


Potential Influence of Shading in Freshwater Ponds on the Water Quality Parameters and the Hematological and Biochemical Profiles of Nile tilapia (Oreochromis niloticus Linnaeus, 1758)

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
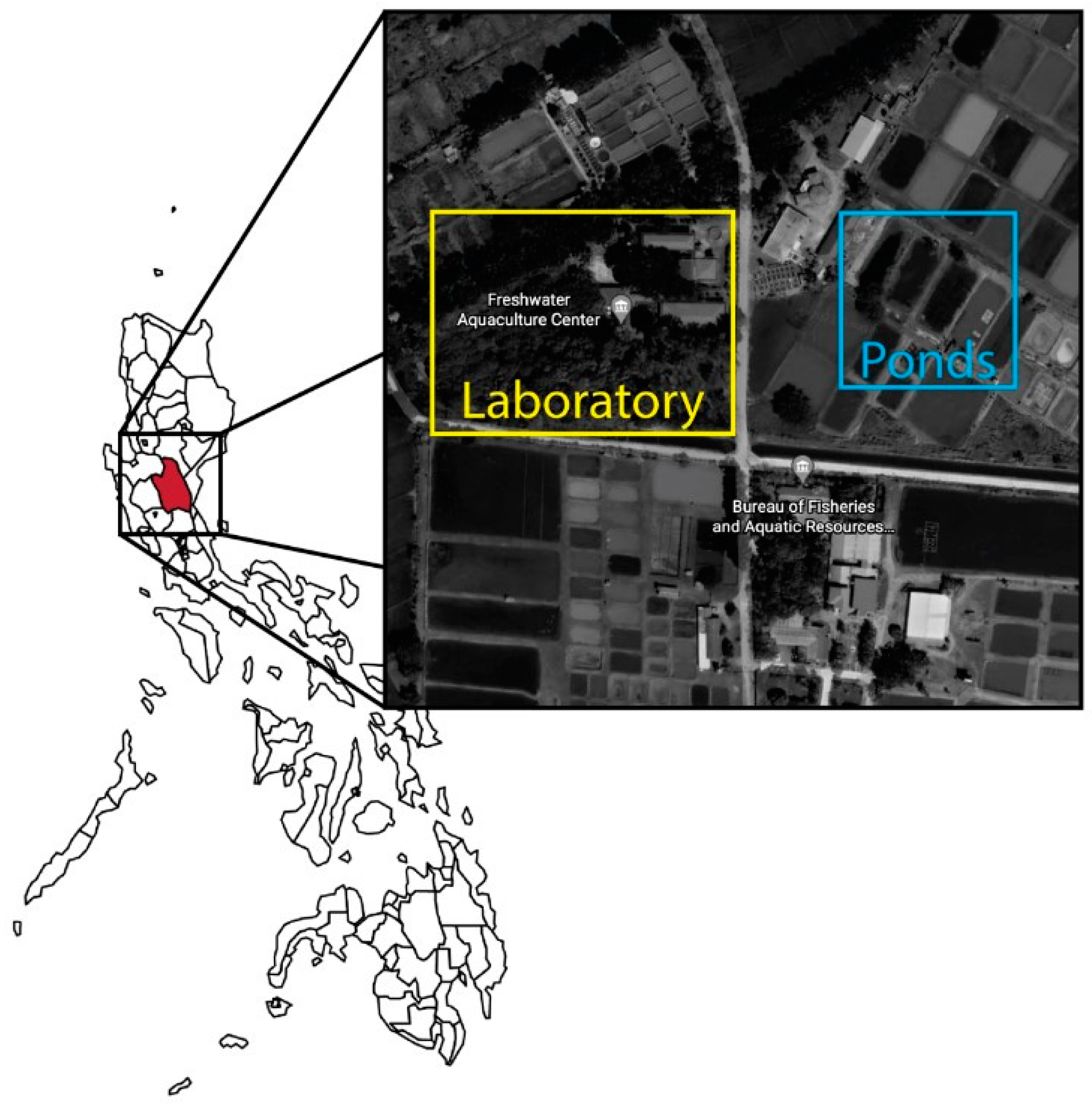
2.1. Area of the Study
2.2. Experimental Setup
2.3. Water Quality Analysis
2.4. Fish Biometric Index
2.5. Hematological Study
2.6. Biochemical Study
2.7. Statistical Analyses
3. Results
3.1. Water Quality
3.2. Biometric Index
