1. Introduction
Golden pompano (
Trachinotus ovatus) is a valuable edible fish with thornless flesh, a tender texture, and a delicious taste, with the unique aroma of the trevally family. It is distributed in the Atlantic, Indian, and Pacific Oceans’ tropical and subtropical seas. In China, the coasts of Guangdong, Guangxi, Hainan, and Fujian are main growing regions [
1]. The demand for protein raw materials (e.g., fishmeal or soybean meal) is rising as aquaculture production increases. Import policies have a significant impact on the aquaculture industry, since fishmeal and soybean meal are import-dependent. To fulfill the rising domestic demand for protein raw materials, guarantee a continuous supply of aquatic products, and maintain global food security, it is crucial to discover and develop novel, effective, sustainable, and environmentally friendly protein sources.
The ability of the intestines to digest and absorb nutrients is crucial for fish, especially stomachless fish, and is considered to be an important indicator of the overall health of the body [
2]. There are large amounts of microflora in the intestines of fish which live in symbiosis with the fish and are known as the normal intestinal flora. Fish with normal intestinal flora can have better digestion and receive essential nutrients which strengthen their natural defenses and immunity [
3]. Dysbacteriosis has the potential to trigger enteritis, leading to diminished appetite, stunted growth, and potentially fatal outcomes in fish [
4]. An effective and properly controlled intestinal barrier is crucial for safeguarding the organism against food antigens and its indigenous intestinal bacteria [
5]. As a result, research on the structure and changes of fish intestinal microflora has recently gained attention in the aquaculture industry.
Cottonseed meal (CSM) serves as a valuable source of protein of superior quality. The utilization of this nutrient-dense resource as animal feed, however, encounters obstacles due to the presence of free gossypol (FG) [
6]. FG-containing diets can have negative effects on animal growth, digestive health, and reproduction. Fermentation technology allows for efficient separation of cottonseed phenols in cottonseed meals, raising the crude protein level of the raw material and producing small peptides and growth-promoting factors that enrich the nutrition of cottonseed protein [
7]. The application of fermented cottonseed protein in mackerel culture was investigated, and it was found that the survival and weight gain rates of mackerel in the group with fermented cottonseed protein were significantly higher than those in the control group [
8]. It was discovered that using 23% FCSM to replace 9% soybean meal and 15% cottonseed meal promoted the growth of grass carp, reduced the feed coefficient, and improved nonspecific immunity [
9]. Most current research on fermented cottonseed protein is gathered in livestock and poultry, with little focus on aquatic animals. Based on these findings, this study was conducted to investigate the effects of the replacement of fish meal (FM) with FCSM on the intestinal enzymatic activity, inflammatory and physical-barrier-related gene expression, and intestinal microflora of juvenile
T. ovatus.
4. Discussion
Fish meal (FM) is widely utilized as a protein source in commercial aquafeeds due to its well-balanced composition of essential amino acids, vitamins, and minerals, and its palatability [
10]. In recent years, the increasing demand for animal nutrition has led to a search for sustainable protein sources of both plant and animal origin, due to the limited availability of fishmeal [
13]. Cottonseed meal is digested relatively well by fish and crustaceans [
14]. When markets are favorable, CSM is an economical alternative protein source for use in aquatic animal feeds. Furthermore, it is generally highly palatable to most aquatic animals [
14].
The intestinal tracts of animals serve as complex microbial ecosystems that harbor dynamic communities of microorganisms. These microorganisms play various roles, such as absorbing nutrients, improving energy production, and maintaining immune homeostasis [
15]. The process of digestion is related to the activity of digestive enzymes, including chymotrypsin and α-amylase. These enzymes play a crucial role in breaking down nutrients, as they facilitate the digestion process [
16]. In the present study, the intestinal amylase content levels in the CSM12.5 and CSM25 groups were higher than that in the FM group. The chymotrypsin and AMY activity levels in the intestine were higher than in the FM group when the substitution rate reached 25%. AMY activity reflects the absorption and utilization of nutrients by fish [
17]. Protease is an important proteolytic enzyme in fish intestine, and its activity can reflect the ability of fish to decompose feed protein [
18]. These findings indicated that 25% substitution of FCSM for FM boosted the activity of digestive enzymes and improved nutrient uptake, which in turn improved growth performance of
T. ovatus. In this study, when 50–100% of FM was replaced by FCSM, the activity levels of LPS and chymotrypsin were lower than those of the control group. This may be due to the lower digestion and absorption ability of FCSM as compared to fish meal. A similar phenomenon was also observed in a previous study by Sillago sihama Forsskál [
19]. Research findings indicate that the American redfish species (
Sciaenops ocellatus) has demonstrated proficient ability in digesting cottonseed meal [
20].
