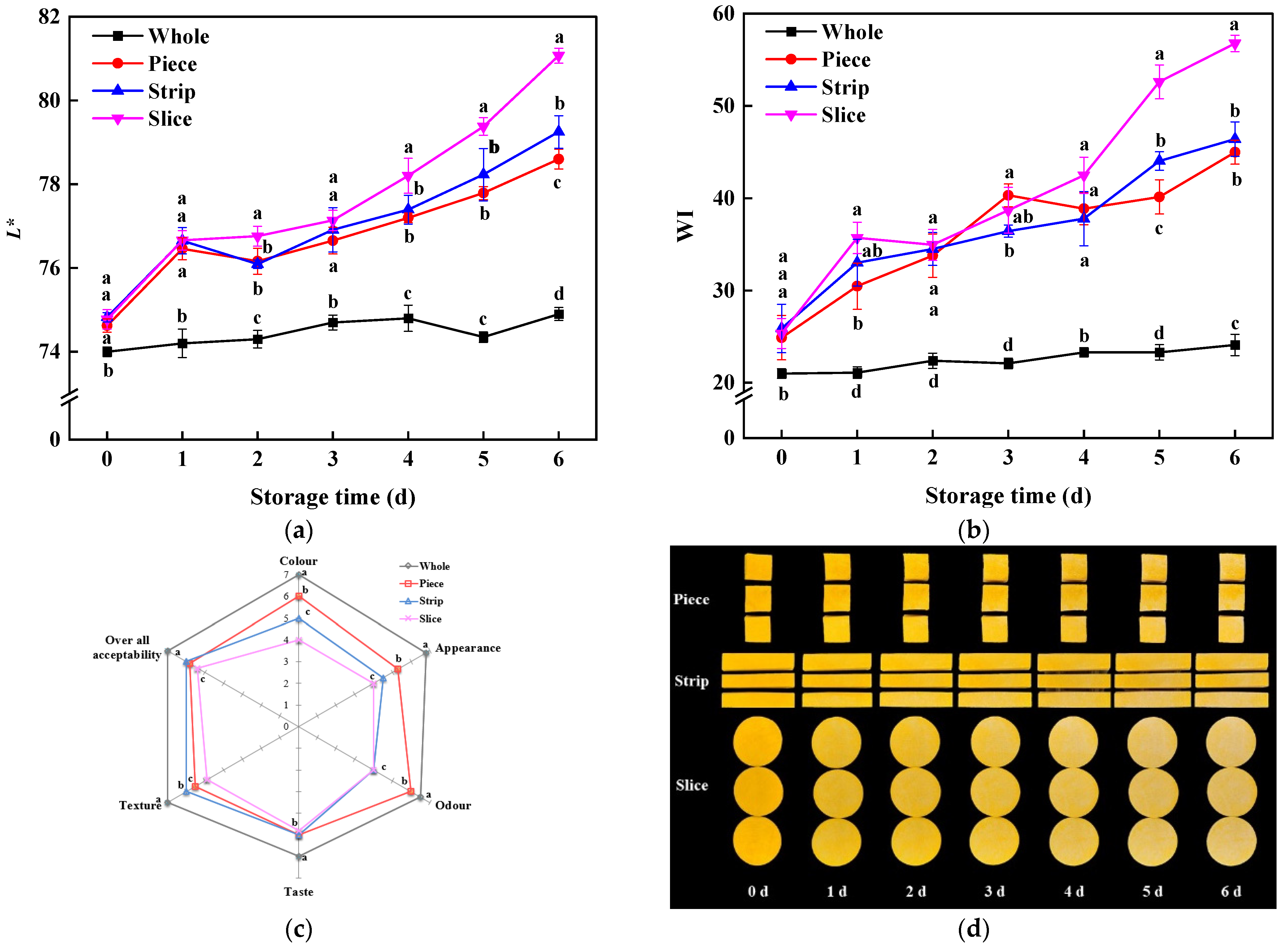
2.2. Lightness, Whiteness Index and Appearance Assay
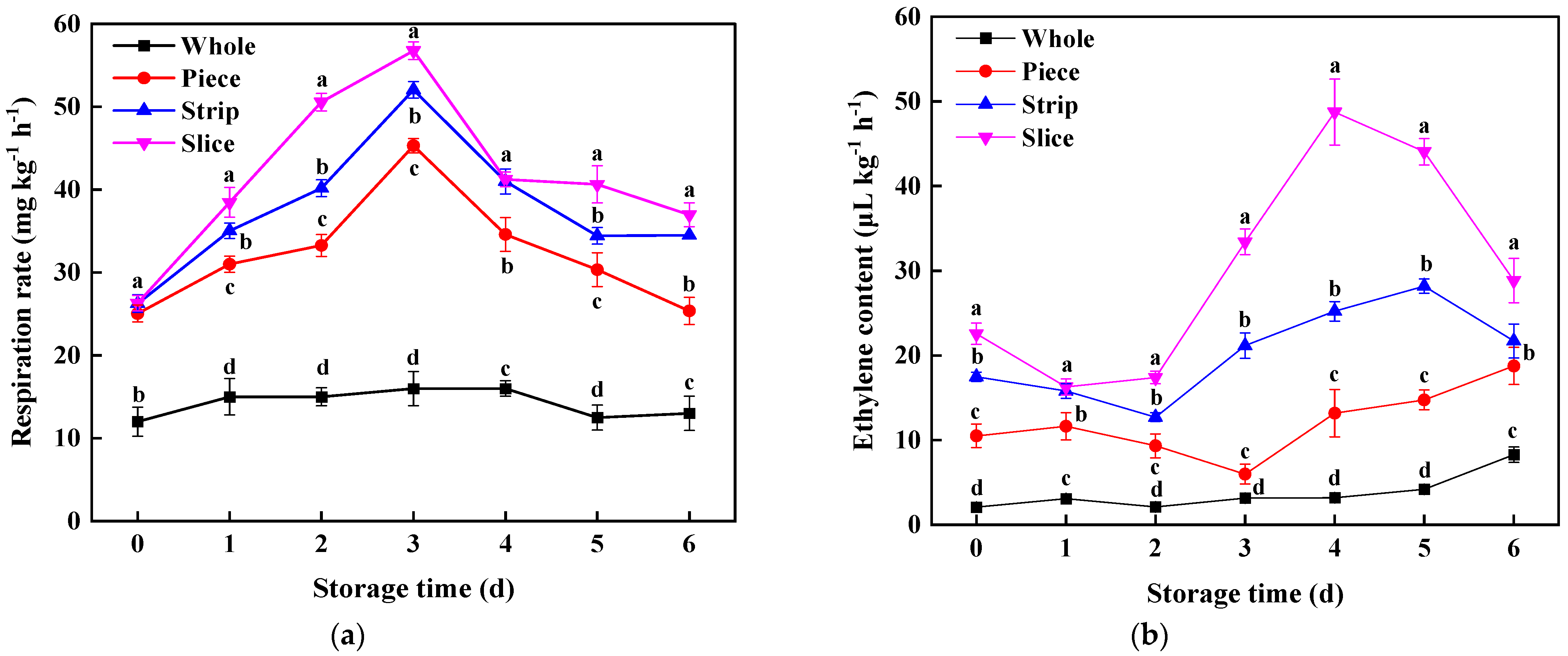
2.3. Respiration Rate and Ethylene Content Assay
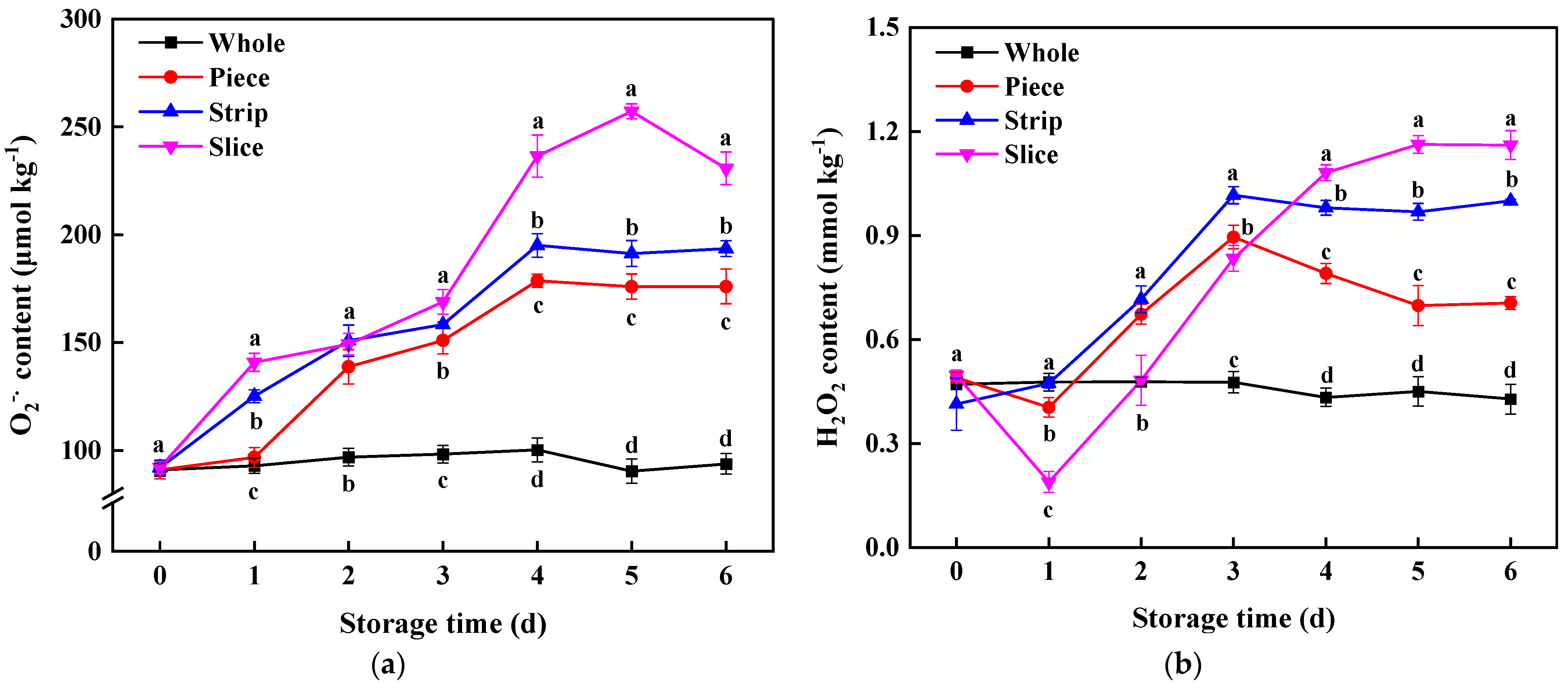
2.4. O2−• and H2O2 Content Assay
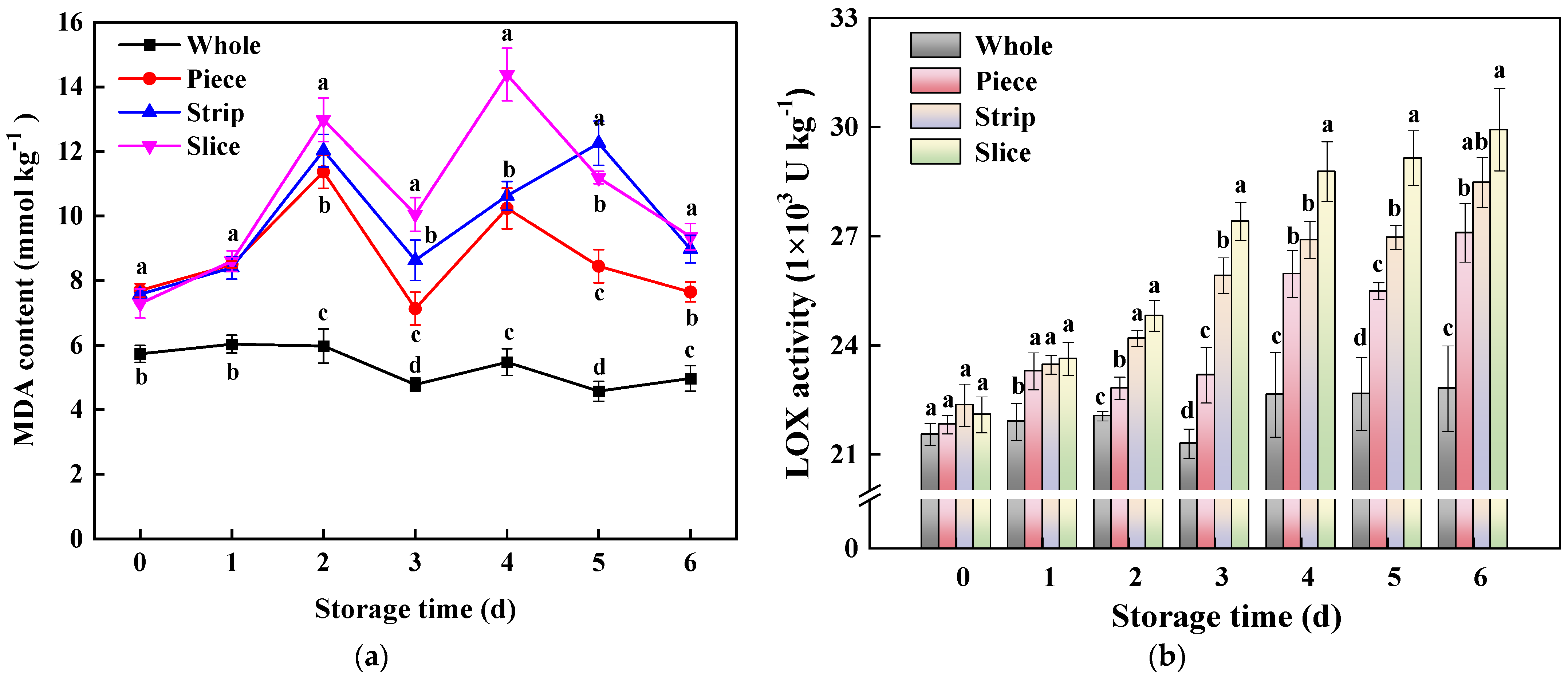
2.5. Malondialdehyde (MDA) Content and Lipoxygenase (LOX) Activity Assay
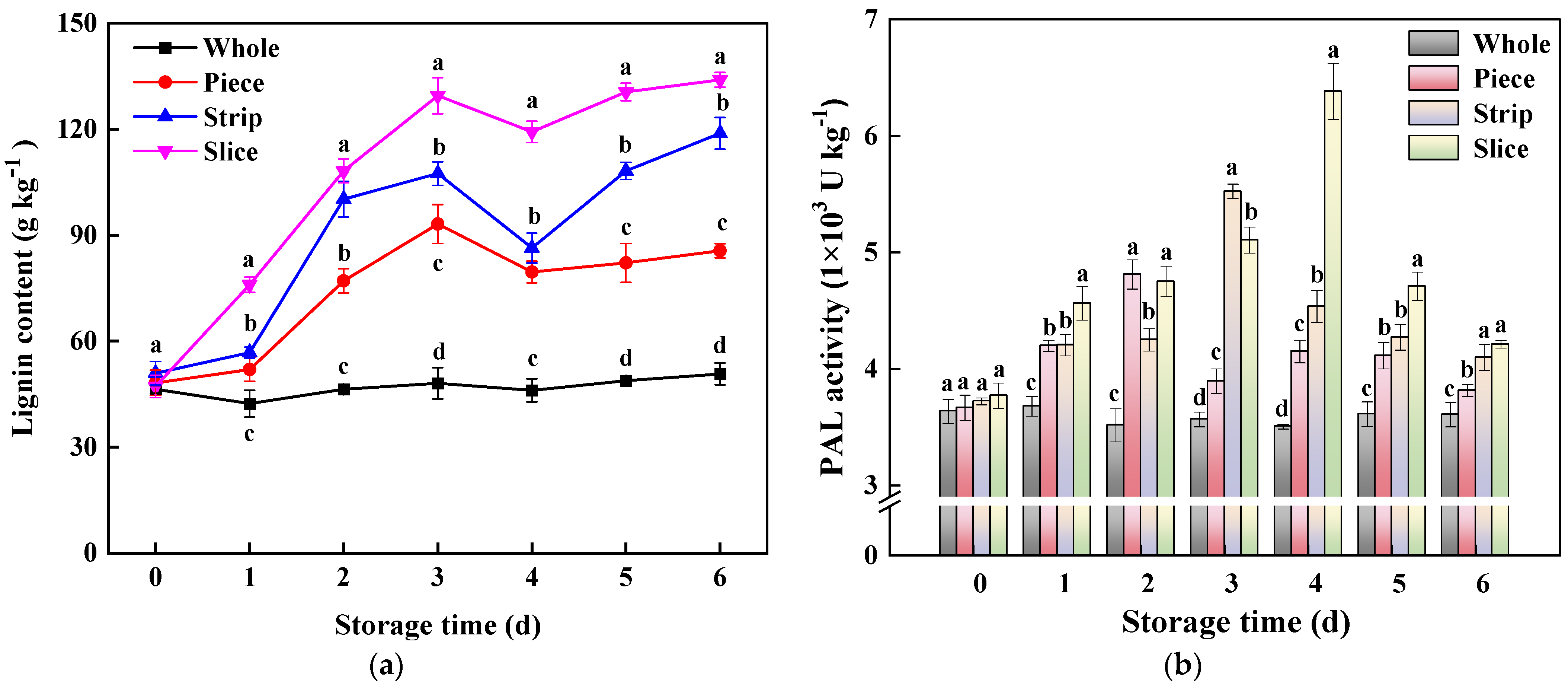
2.6. Lignin Content and Phenylalanine Ammonia-Lyase (PAL) Activity Assay
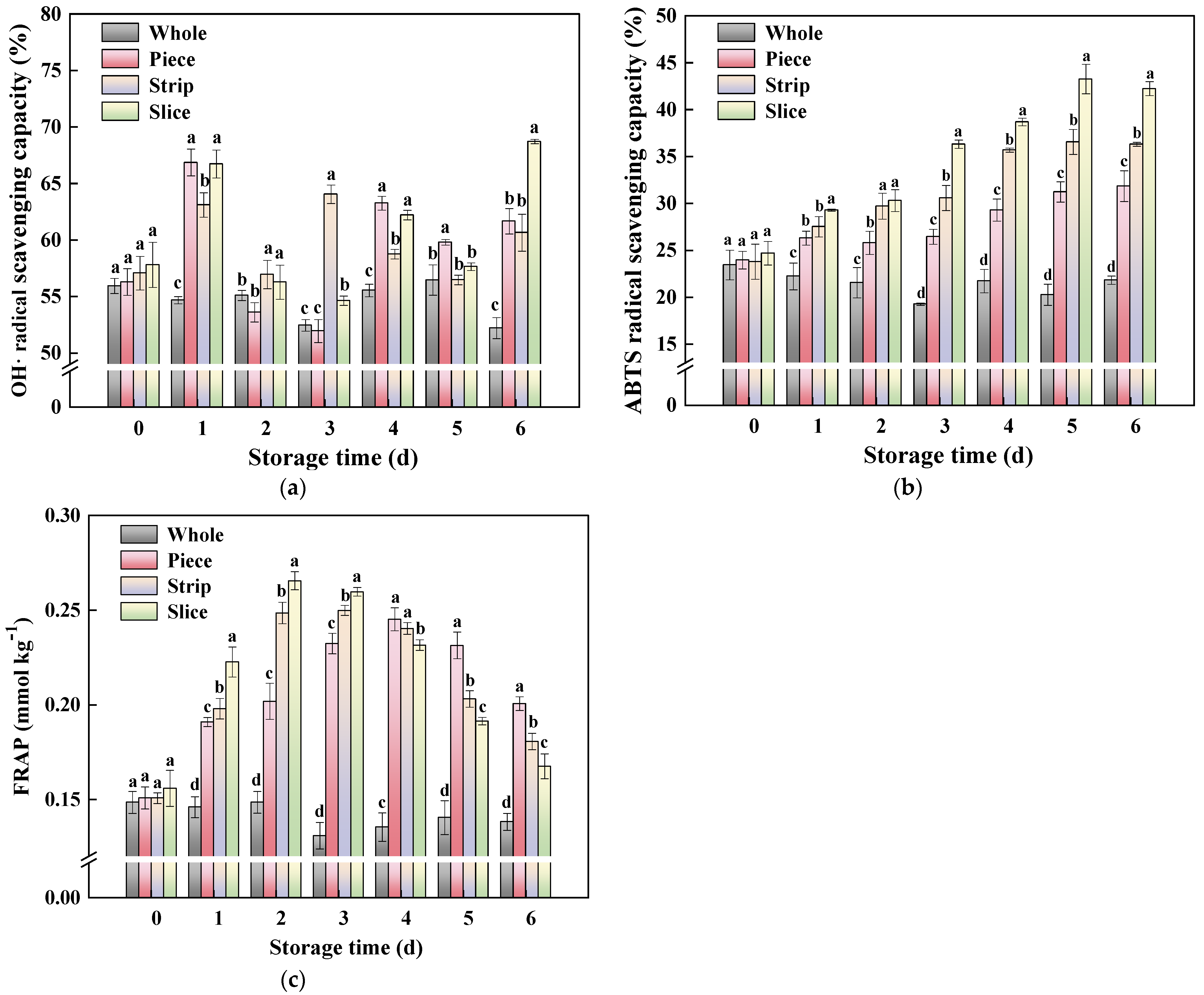
2.7. Antioxidant Activity Assay
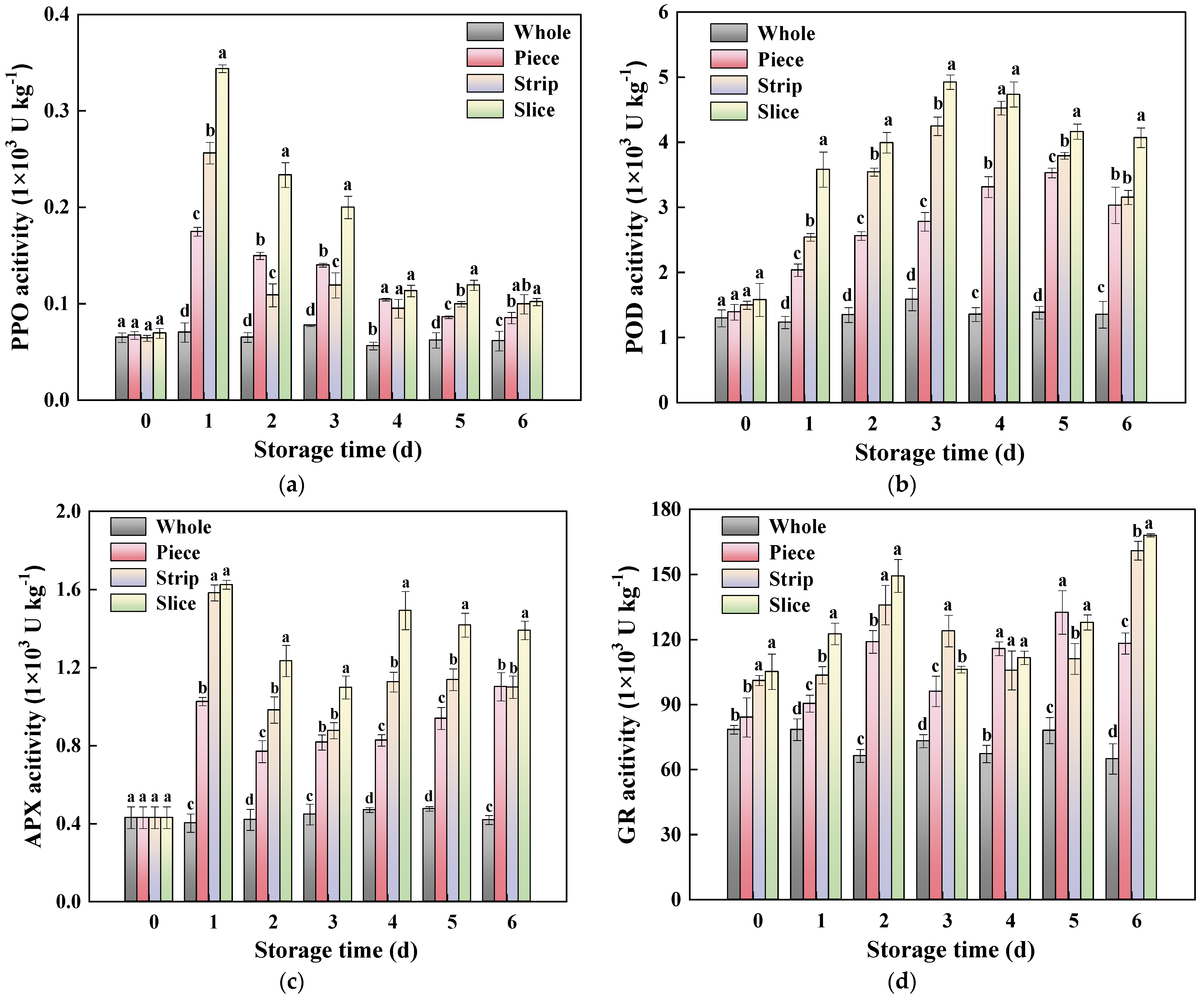
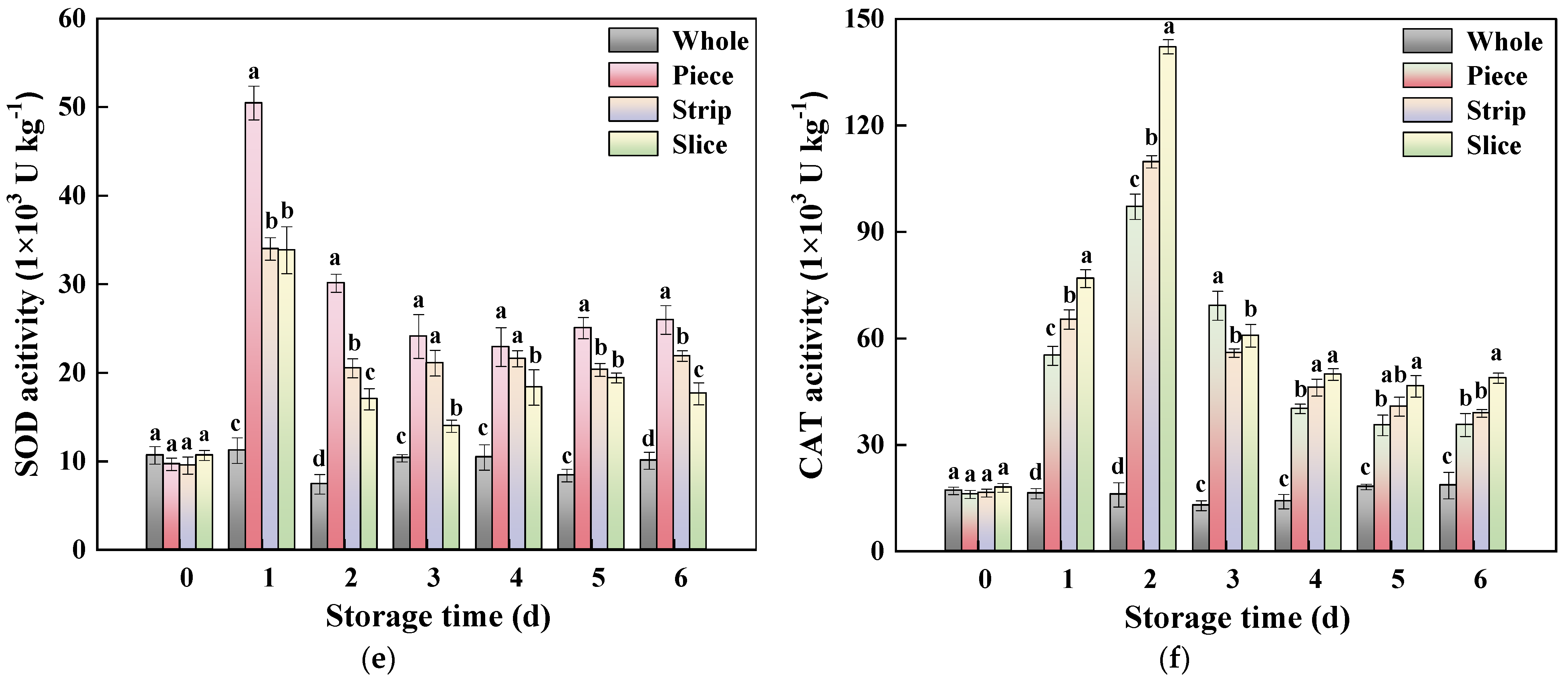
2.8. Antioxidant Enzyme Activity Assay
2.9. Antioxidant Compounds and Total Soluble Solid (TSS) Content Assay
2.10. Statistical Analysis
3. Results and Discussion
3.1. Colorimetric Values and Appearance
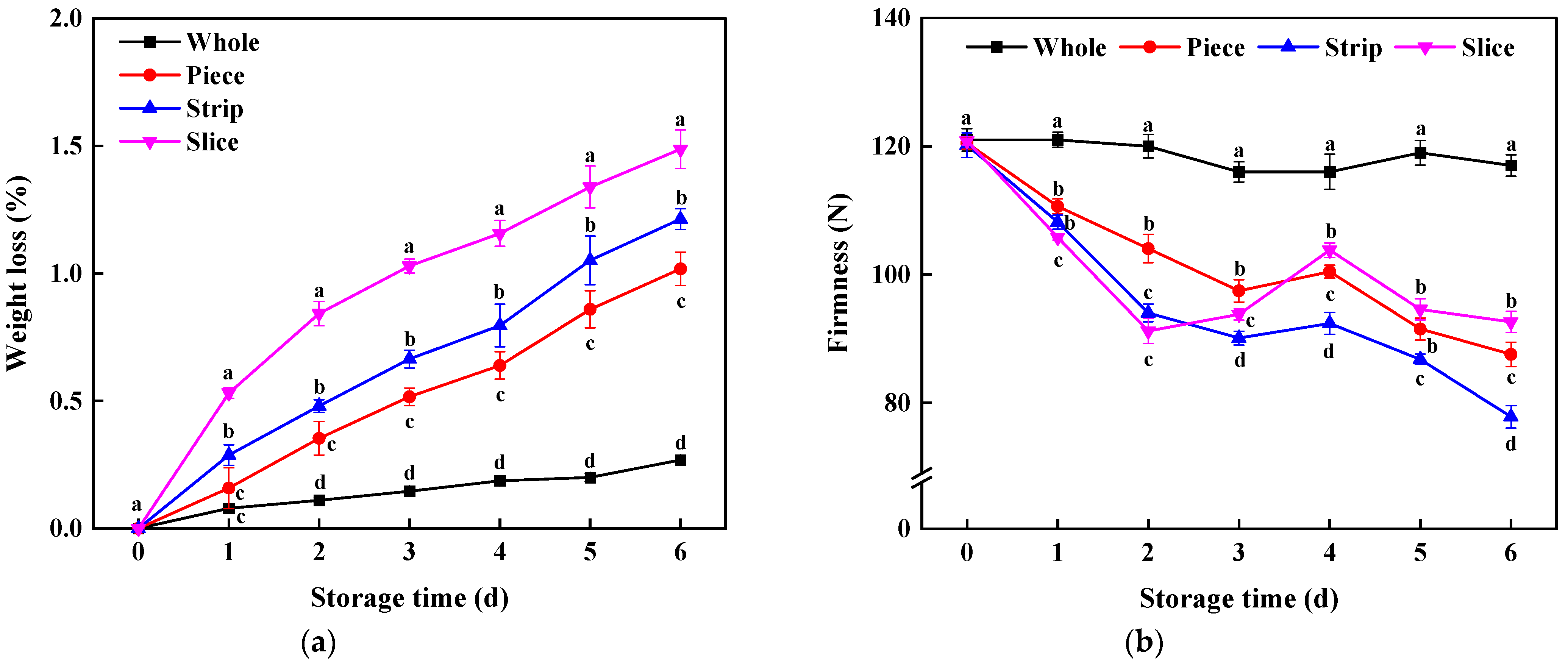
3.2. Weight Loss and Total Soluble Solid (TSS) Content
3.3. Effect of Wounding Intensities on Physiological Metabolism of Fresh-Cut Pumpkins
3.3.1. Respiration Rate and Ethylene Content