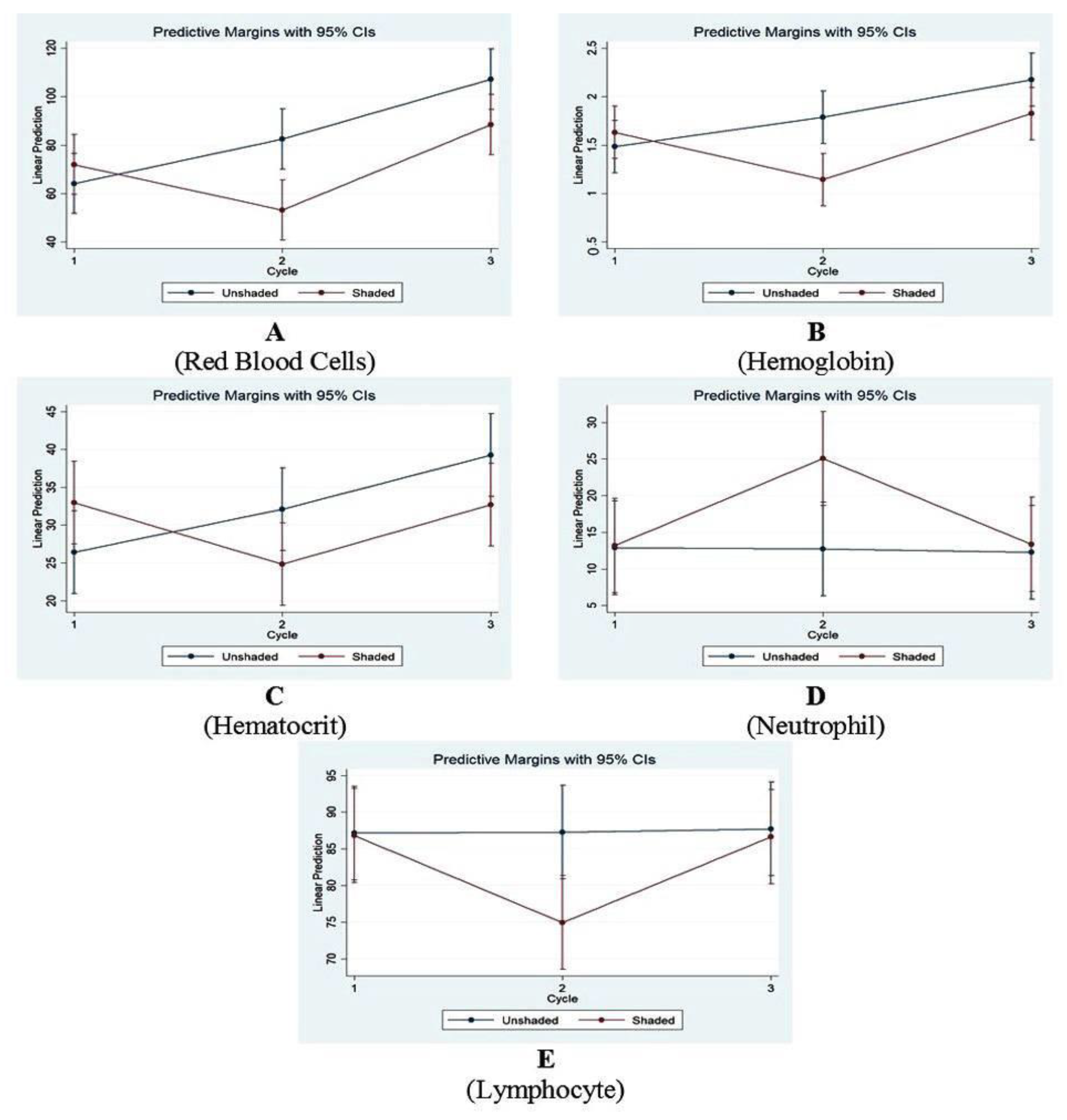
3.3. Hematological Study
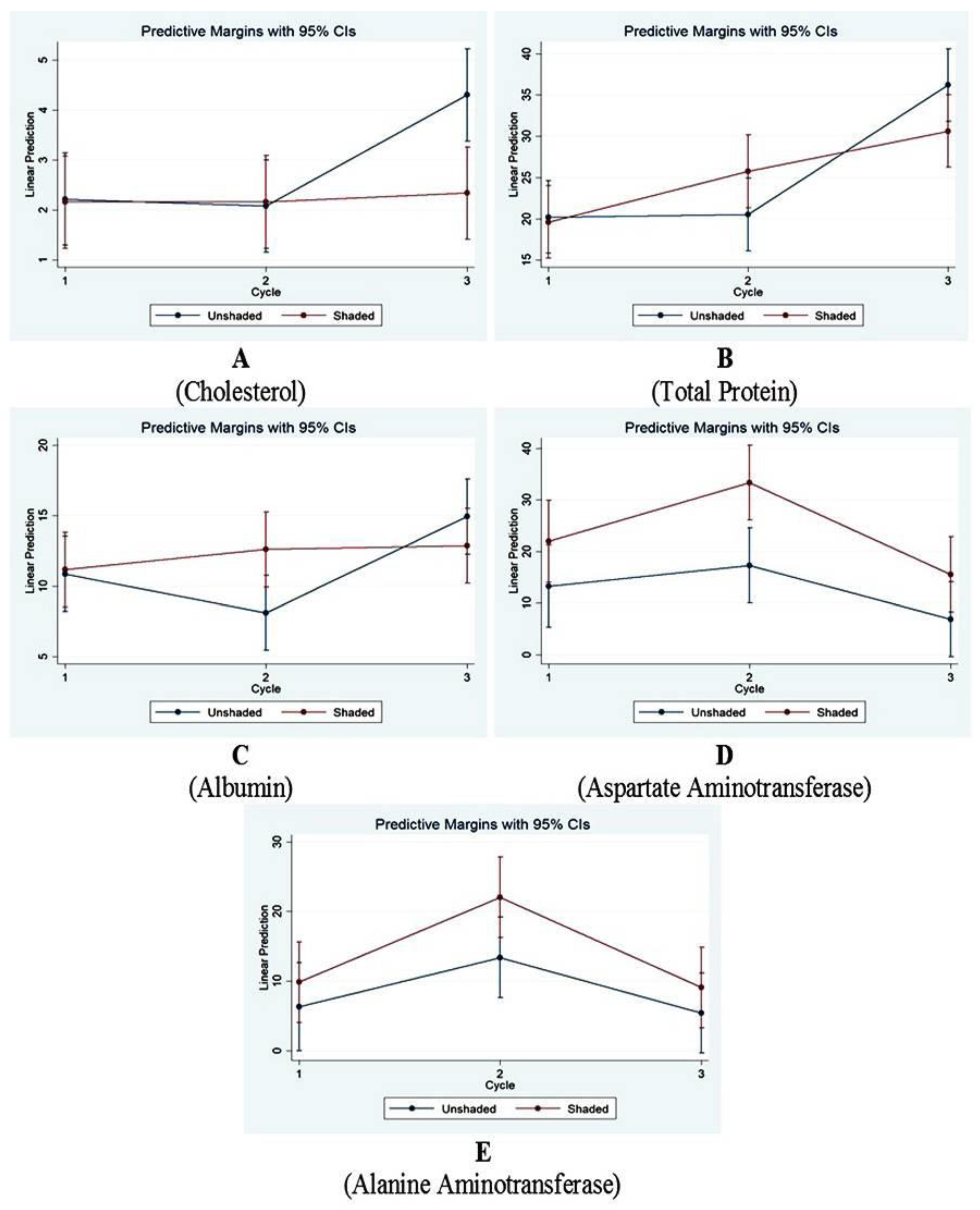
3.4. Biochemical Study
3.5. Canonical Correlation Analysis (CCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Independent Evaluation Overview of Freshwater Aquaculture of Tilapia in the Philippines 2004. Asian Development Bank, July 2004; pp. 48–70. Available online: https://www.adb.org/documents/overview-freshwater-aquaculture-tilapia-philippines (accessed on 6 March 2023).