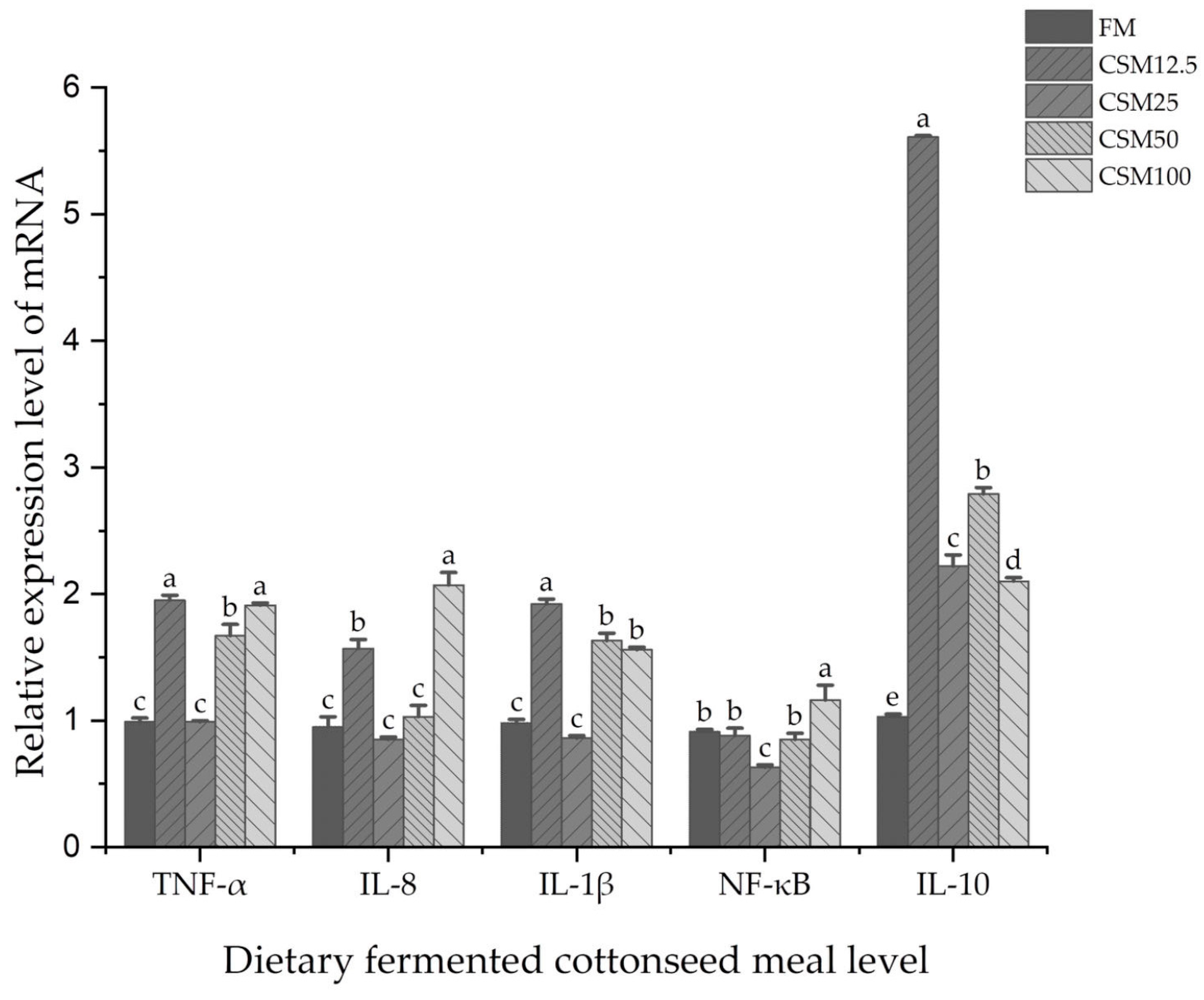
The significant function of pro- and anti-inflammatory cytokines in the maintenance of tissue and immunological homeostasis has been well established [
21]. The pro-inflammatory cytokines, which include
TNF-α and
IL-8, have a variety of effects that cause inflammation [
22]. Interleukin-1β (
IL-1β) plays a crucial role in both the onset and amplification of the inflammatory response, which ultimately leads to intestinal damage [
23]. Inflammatory cytokine
IL-8 is a component of the immune response, and disruption of the balance of these cytokines is key to the pathophysiology of inflammatory bowel disease [
24]. In epithelial tissues,
NF-κB signaling is crucial for preserving immunological homeostasis. Thus,
NF-κB appears to exhibit two sides in chronic inflammation: on the one hand,
NF-κB activation increases and continues to induce inflammation and tissue damage, but on the other hand, inhibition of
NF-κB signaling also disrupts immune homeostasis and triggers inflammation and disease [
22]. In the present study,
TNF-α,
IL-1β, IL-8, and
NF-κB gene expression levels were all significantly decreased at the level of 25% FCSM replacement. In contrast, excessive substitution levels significantly increased
TNF-α,
IL-1β,
IL-8, and
NF-κB gene expression levels, indicating that higher substitution levels of FCSM can induce inflammatory responses in the intestine.