3.3.2. Phenylpropane Metabolism-Related Parameters
3.3.3. Membrane Lipid Peroxidation-Related Parameters
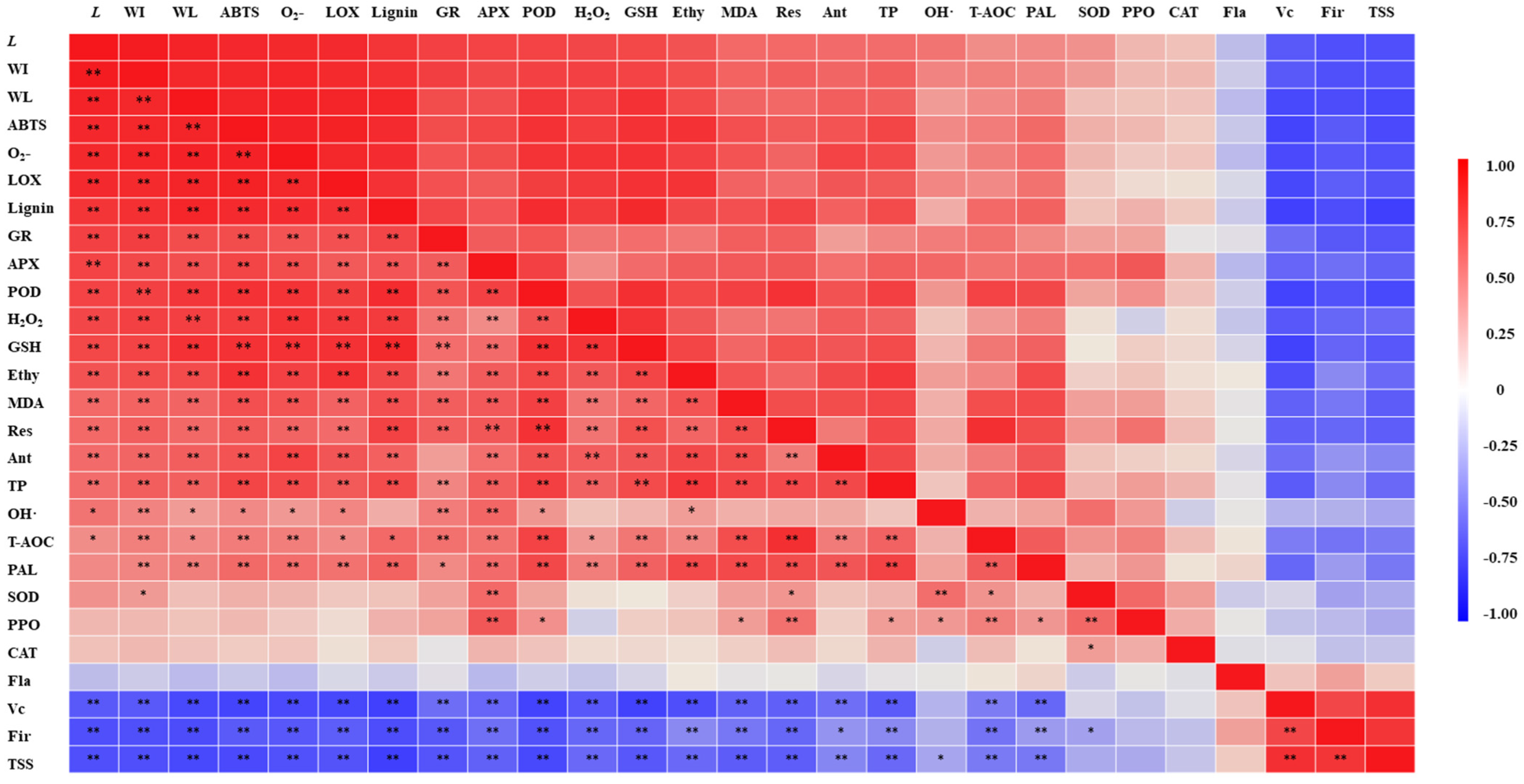
3.4. Effect of Wounding Intensities on ROS Metabolism of Fresh-Cut Pumpkins
3.4.1. ROS Content
3.4.2. Antioxidant Compound Content
3.4.3. ROS Metabolism-Related Enzyme Activity
3.4.4. Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liang, J.Y.; Guo, F.; Cao, S.F.; Zhao, K.; Zhao, K.X.; Wang, H.F.; Shao, X.F.; Wei, Y.Y.; Zhang, C.D.; Zheng, Y.H.; et al. γ-aminobutyric acid (GABA) alleviated oxidative damage and programmed cell death in fresh-cut pumpkins. Plant Physiol. Biochem. 2022, 180, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. Controlled abiotic stresses revisited: From homeostasis through hormesis to extreme stresses and the impact on nutraceuticals and quality during pre- and postharvest applications in horticultural crops. J. Agric. Food Chem. 2020, 68, 11877–11879. [Google Scholar] [CrossRef] [PubMed]