- Nile Tilapia “Super Strains” to Help Philippines. Available online: https://worldfishcenter.org/press-release/nile-tilapia-super-strains-help-philippines (accessed on 6 March 2023).
- Guerrero, R.D.I. Farmed Tilapia Production in the Philippines Is Declining: What Has Happened and What Can Be Done. Philipp. J. Sci. 2019, 148, 9–15. [Google Scholar]
- Ordoñez, J.F.F.; Santos, M.D.; Tayamen, M.M. Tilapia Genetic R&D in the Philippines: Challenges and Prospects for Future Development. Fish People 2014, 12, 30–43. [Google Scholar]
- Fitzsimmons, K.M. Global Developments and Market Trends in Tilapia for 2018. In Proceedings of the Asian Aquaculture Conference, Bangkok, Thailand, 3–6 December 2018. [Google Scholar]
- Lam, V.W.Y.; Cheung, W.W.L.; Reygondeau, G.; Sumaila, U.R. Projected Change in Global Fisheries Revenues under Climate Change. Sci. Rep. 2016, 6, 32607. [Google Scholar] [CrossRef] [Green Version]
- Fatma, S.; Ahmed, I. Effect of Water Temperature on Protein Requirement of Heteropneustes fossilis (Bloch) Fry as Determined by Nutrient Deposition, Hemato-Biochemical Parameters and Stress Resistance Response. Fish. Aquat. Sci. 2020, 23, 1. [Google Scholar] [CrossRef] [Green Version]
- Bhujel, R.C.; Turner, W.A.; Yakupitiyage, A.; Little, D.C. Impacts of Environmental Manipulation on the Reproductive Performance of Nile Tilapia (Oreochromis niloticus). J. Aquac. Trop. 2001, 16, 197–209. [Google Scholar]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Nader, M.M.; Salem, H.M.; El-Tahan, A.M.; Soliman, S.M.; Khafaga, A.F. Effect of Environmental Factors on Growth Performance of Nile Tilapia (Oreochromis niloticus). Int. J. Biometeorol. 2022, 66, 2183–2194. [Google Scholar] [CrossRef]
- Chan, C.Y.; Tran, N.; Dao, D.C.; Sulser, T.B.; Phillips, M.J.; Batka, M.; Wiebe, K.; Preston, N. Fish to 2050 in the ASEAN Region. Available online: https://globalfutures.cgiar.org/2017/02/10/fish-to-2050-in-the-asean-region/#:~:text=Both%20aquaculture%20and%20capture%20fisheries,exports%20will%20continue%20to%20increase (accessed on 6 March 2023).
- Mapfumo, B. Tilapia Trade Global and Regional Trends. Available online: https://www.fao.org/fi/static-media/MeetingDocuments/TiLV/dec2018/p13.pdf (accessed on 6 March 2023).
- Tang, U.M.; Muchlisin, Z.A.; Syawal, H.; Masjudi, H. Effect of Water Temperature on the Physiological Stress and Growth Performance of Tapah (Wallago leeri) during Domestication. Fish. Aquat. Life 2017, 25, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Musa, N.; Ramly, H.R.; Manaf, M.T.A.; Razzak, L.A.; Musa, N. High Water Temperature Impairs Physiological Responses in Red Hybrid Tilapia: Effects on Cortisol and Its Regulation. Aquac. Aquar. Conserv. Legis. 2017, 10, 1297–1308. [Google Scholar]
- O’Gorman, E.J.; Ólafsson, Ó.P.; Demars, B.O.L.; Friberg, N.; Guðbergsson, G.; Hannesdóttir, E.R.; Jackson, M.C.; Johansson, L.S.; McLaughlin, Ó.B.; Ólafsson, J.S.; et al. Temperature Effects on Fish Production across a Natural Thermal Gradient. Glob. Chang. Biol. 2016, 22, 3206–3220. [Google Scholar] [CrossRef] [Green Version]
- Siddique, M.A.B.; Ahammad, A.K.S.; Bashar, A.; Hasan, N.A.; Mahalder, B.; Alam, M.M.; Biswas, J.C.; Haque, M.M. Impacts of Climate Change on Fish Hatchery Productivity in Bangladesh: A Critical Review. Heliyon 2022, 8, e11951. [Google Scholar] [CrossRef]
- Fazio, F. Fish Hematology Analysis as an Important Tool of Aquaculture: A Review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Whitney, J.E.; Al-Chokhachy, R.; Bunnell, D.B.; Caldwell, C.A.; Cooke, S.J.; Eliason, E.J.; Rogers, M.; Lynch, A.J.; Paukert, C.P. Physiological Basis of Climate Change Impacts on North American Inland Fishes. Fisheries 2016, 41, 332–345. [Google Scholar] [CrossRef]
- Harvey, R.; Lye, L.; Khan, A.; Paterson, R. The Influence of Air Temperature on Water Temperature and the Concentration of Dissolved Oxygen in Newfoundland Rivers. Can. Water Resour. J. 2013, 36, 171–192. [Google Scholar] [CrossRef]
- Wang, B.; Mao, H.; Zhao, J.; Liu, Y.; Wang, Y.; Du, X. Influences of Oxygen and Temperature Interaction on the Antibacterial Activity, Antioxidant Activity, Serum Biochemical Indices, Blood Indices and Growth Performance of Crucian Carp. PeerJ 2023, 11, e14530. [Google Scholar] [CrossRef]
- Jiang, X.; Dong, S.; Liu, R.; Huang, M.; Dong, K.; Ge, J.; Gao, Q.; Zhou, Y. Effects of Temperature, Dissolved Oxygen, and Their Interaction on the Growth Performance and Condition of Rainbow Trout (Oncorhynchus mykiss). J. Therm. Biol. 2021, 98, 102928. [Google Scholar] [CrossRef]
- Imsland, A.K.; Foss, A.; Gunnarsson, S.; Berntssen, M.H.G.; FitzGerald, R.; Bonga, S.W.; Ham, E.v.; Nævdal, G.; Stefansson, S.O. The Interaction of Temperature and Salinity on Growth and Food Conversion in Juvenile Turbot (Scophthalmus maximus). Aquaculture 2001, 198, 353–367. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Bock, C.; Mark, F.C. Oxygen- and Capacity-Limited Thermal Tolerance: Bridging Ecology and Physiology. J. Exp. Biol. 2017, 220, 2685–2696. [Google Scholar] [CrossRef] [Green Version]
- Sopinka, N.M.; Donaldson, M.R.; O’Connor, C.M.; Suski, C.D.; Cooke, S.J. Stress Indicators in Fish. Fish Physiol. 2016, 35, 405–462. [Google Scholar] [CrossRef]
- Marcos-López, M.; Gale, P.; Oidtmann, B.C.; Peeler, E.J. Assessing the Impact of Climate Change on Disease Emergence in Freshwater Fish in the United Kingdom. Transbound. Emerg. Dis. 2010, 57, 293–304. [Google Scholar] [CrossRef]
- Hassan, M.A.; Abd Allah, N.A.; Mabrok, M. Inevitable Impact of Some Environmental Stressors on the Frequency and Pathogenicity of Marine Vibriosis. Aquaculture 2021, 536, 736447. [Google Scholar] [CrossRef]
- Cho, H.C.; Kim, J.E.; Kim, H.B.; Baek, H.J. Effects of Water Temperature Change on the Hematological Responses and Plasma Cortisol Levels in Growing of Red Spotted Grouper, Epinephelus akaara. Dev. Reprod. 2015, 19, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Clauss, T.M.; Dove, A.D.M.; Arnold, J.E. Hematologic Disorders of Fish. Vet. Clin. N. Am. Exot. Anim. Pract. 2008, 11, 445–462. [Google Scholar] [CrossRef]
- Fazio, F.; Ferrantelli, V.; Piccione, G.; Saoca, C.; Levanti, M.; Mucciardi, M. Biochemical and Hematological Parameters in European Sea Bass (Dicentrarchus labrax Linnaeus, 1758) and Gilthead Sea Bream (Sparus aurata Linnaeus, 1758) in Relation to Temperature. Vet. Arh. 2018, 88, 397–411. [Google Scholar] [CrossRef]
- Harvey, J.W. Chapter 2—Hematology Procedures. In Veterinary Hematology; Harvey, J.W., Ed.; W.B. Saunders: Saint Louis, MO, USA, 2012; pp. 11–32. ISBN 978-1-4377-0173-9. [Google Scholar]
- Gracey, A.Y.; Fraser, E.J.; Li, W.; Fang, Y.; Taylor, R.R.; Rogers, J.; Brass, A.; Cossins, A.R. Coping with Cold: An Integrative, Multitissue Analysis of the Transcriptome of a Poikilothermic Vertebrate. Proc. Natl. Acad. Sci. USA 2004, 101, 16970–16975. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.R.; Johnston, I.A. Temperature during Embryonic Development Has Persistent Effects on Thermal Acclimation Capacity in Zebrafish. Proc. Natl. Acad. Sci. USA 2012, 109, 14247–14252. [Google Scholar] [CrossRef] [Green Version]
- Bowden, T.J.; Thompson, K.D.; Morgan, A.L.; Gratacap, R.M.L.; Nikoskelainen, S. Seasonal Variation and the Immune Response: A Fish Perspective. Fish Shellfish Immunol. 2007, 22, 695–706. [Google Scholar] [CrossRef]
- Makori, A.J.; Abuom, P.O.; Kapiyo, R.; Anyona, D.N.; Dida, G.O. Effects of Water Physico-Chemical Parameters on Tilapia (Oreochromis niloticus) Growth in Earthen Ponds in Teso North Sub-County, Busia County. Fish. Aquat. Sci. 2017, 20, 30. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, P.; Andersen, K.H. Thermal Performance of Fish Is Explained by an Interplay between Physiology, Behaviour and Ecology. Conserv. Physiol. 2019, 7, coz025. [Google Scholar] [CrossRef] [Green Version]
- Pullin, R.S.V.; McConnell, R. The Biology and Culture of Tilapias; Conference Proceedings; ICLARM: Makati, Philippines, 1982; ISBN 978-971-0400-04-1. [Google Scholar]
- Ling, T.-Y.; Soo, C.-L.; Phan, T.-P.; Lee, N.; Sim, S.-F.; Grinang, J. Assessment of the Water Quality of Batang Rajang at Pelagus Area, Sarawak, Malaysia. Sains Malays. 2017, 46, 401–411. [Google Scholar] [CrossRef]