- Gade, S.R.; Meghwal, M.; Prabhakar, P.K.; Giuffre, A.M. A comparative study on the nutritional, antioxidant, thermal, morphological and diffraction properties of selected cucurbit seeds. Agronomy 2022, 12, 2242. [Google Scholar] [CrossRef]
- Valdez-Arjona, L.P.; Ramírez-Mella, M. Pumpkin waste as livestock feed: Impact on nutrition and animal health and on quality of meat, milk, and egg. Animals 2019, 9, 769. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.Z.; Sarengaowa; Guan, Y.G.; Feng, K. Biosynthesis of phenolic compounds and antioxidant activity in fresh-cut fruits and vegetables. Front. Microbiol. 2022, 13, 906069. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.G.; Hu, W.Z.; Jiang, A.L.; Xu, Y.P.; Yu, J.X.; Zhao, M.R.; Ji, Y.R.; Feng, K.; Sa, R.G.W.; Yang, X.Z. Influence of cut type on quality, antioxidant substances and antioxidant activity of fresh-cut broccoli. Int. J. Food Sci. Technol. 2020, 55, 3019–3030. [Google Scholar] [CrossRef]
- Han, C.; Ji, Y.; Li, M.L.; Li, X.A.; Jin, P.; Zheng, Y.H. Influence of wounding intensity and storage temperature on quality and antioxidant activity of fresh-cut Welsh onions. Sci. Hortic. 2016, 212, 203–209. [Google Scholar] [CrossRef]