- Memiş, D.; Ercan, E.; Yamaner, G. The Effects of Shaded Pond on Growth Performance of Russian Sturgeon (Acipenser gueldenstaedtii). J. Appl. Ichthyol. 2011, 27, 571–575. [Google Scholar] [CrossRef]
- Schulte, P.M.; Davies, S.A.; Dow, J.A.T.; Lukowiak, K. What Is Environmental Stress? Insights from Fish Living in a Variable Environment. J. Exp. Biol. 2014, 217, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Chong, S.; Park, C.; Lee, K.R.; An, K.-G. Modeling Summer Hypoxia Spatial Distribution and Fish Habitat Volume in Artificial Estuarine Waterway. Water 2018, 10, 1695. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, H.V.; Acre, M.R.; Faulkner, J.D.; da Cunha, L.R.; Lawson, K.M.; Wamboldt, J.J.; Brey, M.K.; Woodley, C.M.; Calfee, R.D. Effects of Shady Environments on Fish Collective Behavior. Sci. Rep. 2022, 12, 17873. [Google Scholar] [CrossRef]
- Vivanco-Aranda, M.; Tanahara-Romero, S.A.; Lugo-Ibarra, K.D.C.; Rio-Zaragoza, O.B.D. Hematological and Serum Chemistry Profile in Wild Pacific Mackerel (Scomber japonicus) from Todos Santos Bay, Baja California, Mexico. Lat. Am. J. Aquat. Res. 2022, 50, 50–59. [Google Scholar] [CrossRef]
- Kohanestani, Z.M.; Hajimoradloo, A.; Ghorbani, R.; Yulghi, S.; Hoseini, A.; Molaee, M. Seasonal Variations in Hematological Parameters of Alburnoides eichwaldii in Zaringol Stream-Golestan Province, Iran. World J. Fish Mar. Sci. 2013, 5, 121–126. [Google Scholar]
- Panase, P.; Saenphet, S.; Saenphet, K. Biochemical and Physiological Responses of Nile Tilapia Oreochromis niloticus Lin Subjected to Cold Shock of Water Temperature. Aquac. Rep. 2018, 11, 17–23. [Google Scholar] [CrossRef]
- Assem, H.; Hassan, B.; Khalifa, A.; El Salhia, M.; Al Basomy, A.; El Sayed, M. The Combined Effect of Environmental Thermal Drop and Isotonicity on Metabolic Stores of the Teleost, Oreochromis niloticus. Egypt. J. Aquat. Res. 2013, 39, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Del Rio-Zaragoza, O.B.; Fajer-Avila, E.J.; Almazán-Rueda, P. Haematological and Gill Responses to an Experimental Infection of Dactylogyrid Monogeneans on the Spotted Rose Snapper Lutjanus guttatus (Steindachner, 1869). Aquac. Res. 2010, 41, 1592–1601. [Google Scholar] [CrossRef]
- Rodnick, K.J.; Planas, J.V. The Stress and Stress Mitigation Effects of Exercise: Cardiovascular, Metabolic, and Skeletal Muscle Adjustments. Fish Physiol. 2016, 35, 251–294. [Google Scholar] [CrossRef]
- Ndong, D.; Chen, Y.-Y.; Lin, Y.H.; Vaseeharan, B.; Chen, J.C. The Immune Response of Tilapia Oreochromis mossambicus and Its Susceptibility to Streptococcus iniae under Stress in Low and High Temperatures. Fish Shellfish Immunol. 2007, 22, 686–694. [Google Scholar] [CrossRef]
- Weyts, F.A.A.; Cohen, N.; Flik, G.; Verburg-van Kemenade, B.M.L. Interactions between the Immune System and the Hypothalamo-Pituitary-Interrenal Axis in Fish. Fish Shellfish Immunol. 1999, 9, 1–20. [Google Scholar] [CrossRef]
- Macnab, V.; Barber, I. Some (Worms) like It Hot: Fish Parasites Grow Faster in Warmer Water, and Alter Host Thermal Preferences. Glob. Chang. Biol. 2012, 18, 1540–1548. [Google Scholar] [CrossRef]
- De, M.; Abd. Ghaffar, M.; Md. Noor, N.; Cob, Z.C.; Bakar, Y.; Das, S.K. Effects of Water Temperature and Diet on Blood Parameters and Stress Levels in Hybrid Grouper (Epinephelus fuscoguttatus ♀ × E. Lanceolatus ♂) Juveniles. Aquac. Rep. 2019, 15, 100219. [Google Scholar] [CrossRef]
- EL-Khaldi, A.T.F. Effect of Different Stress Factors on Some Physiological Parameters of Nile Tilapia (Oreochromis niloticus). Saudi J. Biol. Sci. 2010, 17, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Volpato, G.L.; Barreto, R.E. Environmental Blue Light Prevents Stress in the Fish Nile Tilapia. Braz. J. Med. Biol. Res. 2001, 34, 1041–1045. [Google Scholar] [CrossRef] [Green Version]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Hepatic Glucokinase and Glucose-6-Phosphatase Responses to Dietary Glucose and Starch in Gilthead Sea Bream (Sparus aurata) Juveniles Reared at Two Temperatures. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 149, 80–86. [Google Scholar] [CrossRef]