- Li, X.A.; Long, Q.H.; Gao, F.; Han, C.; Jin, P.; Zheng, Y.H. Effect of cutting styles on quality and antioxidant activity in fresh-cut pitaya fruit. Postharvest Biol. Technol. 2017, 124, 1–7. [Google Scholar] [CrossRef]
- Han, C.; Jin, P.; Li, M.L.; Wang, L.; Zheng, Y.H. Physiological and transcriptomic analysis validates previous findings of changes in primary metabolism for the production of phenolic antioxidants in wounded carrots. J. Agric. Food Chem. 2017, 65, 7159–7167. [Google Scholar] [CrossRef]
- Chai, Z.Y.; Zhang, F.; Liu, B.J.; Chen, X.F.; Meng, X.H. Antibacterial mechanism and preservation effect of curcumin-based photodynamic extends the shelf life of fresh-cut pears. LWT-Food Sci. Technol. 2021, 142, 110941. [Google Scholar] [CrossRef]
- Hu, W.Z.; Guan, Y.G.; Ji, Y.R.; Yang, X.Z. Effect of cutting styles on quality, antioxidant activity, membrane lipid peroxidation, and browning in fresh-cut potatoes. Food Biosci. 2021, 44, 101435. [Google Scholar]
- Han, C.; Li, J.; Jin, P.; Li, X.; Wang, L.; Zheng, Y. The effect of temperature on phenolic content in wounded carrots. Food Chem. 2017, 215, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.G.; Hu, W.Z.; Jiang, A.L.; Xu, Y.P.; Zhao, M.R.; Yu, J.X.; Ji, Y.R.; Sa, R.G.W.; Yang, X.Z.; Feng, K. The effect of cutting style on the biosynthesis of phenolics and cellular antioxidant capacity in wounded broccoli. Food Res. Int. 2020, 137, 109565. [Google Scholar] [CrossRef]
- Sarengaowa; Hu, W.Z.; Jiang, A.L.; Xiu, Z.L.; Feng, K. Effect of thyme oil-alginate based coating on quality and microbial safety of fresh-cut apples. J. Sci. Food Agric. 2018, 98, 2302–2311. [Google Scholar] [CrossRef]
- Zhou, F.H.; Xu, D.Y.; Liu, C.H.; Chen, C.; Tian, M.X.; Jiang, A.L. Ascorbic acid treatment inhibits wound healing of fresh-cut potato strips by controlling phenylpropanoid metabolism. Postharvest Biol. Technol. 2021, 181, 111644. [Google Scholar] [CrossRef]
- Coming Biotechnology Company Home Page of MDA Content Kit. Available online: http://www.cominbio.com/a/shijihe/shenghuashiji/yanghuayukangyanghuaxilie/2018/0708/109.html (accessed on 15 November 2022).