- Couto, A.; Enes, P.; Peres, H.; Oliva-Teles, A. Effect of Water Temperature and Dietary Starch on Growth and Metabolic Utilization of Diets in Gilthead Sea Bream (Sparus aurata) Juveniles. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 45–50. [Google Scholar] [CrossRef]
- Hemre, G.-I.; Mommsen, T.P.; Krogdahl, Å. Carbohydrates in Fish Nutrition: Effects on Growth, Glucose Metabolism and Hepatic Enzymes. Aquac. Nutr. 2002, 8, 175–194. [Google Scholar] [CrossRef]
- Mauel, M.J.; Miller, D.L.; Merrill, A.L. Hematologic and Plasma Biochemical Values of Healthy Hybrid Tilapia (Oreochromis aureus × Oreochromis nilotica) Maintained in a Recirculating System. J. Zoo Wildl. Med. 2007, 38, 420–424. [Google Scholar] [CrossRef]
- Coz-Rakovac, R.; Strunjak-Perovic1, I.; Hacmanjek, M.; Popovic, N.T.; Lipej, Z.; Sostaric, B. Blood Chemistry and Histological Properties of Wild and Cultured Sea Bass (Dicentrarchus labrax) in the North Adriatic Sea. Vet. Res. Commun. 2005, 29, 677–687. [Google Scholar] [CrossRef]
- Sadauskas-Henrique, H.; Sakuragui, M.M.; Paulino, M.G.; Fernandes, M.N. Using Condition Factor and Blood Variable Biomarkers in Fish to Assess Water Quality. Environ. Monit. Assess. 2011, 181, 29–42. [Google Scholar] [CrossRef]
- Tavares-Dias, M.; Martins, M.L.; Moraes, F.R. Relação hepatosomática e esplenosomática em peixes teleósteos de cultivo intensivo. Rev. Bras. Zool. 2000, 17, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Chacoff, L.; Arjona, F.J.; Ruiz-Jarabo, I.; García-Lopez, A.; Flik, G.; Mancera, J.M. Water Temperature Affects Osmoregulatory Responses in Gilthead Sea Bream (Sparus aurata L.). J. Therm. Biol. 2020, 88, 102526. [Google Scholar] [CrossRef]
- Phinrub, W.; Lunjirapan, T.; Srirum, T.; Kumjumrern, K.; Srisuttha, P.; Panase, A.; Panase, P. Alterations of Serum Electrolytes and Biochemical Indices of Panagasianodon gigas Subjected to Different Water Temperatures and the Appropriate Temperature Range for Sustaining Life. J. Appl. Anim. Res. 2023, 51, 342–349. [Google Scholar] [CrossRef]
- Fiess, J.C.; Kunkel-Patterson, A.; Mathias, L.; Riley, L.G.; Yancey, P.H.; Hirano, T.; Grau, E.G. Effects of Environmental Salinity and Temperature on Osmoregulatory Ability, Organic Osmolytes, and Plasma Hormone Profiles in the Mozambique Tilapia (Oreochromis mossambicus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 252–264. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Parameters | Environmental Condition | |
|---|---|---|
| Unshaded | Shaded | |
| Water temperature (°C) | 32.89 | 30.73 |
| Dissolved oxygen (mg/L) | 4.26 | 4.99 |
| Sunshine duration (min) | 550 | |
| Water Quality | Cycle 1 | Cycle 2 | Cycle 3 | |||
|---|---|---|---|---|---|---|
| Setups | Unshaded | Shaded | Unshaded | Shaded | Unshaded | Shaded |
| Water Temperature (°C) | 30.3 ± 0.55 a | 29.0 ± 0.33 b | 28.9 ± 0.66 a | 27.85 ± 0.49 b | 29.85 ± 0.60 a | 28.45 ± 0.38 b |
| Dissolved Oxygen (mg/L) | 1.69 ± 0.33 a | 2.45 ± 0.55 b | 1.39 ± 0.038 | 1.38 ± 0.21 | 2.27 ± 0.93 | 2.31 ± 1.11 |
| Water Quality | Unshaded | Shaded |
|---|---|---|
| Water Temperature (°C) | 29.68 ± 0.83 a | 28.43 ± 0.62 b |
| Dissolved Oxygen (mg/L) | 1.78 ± 0.65 a | 2.04 ± 0.81 a |
| S | C | WBCs (109/L) | RBCs (1012/L) | Hgb (g/L) | Hct (%) | Pct (109/L) | MCV (fL) | MCH (pg) | MCHC (g/L) | Neutro (109/L) | Lympho (109/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| No | 1 | 64.08 ± 10.64 a | 1.48 ± 0.43 a | 64.17 ± 17.94 a | 26.43 ± 5.14 a | 7.67 ± 3.61 a | 185.00 ± 28.49 a | 43.50 ± 1.55 a | 238.83 ± 29.14 a | 12.88 ± 4.33 a | 87.13 ± 4.33 a |
| Yes | 1 | 67.92 ± 12.34 a | 1.63 ± 0.25 b | 72 ± 10.02 b | 32.95 ± 7.35 b | 7.67 ± 2.80 a | 200.04 ± 22.04 b | 44.27 ± 2.12 a | 223.00 ± 28.74 a | 13.20 ± 6.07 b | 86.80 ± 6.07 b |
| No | 2 | 70.88 ± 18.75 a | 1.79 ± 0.19 c | 82.50 ± 11.57 c | 32.12 ± 2.84 c | 7.83 ± 2.93 b | 180.95 ± 20.76 a | 46.03 ± 2.89 b | 256.17 ± 22.94 a | 12.72 ± 3.55 c | 87.28 ± 3.55 c |