- Coming Biotechnology Company Home Page of LOX Activity Kit. Available online: http://www.cominbio.com/a/shijihe/shenghuashiji/zhifangsuandaixiexilie/2014/1112/509.html (accessed on 25 November 2022).
- Coming Biotechnology Company Home Page of Lignin Content Kit. Available online: http://www.cominbio.com/a/shijihe/shenghuashiji/qitaxilie/2016/0502/969.html (accessed on 3 November 2022).
- Jiancheng Bioengineering Institute Company Home Page of OH. Scavenging Ability Kit. Available online: http://www.njjcbio.com/products.asp?id=318 (accessed on 20 November 2022).
- Jiancheng Bioengineering Institute Company Home Page of ABTS Scavenging Ability Kit. Available online: http://www.njjcbio.com/product.asp?commend=1&q=ABTS&sort=hitsorder (accessed on 20 November 2022).
- Jiancheng Bioengineering Institute Company Home Page of DPPH Scavenging Ability Kit. Available online: http://www.njjcbio.com/product.asp?commend=1&q=FRAP&sort=hitsorder (accessed on 20 November 2022).
- Tian, S.P.; Xu, Y.; Jiang, A.; Gong, Q.Q. Physiological and quality responses of longan fruit to high O2 or high CO2 atmospheres in storage. Postharvest Biol. Technol. 2002, 24, 335–340. [Google Scholar] [CrossRef]
- Chen, C.; Hu, W.; He, Y.; Jiang, A.; Zhang, R. Effect of citric acid combined with UV-C on the quality of fresh-cut apples. Postharvest Biol. Technol. 2016, 111, 126–131. [Google Scholar] [CrossRef]
- Coming Biotechnology Company Home Page of SOD Activity Kit. Available online: http://www.cominbio.com/a/shijihe/shenghuashiji/yanghuayukangyanghuaxilie/2019/0411/1795.html) (accessed on 18 December 2022).
- Coming Biotechnology Company Home Page of APX Activity Kit. Available online: http://www.cominbio.com/a/shijihe/shenghuashiji/weishengsuCdaixiexilie/2014/1111/439.html (accessed on 28 December 2022).
- Coming Biotechnology Company Home Page of GR Activity Kit. Available online: http://www.cominbio.com/a/shijihe/shenghuashiji/guguanggataixilie/2014/1111/432.html (accessed on 28 December 2022).
- Guan, Y.G.; Ji, Y.R.; Yang, X.Z.; Pang, L.J.; Cheng, J.Y.; Lu, X.H.; Zheng, J.; Yin, L.Q.; Hu, W.Z. Antioxidant activity and microbial safety of fresh-cut red cabbage stored in different packaging films. LWT-Food Sci. Technol. 2023, 17, 114478. [Google Scholar] [CrossRef]
- Song, Z.; Feng, L.; Hui, G.; Xu, Y.; Li, D. Combination of nisin and ε-polylysine with chitosan coating inhibits the white blush of fresh-cut carrots. Food Control. 2017, 74, 34–44. [Google Scholar] [CrossRef]
- Chen, C.C.; Xu, L.; Wang, S.W.; Chen, X.N.; Shen, Y.Y. Characterization of abscisic acid and ethylene in regulating the white blush in fresh-cut carrots. Int. J. Mol. Sci. 2022, 23, 12788. [Google Scholar] [CrossRef]
- Zhao, P.; Li, W.; Zhen, C.; Wang, K.; Qin, Z.; Gao, H. Transcriptomic analysis of the effect of γ-aminobutyric acid treatment on browning and induced disease resistance in fresh-cut apples. Postharvest Biol. Technol. 2021, 181, 111686. [Google Scholar] [CrossRef]
- Wang, L.; Guo, Y.Y.; Wang, X.Y.; Zhang, X.H. Short-term O2/CO2 controlled atmosphere altered the water status and thus promoted phenolic biosynthesis during wound healing of fresh-cut white mushroom (Agaricus bisporus). Postharvest Biol. Technol. 2022, 188, 111879. [Google Scholar] [CrossRef]
- Zhou, Z.C.; Han, P.; Bai, S.Q.; Ma, N.; Fang, D.L.; Yang, W.J.; Hu, Q.H.; Pei, F. Caffeic acid-grafted-chitosan/polylactic acid film packaging enhances the postharvest quality of Agaricus bisporus by regulating membrane lipid metabolism. Food Res. Int. 2022, 158, 111557. [Google Scholar] [CrossRef]
- Renna, M.; Pace, B.; Cefola, M.; Santamaria, P.; Serio, F.; Gonnella, M. Comparison of two jam-making methods to preserve the quality of colored carrots. LWT-Food Sci. Technol. 2013, 53, 547–554. [Google Scholar] [CrossRef]
- Lin, X.; Yang, R.; Dou, Y.; Zhang, W.; Du, H.; Zhu, L.; Chen, J. Transcriptome analysis reveals delaying of the ripening and cell-wall degradation of kiwifruit by hydrogen sulfide. J. Sci. Food Agric. 2020, 100, 2280–2287. [Google Scholar] [CrossRef]
- Baccio, D.D.; Ederli, L.; Marabottini, R.; Badiani, M.; Francini, A.; Nali, C.; Antonelli, M.; Santangelo, E.; Sebastiani, L.; Pasqualini, S. Similar foliar lesions but opposite hormonal patterns in a tomato mutant impaired in ethylene perception and its near isogenic wild type challenged with ozone. Environ. Exp. Bot. 2012, 75, 286–297. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Q.; Zhou, X.; Zhang, F.; Ji, S. Ethylene plays an important role in the softening and sucrose metabolism of blueberries postharvest. Food Chem. 2020, 310, 125965. [Google Scholar] [CrossRef]
- Zhao, T.; Nakano, A.; Iwasaki, Y. Differences between ethylene emission characteristics of tomato cultivars in tomato production at plant factory. J. Agric. Food Res. 2021, 5, 100181. [Google Scholar] [CrossRef]
- Fan, X.; Du, Z.; Cui, X.; Ji, W.; Gong, H. Preharvest methyl salicylate treatment enhance the chilling tolerance and improve the postharvest quality of apricot during low temperature storage. Postharvest Biol. Technol. 2021, 177, 111535. [Google Scholar] [CrossRef]
- Ferrer, J.L.; Austin, M.B.; Stewart, C.; Noel, J.P. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef]
- Jacobo-Velazquez, D.A.; Martinez-Hernandez, G.B.; Rodriguez, S.D.C.; Cao, C.M.; Cisneros-Zevallos, L. Plants as biofactories: Physiological role of reactive oxygen species on the accumulation of phenolic antioxidants in carrot tissue under wounding and hyperoxia stress. J. Agric. Food Chem. 2011, 59, 6583–6593. [Google Scholar] [CrossRef]
- Giuffre, A.M.; Zappia, C.; Capocasale, M. Physico-chemical stability of blood orange juice during frozen storage. Int. J. Food Prop. 2017, 20, 1930–1943. [Google Scholar]
- Mele, M.A. Bioactive compounds and biological activity of ginger. J. Multidiscip. Sci. 2019, 1, 1–7. [Google Scholar] [CrossRef]
- Fan, K.; Zhang, M.; Jiang, F.J. Ultrasound treatment to modified atmospheric packaged fresh-cut cucumber: Influence on microbial inhibition and storage quality. Ultrason. Sonochem. 2019, 54, 162–170. [Google Scholar] [CrossRef]
- Huang, H.; Guo, L.F.; Wang, L.; Wang, H.; Ma, S.M.; Jiang, Y.M.; Qu, H.X. 1-Methylcyclopropene (1-MCP) slows ripening of kiwifruit and affects energy status, membrane fatty acid contents and cell membrane integrity. Postharvest Biol. Technol. 2019, 156, 110941. [Google Scholar] [CrossRef]
- Mao, L.C.; Pang, H.Q.; Wang, G.; Zhu, C.G. Phospholipase D and lipoxygenase activity of cucumber fruit in response to chilling stress. Postharvest Biol. Technol. 2007, 44, 42–47. [Google Scholar] [CrossRef]
- Hu, K.D.; Wang, Q.; Hu, L.Y.; Gao, S.P.; Wu, J.; Li, Y.H. Hydrogen sulfide prolongs postharvest storage of fresh-cut pears (Pyrus pyrifolia) by alleviation of oxidation of oxidative damage and inhibition of fungal growth. PLoS ONE 2014, 9, e85524. [Google Scholar]
- Gao, H.; Chai, H.K.; Cheng, N.; Cao, W. Effects of 24-epibrassinolide on enzymatic browning and antioxidant activity of fresh-cut lotus root slices. Food Chem. 2017, 217, 45–51. [Google Scholar] [CrossRef]
- You, Y.L.; Jiang, Y.M.; Sun, J.; Liu, H.; Song, L.L.; Duan, X.W. Effects of short-term anoxia treatment on browning of fresh-cut Chinese water chestnut in relation to antioxidant activity. Food Chem. 2012, 132, 1191–1196. [Google Scholar] [CrossRef]
- Guan, Y.G.; Hu, W.Z.; Xu, Y.P.; Yang, X.Z.; Ji, Y.R.; Feng, K.; Sa, R.G.W. Metabolomics and physiological analyses validates previous findings on the mechanism of response to wounding stress of different intensities in broccoli. Food Res. Int. 2020, 140, 110058. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, D.C.; Chen, T.; Li, B.Q.; Zhang, Z.Q.; Qin, G.Z.; Tian, S.P. Production, signaling, and scavenging mechanisms of reactive oxygen species in fruit-pathogen interactions. Int. J. Mol. Sci. 2019, 20, 2994. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.F.; Lu, L.; Su, M.Y.; Zhang, J.; Li, Y.Y.; Huang, S.L.; Li, C.X.; Wang, N.; Zhang, Z.Y.; Chen, X.S. Analysis of differentially expressed genes and differentially abundant metabolites associated with the browning of Meihong red-fleshed apple fruit. Postharvest Biol. Technol. 2021, 174, 111437. [Google Scholar] [CrossRef]
- He, Q.; Xu, Y.H.; Wang, Y.X.; Zhang, N.N.; Zhang, L.G. Metabolic profiling and transcriptomic data providing critical flavonoid biosynthesis mechanisms disclose color differences of purple heading Chinese cabbages (Brassica rapa L.). LWT-Food Sci. Technol. 2022, 168, 113885. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Come, D. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiol. Plant. 2010, 97, 104–110. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, L.; Liu, W.; Lin, Q.; Wang, Z.; Guan, W. Effects of UV-C on antioxidant capacity, antioxidant enzyme activity and color of fresh-cut red cabbage during storage. Int. J. Food Sci. Technol. 2016, 52, 626–634. [Google Scholar] [CrossRef]
- Mori, T.; Terai, H.; Yamauchi, N.; Suzuki, Y. Effects of postharvest ethanol vapor treatment on the ascorbate-glutathione cycle in broccoli florets. Postharvest Biol. Technol. 2009, 52, 134–136. [Google Scholar] [CrossRef]
- Barrett, D.M.; Beaulieu, J.C.; Shewfelt, R. Color, flavor, texture, and nutritional quality of fresh-cut fruits and vegetables: Desirable levels, instrumental and sensory measurement, and the effects of processing. Crit. Rev. Food Sci. 2010, 50, 369–389. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.G.; Hu, W.Z.; Xu, Y.P.; Sarengaowa; Ji, Y.R.; Yang, X.Z.; Feng, K. Proteomic analysis validates previous findings on wounding-responsive plant hormone signaling and primary metabolism contributing to the biosynthesis of secondary metabolites based on metabolomic analysis in harvested broccoli (Brassica oleracea L. var. italica). Food Res. Int. 2021, 145, 110388. [Google Scholar]
- Torres-Contreras, A.M.; Senes-Guerrero, C.; Pacheco, A.; Gonzalez-Agüero, M.; Ramos-Parra, P.A.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. Genes differentially expressed in broccoli as an early and late response to wounding stress. Postharvest Biol. Technol. 2018, 145, 172–182. [Google Scholar] [CrossRef]
- Martinez-Hernandez, G.B.; Artés-Hernández, F.; Gómez, P.A.; Formica, A.C.; Artés, F. Combination of electrolysed water, UV-C, and super atmospheric O2; packaging for improving fresh-cut broccoli quality. Postharvest Biol. Technol. 2013, 76, 125–134. [Google Scholar] [CrossRef]
- Li, H.; Tian, S.P.; Qin, G.Z. NADPH oxidase is crucial for the cellular redox homeostasis in fungal pathogen Botrytis cinerea. Mol. Plant Microbe Interact. 2019, 32, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Song, J.L.; Huang, H.; Hao, Y.W.; Song, S.W.; Zhang, Y.T.; Su, W.; Liu, H.C. Nutritional quality, mineral and antioxidant content in lettuce afected by interaction of light intensity and nutrient solution concentration. Sci. Rep. 2020, 10, 2796. [Google Scholar] [CrossRef]
- Singh, S.; Dey, S.S.; Kumar, R.; Bhatia, R.; Ghemeray, H.; Behera, T.K. Genetic analysis and interaction among CUPRAC, FRAP, phytochemical and phenotypic traits in cauliflower (Brassica oleracea var. botrytis L.). Int. J. Chem. Stud. 2019, 7, 1484–1494. [Google Scholar]
- Purkiewicz, A.; Ciborska, J.; Tanska, M.; Narwojsz, A.; Starowicz, M.; Przybylowicz, K.E.; Sawicki, T. The impact of the method extraction and different carrot variety on the carotenoid profile, total phenolic content and antioxidant properties of juices. Plants 2020, 9, 1759. [Google Scholar] [CrossRef]
- Reyes, L.F.; Villarreal, J.E.; Cisneros-Zevallos, L. The increase in antioxidant capacity after wounding depending on the type of fruit or vegetable tissue. Food Chem. 2007, 101, 1254–1262. [Google Scholar]
- Surjadinata, B.B.; Cisneros-Zevallos, L. Biosynthesis of phenolic antioxidants in carrot tissue increases with wounding intensity. Food Chem. 2012, 134, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Vina, S.Z.; Chaves, A.R. Antioxidant responses in minimally processed celery during refrigerated storage. Food Chem. 2006, 94, 68–74. [Google Scholar] [CrossRef]