| Yes | 2 | 54.90 ± 16.69 a | 1.14 ± 0.09 d | 53.17 ± 3.54 d | 24.82 ± 3.43 d | 11.67 ± 5.54 b | 206.08 ± 29.22 b | 46.45 ± 1.34 b | 228.17 ± 36.49 a | 25.05 ± 7.88 d | 74.95 ± 7.88 d |
| No | 3 | 75.58 ± 7.29 a | 2.17 ± 0.18 e | 107.17 ± 11.11 e | 39.27 ± 5.59 e | 4.50 ± 1.38 c | 175.38 ± 12.94 a | 48.18 ± 2.29 c | 235.73 ± 110.98 a | 12.27 ± 2.22 e | 87.73 ± 2.22 e |
| Yes | 3 | 64.55 ± 8.13 a | 1.83 ± 0.08 f | 88.50 ± 1.87 f | 32.72 ± 1.68 f | 7.33 ± 4.03 c | 179.02 ± 3.62 b | 48.53 ± 1.75 c | 271.50 ± 13.31 a | 13.37 ± 7.07 f | 86.63 ± 7.07 f |
| Parameters | Pond Setup | |
|---|---|---|
| Unshaded | Shaded | |
| WBCs (109/L) | 70.18 ± 13.26 | 62.46 ± 13.36 |
| RBCs (1012/L) | 1.82 ± 0.40 a | 1.53 ± 0.33 b |
| Hgb (g/L) | 84.61 ± 22.34 a | 71.22 ± 15.97 b |
| Hct (%) | 32.61 ± 6.97 | 30.16 ± 5.94 |
| Pct (109/L) | 6.67 ± 2.64 | 8.89 ± 4.12 |
| MCV (fL) | 180.44 ± 20.76 b | 195.29 ± 23.30 a |
| MCH (pg) | 45.91 ± 2.93 | 46.42 ± 2.44 |
| MCHC (g/L) | 243.58 ± 64.13 | 240.89 ± 34.46 |
| Neutrophil (109/L) | 12.62 ± 3.28 b | 17.21 ± 8.74 a |
| Lymphocyte (109/L) | 87.38 ± 3.28 a | 82.79 ± 8.74 b |
| S | C | Cor (ng/mL) | Glu (mmol/L) | Chole (mmol/L) | TP (g/L) | Alb (g/L) | Glb (U/L) | AST (U/L) | ALT (U/L) | ALP (U/L) | K+ (mmol/L) | Na+ (mmol/L) | Cl− (mmol/L) | Mg2+ (mmol/L) | TCa (mmol/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No | 1 | 299.24 ± 4.4 a | 4.19 ± 1.15 a | 2.22 ± 0.58 a | 20.23 ± 3.44 a | 10.87 ± 1.79 a | 8.49 ± 3.40 a | 13.28 ± 1.45 aa | 6.33 ± 2.63 aa | 2.41 ± 16.98 a | 3.77 ± 0.62 a | 165.28 ± 8.09 a | 157.93 ± 7.42 a | 2.93 ± 0.58 a | 3.98 ± 0.70 a |
| Yes | 1 | 232.89 ± 49.60 a | 5.27 ± 2.23 a | 2.16 ± 0.71 b | 19.63 ± 2.22 b | 11.17 ± 1.76 b | 9.06 ± 1.50 a | 21.99 ± 7.68 ba | 9.88 ± 1.07 ba | 56.54 ± 11.53 a | 3.27 ± 1.26 a | 160.17 ± 7.20 a | 150.85 ± 7.53 a | 3.33 ± 0.55 a | 3.63 ± 0.48 a |
| No | 2 | 177.03 ± 95.00 b | 4.24 ± 1.09 a | 2.08 ± 0.93 c | 20.54 ± 3.36 c | 8.12 ± 3.20 c | 10.90 ± 4.39 b | 17.30 ± 6.84 ab | 13.39 ± 4.13 ab | 54.57 ± 11.08 a | 4.05 ± 0.59 b | 156.25 ± 5.61 a | 144.43 ± 7.22 b | 3.10 ± 1.06 b | 2.84 ± 0.40 b |
| Yes | 2 | 176.78 ± 78.94 b | 3.84 ± 0.53 a | 2.16 ± 1.07 d | 25.76 ± 4.48 d | 12.60 ± 2.30 d | 13.14 ± 2.61 b | 33.33 ± 9.20 bb | 22.06 ± 10.24 b | 58.44 ± 10.14 a | 4.06 ± 0.99 b | 157.28 ± 3.64 a | 145.47 ± 6.54 b | 3.90 ± 1.73 b | 2.79 ± 0.47 b |
| No | 3 | 245.17 ± 77.86 c | 4.09 ± 0.83 a | 4.31 ± 0.94 e | 36.20 ± 4.12 e | 14.93 ± 1.66 e | 21.28 ± 2.82 c | 6.88 ± 1.68 ac | 5.43 ± 2.24 ac | 67.39 ± 11.78 a | 2.91 ± 0.63 c | 163.88 ± 5.63 a | 148.85 ± 4.34 c | 5.42 ± 1.72 c | 4.50 ± 1.29 c |
| Yes | 3 | 308.60 ± 157.51 c | 4.02 ± 0.76 a | 2.34 ± 0.34 f | 30.63 ± 4.74 f | 12.87 ± 2.70 f | 17.76 ± 2.19 c | 15.56 ± 6.15 bc | 9.09 ± 3.31 bc | 63.82 ± 7.35 a | 2.59 ± 0.99 c | 157.25 ± 5.17 a | 145.03 ± 4.13 c | 5.70 ± 2.53 c | 4.28 ± 1.42 c |
| Parameters | Pond Setup | |
|---|---|---|
| Unshaded | Shaded | |
| Cor (ng/mL) | 244.21 ± 84.07 | 239.42 ± 113.76 |
| Glu (mmol/L) | 4.17 ± 0.97 | 4.38 ± 1.47 |
| Chole (mmol/L) | 2.87 ± 1.31 | 2.22 ± 0.73 |
| TP (g/L) | 25.65 ± 8.41 | 25.34 ± 5.95 |
| Alb (g/L) | 11.31 ± 3.61 | 12.21 ± 2.28 |
| Glb (g/L) | 13.56 ± 6.63 | 13.32 ± 4.18 |
| AST (U/L) | 12.44 ± 6.06 a | 23.73 ± 10.66 b |
| ALT (U/L) | 8.50 ± 4.75 a | 13.68 ± 8.47 b |
| ALP (U/L) | 61.46 ± 13.83 | 59.60 ± 9.76 |
| K+ (mmol/L) | 3.58 ± 0.77 | 3.31 ± 1.19 |
| Na+ (mmol/L) | 161.81 ± 7.38 | 158.23 ± 5.38 |
| Cl− (mmol/L) | 150.41 ± 8.40 | 147.12 ± 6.46 |
| Mg2+ (mg/dL) | 3.82 ± 1.63 | 4.31 ± 1.98 |
| TCa (mmol/L) | 3.77 ± 1.09 | 3.57 ± 1.06 |
| DO | WT | |||
|---|---|---|---|---|
| Morning | Afternoon | Morning | Afternoon | |
| AM_DO | 1.0000 | |||
| PM_DO | 0.9299 * | 1.0000 | ||
| AM_WT | 0.4539 | 0.7402 * | 1.0000 | |
| PM_WT | 0.6124 * | 0.8551 * | 0.9706 * | 1.0000 |
| WBCs | RBCs | HGB | HCT | |
|---|---|---|---|---|
| WBCs | 1.0000 | |||
| RBCs | 0.3710 | 1.0000 | ||
| Hgb | 0.2931 | 0.9648 * | 1.0000 | |
| Hct | 0.5496 * | 0.8459 * | 0.7392 * | 1.0000 |
| Cortisol | Glu | K | Na | Alb | Alt | |
|---|---|---|---|---|---|---|