- Zhan, L.J.; Li, Y.; Hu, J.Q.; Pang, L.Y.; Fan, H.P. Browning inhibition and quality preservation of fresh-cut romaine lettuce exposed to high intensity light. Innov. Food Sci. Emerg. Technol. 2012, 14, 70–76. [Google Scholar] [CrossRef]
- Oms-Oliu, G.; Aguilo-Aguayo, I.; Martin-Belloso, O.; Soliva-Fortuny, R. Effects of pulsed light treatments on quality and antioxidant properties of fresh-cut mushrooms (Agaricus bisporus). Postharvest Biol. Technol. 2010, 56, 216–222. [Google Scholar] [CrossRef]
- Berno, N.D.; Tezotto-Uliana, J.V.; dos Santos Dias, C.T.; Kluge, R.A. Storage temperature and type of cut affect the biochemical and physiological characteristics of fresh-cut purple onions. Postharvest Biol. Technol. 2014, 93, 91–96. [Google Scholar] [CrossRef]
- Robles-Sanchez, M.R.; Alejandra Rojas-Gratue, M.; Odriozola-Serrano, I.; Gonzalez-Aguilar, G.; Martin-Belloso, O. Influence of alginate-based edible coating as carrier of antibrowning agents on bioactive compounds and antioxidant activity in fresh-cut kent mangoes. LWT-Food Sci. Technol. 2013, 50, 240–246. [Google Scholar] [CrossRef]
- Torres-Contreras, A.M.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. Plants as biofactories: Stress-induced production of chlorogenic acid isomers in potato tubers as affected by wounding intensity and storage time. Ind. Crops Prod. 2014, 62, 61–66. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Treatment | 0 d | 1 d | 2 d | 3 d | 4 d | 5 d | 6 d |
|---|---|---|---|---|---|---|---|---|
| TSS (%) | Whole | 16.00 ± 0.54 a | 15.70 ± 0.20 a | 15.30 ± 0.50 a | 15.61 ± 0.06 a | 15.13 ± 0.44 a | 15.52 ± 0.11 a | 16.03 ± 0.07 a |
| Piece | 15.65 ± 0.39 a | 13.81 ± 0.11 c | 12.45 ± 0.39 b | 14.57 ± 0.34 b | 14.23 ± 0.37 b | 12.97 ± 0.11 b | 12.54 ± 0.29 b | |
| Strip | 15.87 ± 0.13 a | 14.62 ± 0.40 b | 11.67 ± 0.19 c | 12.32 ± 0.33 c | 13.12 ± 0.29 c | 11.97 ± 0.13 c | 11.87 ± 0.15 c | |
| Slice | 15.45 ± 0.21 a | 13.52 ± 0.16 d | 11.55 ± 0.37 c | 10.75 ± 0.05 d | 12.22 ± 0.22 d | 11.07 ± 0.07 d | 10.43 ± 0.11 d | |
| Total phenols content (g kg−1) | Whole | 10.74 ± 0.42 c | 10.19 ± 0.25 c | 10.04 ± 0.11 c | 9.92 ± 0.23 d | 10.19 ± 0.44 d | 10.35 ± 0.22 d | 10.53 ± 0.22 c |
| Piece | 11.16 ± 0.26 b | 12.01 ± 0.19 b | 13.47 ± 0.21 a | 12.74 ± 0.26 c | 12.70 ± 0.38 c | 12.02 ± 0.29 c | 11.07 ± 0.32 b | |
| Strip | 11.32 ± 0.35 b | 12.32 ± 0.20 ab | 12.41 ± 0.23 b | 13.52 ± 0.21 b | 14.58 ± 0.33 b | 13.40 ± 0.22 b | 11.61 ± 0.23 b | |
| Slice | 12.07 ± 0.28 a | 12.71 ± 0.31 a | 13.92 ± 0.29 a | 15.74 ± 0.04 a | 16.30 ± 0.44 a | 18.54 ± 0.24 a | 12.49 ± 0.34 a | |
| Flavonoids content (g kg−1) | Whole | 5.95 ± 0.17 a | 5.46 ± 0.21 c | 5.57 ± 0.35 c | 5.89 ± 0.16 d | 5.45 ± 0.11 d | 5.49 ± 0.31 c | 5.58 ± 0.15 b |
| Piece | 5.83 ± 0.19 a | 7.60 ± 0.25 a | 7.58 ± 0.38 b | 8.40 ± 0.19 b | 8.97 ± 0.28 b | 7.89 ± 0.18 b | 7.08 ± 0.19 a | |
| Strip | 5.63 ± 0.44 a | 6.66 ± 0.12 b | 7.37 ± 0.16 b | 7.69 ± 0.14 c | 7.79 ± 0.16 c | 8.21 ± 0.11 b | 7.20 ± 0.16 a | |
| Slice | 5.98 ± 0.55 a | 7.65 ± 0.24 a | 8.46 ± 0.18 a | 9.10 ± 0.41 a | 11.04 ± 0.28 a | 8.57 ± 0.07 a | 7.44 ± 0.26 a | |
| Anthocyanin content (g kg−1) | Whole | 0.11 ± 0.04 a | 0.13 ± 0.01 c | 0.13 ± 0.03 c | 0.11 ± 0.01 c | 0.16 ± 0.01 d | 0.13 ± 0.04 c | 0.15 ± 0.01 c |
| Piece | 0.17 ± 0.03 a | 0.19 ± 0.02 ab | 0.21 ± 0.02 b | 0.24 ± 0.01 b | 0.55 ± 0.02 b | 0.24 ± 0.03 b | 0.22 ± 0.02 b | |
| Strip | 0.15 ± 0.01 a | 0.18 ± 0.02 b | 0.20 ± 0.03 ab | 0.28 ± 0.02 a | 0.35 ± 0.01 c | 0.45 ± 0.03 a | 0.21 ± 0.02 b | |
| Slice | 0.14 ± 0.02 a | 0.24 ± 0.03 a | 0.26 ± 0.03 a | 0.31 ± 0.01 a | 0.72 ± 0.01 a | 0.50 ± 0.02 a | 0.28 ± 0.03 a | |
| Vitamin C (mg kg−1) | Whole | 347.94 ± 1.55 a | 350.30 ± 11.28 a | 348.57 ± 5.77 a | 333.85 ± 1.46 a | 337.62 ± 11.41 a | 334.68 ± 2.08 a | 340.39 ± 1.42 a |
| Piece | 342.21 ± 10.40 a | 345.30 ± 10.00 a | 315.64 ± 5.13 b | 315.76 ± 8.62 b | 299.77 ± 7.64 b | 287.70 ± 3.54 b | 245.30 ± 2.03 b | |
| Strip | 344.06 ± 1.85 a | 275.20 ± 7.61 c | 230.15 ± 2.76 c | 218.55 ± 7.36 c | 222.85 ± 7.80 c | 231.59 ± 1.92 c | 228.97 ± 5.30 c | |
| Slice | 325.56 ± 3.11 b | 322.27 ± 2.96 b | 222.69 ± 5.04 c | 165.27 ± 7.23 d | 159.38 ± 13.80 d | 179.79 ± 3.88 d | 193.57 ± 7.68 d | |
| Glutathione (mmol kg−1) | Whole | 0.23 ± 0.004 a | 0.20 ± 0.015 c | 0.25 ± 0.011 d | 0.27 ± 0.007 d | 0.24 ± 0.012 d | 0.25 ± 0.015 d | 0.26 ± 0.015 d |
| Piece | 0.20 ± 0.006 c | 0.15 ± 0.007 d | 0.29 ± 0.005 c | 0.52 ± 0.015 c | 0.52 ± 0.006 c | 0.35 ± 0.007 c | 0.43 ± 0.014 c | |
| Strip | 0.22 ± 0.004 b | 0.24 ± 0.011 b | 0.41 ± 0.004 b | 0.68 ± 0.002 b | 0.75 ± 0.016 b | 0.58 ± 0.009 b | 0.58 ± 0.005 b | |
| Slice | 0.23 ± 0.017 ab | 0.33 ± 0.003 a | 0.54 ± 0.004 a | 0.88 ± 0.006 a | 0.80 ± 0.016 a | 0.77 ± 0.006 a | 0.87 ± 0.014 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.; Guan, Y.; Wang, Y.; Yuan, N. Effect of Wounding Intensity on Edible Quality by Regulating Physiological and ROS Metabolism in Fresh-Cut Pumpkins. Horticulturae 2023, 9, 512. https://doi.org/10.3390/horticulturae9040512
Hu W, Guan Y, Wang Y, Yuan N. Effect of Wounding Intensity on Edible Quality by Regulating Physiological and ROS Metabolism in Fresh-Cut Pumpkins. Horticulturae. 2023; 9(4):512. https://doi.org/10.3390/horticulturae9040512
Chicago/Turabian StyleHu, Wenzhong, Yuge Guan, Yi Wang, and Ning Yuan. 2023. "Effect of Wounding Intensity on Edible Quality by Regulating Physiological and ROS Metabolism in Fresh-Cut Pumpkins" Horticulturae 9, no. 4: 512. https://doi.org/10.3390/horticulturae9040512
APA StyleHu, W., Guan, Y., Wang, Y., & Yuan, N. (2023). Effect of Wounding Intensity on Edible Quality by Regulating Physiological and ROS Metabolism in Fresh-Cut Pumpkins. Horticulturae, 9(4), 512. https://doi.org/10.3390/horticulturae9040512