| Cor | 1.0000 | |||||
| Glu | −0.1447 | 1.0000 | ||||
| K+ | 0.1147 | 0.1789 | 1.0000 | |||
| Na+ | −0.3888 | 0.8811 * | 0.0536 | 1.0000 | ||
| Alb | −0.6378 * | 0.2638 | −0.2220 | 0.5551 * | 1.0000 | |
| ALT | −0.2072 | −0.1759 | 0.5361 * | −0.0374 | 0.0417 | 1.0000 |
| CC1 | CC2 | CC3 | CC4 | |
|---|---|---|---|---|
| Canonical correlation coefficient | 0.9639 ** | 0.9000 ** | 0.8578 ** | 0.3165 |
| Standardized canonical coefficients for the hematological parameters | ||||
| WBCs | 0.6572 * | −0.0123 | −0.7105 | 1.2837 |
| RBCs | −0.5346 | 3.8764 * | 1.0083 | 2.0095 |
| Hgb | 1.3193 * | −3.6007 * | 0.0155 | −1.2506 |
| Hct | −0.2418 | −0.0547 | −0.2952 | −2.1678 |
| Standardized canonical coefficients for the water quality parameters | ||||
| AM_DO | −7.7032 * | 5.4862 * | 2.4225 | −1.8528 |
| PM_DO | 10.1404 * | −8.1625 * | −5.9993 * | 3.3276 |
| AM_WT | −1.8084 * | 1.7332 | −4.8970 * | −2.8808 |
| PM_WT | −1.7801 * | 2.7274 * | 8.2713 * | 1.5154 |
| CC1 | CC2 | CC3 | CC4 | |
|---|---|---|---|---|
| Canonical Correlation Coefficient | 0.9698 ** | 0.9675 ** | 0.8070 ** | 0.6085 |
| Standardized canonical coefficients for the biochemical parameters | ||||
| Cor | 0.6819 * | −0.7661 * | 0.6282 | 0.5375 |
| Glu | 0.5817 * | 0.7888 * | 0.1783 | 1.1354 |
| K+ | 0.3396 * | 0.5162 * | 0.2286 | −0.6224 |
| Na+ | −0.3353 | −0.3648 | −0.5744 | −0.3751 |
| Alb | −0.1065 | −0.1469 | 1.2720 * | 0.5154 |
| ALT | 0.1710 | −0.0067 | 0.5244 | 0.1719 |
| Standardized canonical coefficients for the water quality parameters | ||||
| AM_DO | 2.5705 * | 3.0767 * | −2.3101 | −8.7937 * |
| PM_DO | −5.8858 * | −3.3630 * | 2.0054 | 12.9052 * |
| AM_WT | −3.9112 * | −1.3316 * | −3.8975 * | −2.5087 |
| PM_WT | 7.7767 * | 2.3156 * | 2.7127 | −2.8495 |
| CC1 | CC2 | CC3 | CC4 | CC5 | |
|---|---|---|---|---|---|
| Canonical correlation coefficient | 0.9613 ** | 0.9027 ** | 0.7551 | 0.6140 | 0.2428 |
| Standardized canonical coefficients for the biochemical parameters | |||||
| Cor | 0.6467 * | −0.9950 * | 0.3943 | 0.5198 | 0.1168 |
| Glu | −0.4169 | −0.0598 | −1.1192 | 0.7670 | −0.5938 |
| K+ | 0.5407 * | 0.4905 * | 0.8814 * | −0.6632 | −0.5638 |
| Na | 0.4636 | −0.2759 | 1.0263 | −1.2460 | 1.6634 |
| Alb | −0.1743 | −0.2345 | 0.8952 | 0.9808 | −0.6221 |
| ALT | 0.2925 * | 0.1291 | −0.6186 | 0.9984 | 0.6875 |
| Standardized canonical coefficients for the hematological parameters | |||||
| WBCs | −0.5574 * | 0.2547 | 0.1556 | 0.1665 | −2.0119 |
| RBCs | 2.6496 * | −1.3751 | 0.5989 | −5.7538 * | 1.2531 |
| Hgb | −1.5591 * | 0.0257 | 0.7708 | 4.5073 * | −0.6533 |
| Hct | −0.3965 | 0.1456 | −1.7538 * | 1.5066 | 0.7254 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dayrit, G.B.; Vera Cruz, E.M.; Rodkhum, C.; Mabrok, M.; Ponza, P.; Santos, M.D. Potential Influence of Shading in Freshwater Ponds on the Water Quality Parameters and the Hematological and Biochemical Profiles of Nile tilapia (Oreochromis niloticus Linnaeus, 1758). Fishes 2023, 8, 322. https://doi.org/10.3390/fishes8060322
Dayrit GB, Vera Cruz EM, Rodkhum C, Mabrok M, Ponza P, Santos MD. Potential Influence of Shading in Freshwater Ponds on the Water Quality Parameters and the Hematological and Biochemical Profiles of Nile tilapia (Oreochromis niloticus Linnaeus, 1758). Fishes. 2023; 8(6):322. https://doi.org/10.3390/fishes8060322
Chicago/Turabian StyleDayrit, Geraldine B., Emmanuel M. Vera Cruz, Channarong Rodkhum, Mahmoud Mabrok, Pattareeya Ponza, and Mudjekeewis D. Santos. 2023. "Potential Influence of Shading in Freshwater Ponds on the Water Quality Parameters and the Hematological and Biochemical Profiles of Nile tilapia (Oreochromis niloticus Linnaeus, 1758)" Fishes 8, no. 6: 322. https://doi.org/10.3390/fishes8060322
APA StyleDayrit, G. B., Vera Cruz, E. M., Rodkhum, C., Mabrok, M., Ponza, P., & Santos, M. D. (2023). Potential Influence of Shading in Freshwater Ponds on the Water Quality Parameters and the Hematological and Biochemical Profiles of Nile tilapia (Oreochromis niloticus Linnaeus, 1758). Fishes, 8(6), 322. https://doi.org/10.3390/fishes8060322