IL-10 is an anti-inflammatory factor which plays a role in down-regulating inflammatory response and antagonizing inflammatory mediators [
25]. Gene expression of the pro-inflammatory factor
IL-10 was significantly higher than that of the control group at 12.5%, 25% and 50% substitution levels, suggesting that intestinal inflammation improves at appropriate substitution ranges.
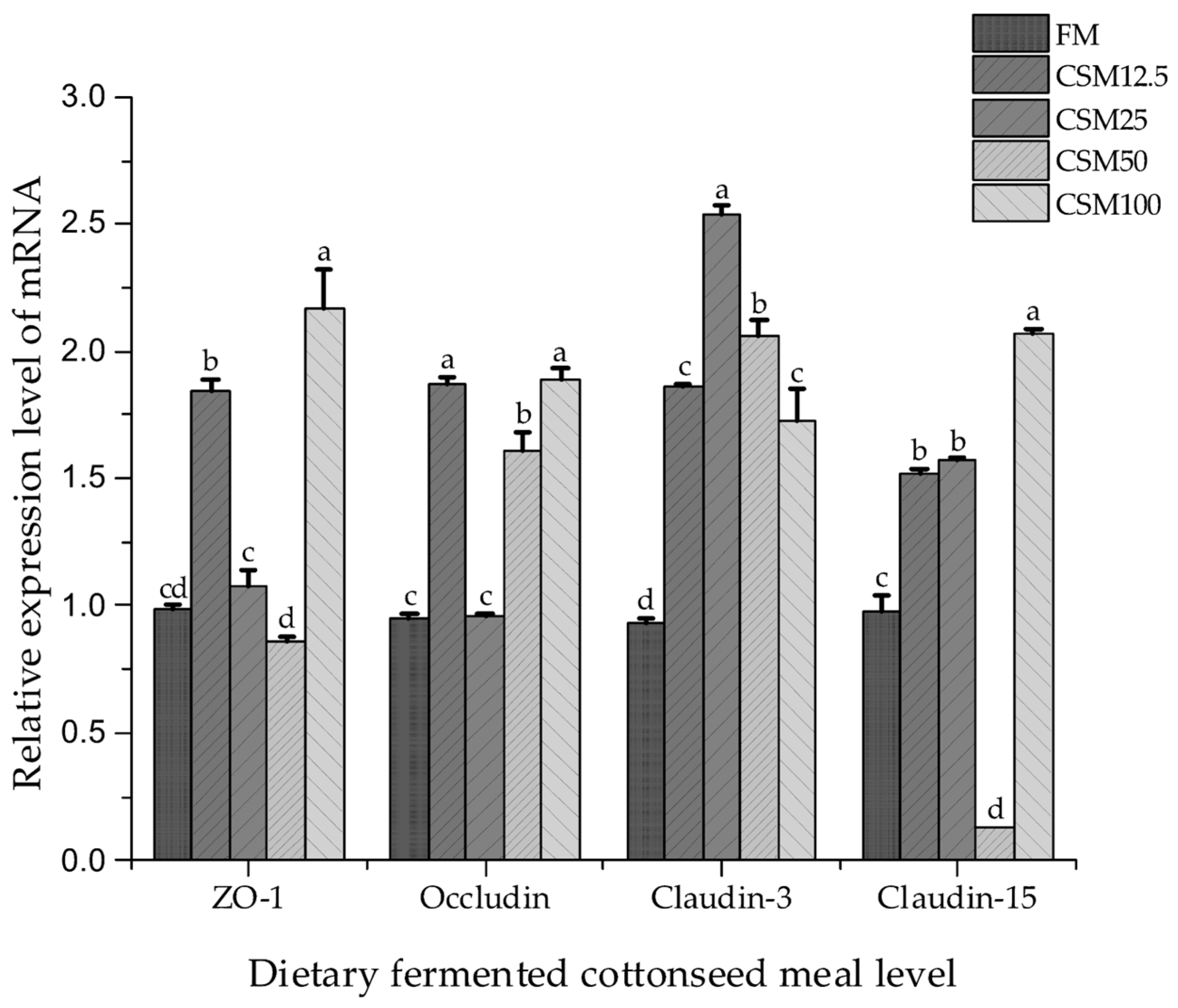
The intestinal tract in fish is widely recognized as a significant physical barrier, and maintaining its structural integrity is essential for effectively resisting foreign antigens [
26]. The presence of compromised or diminished physical barrier function within the intestinal tract of fish is associated with an elevated susceptibility to pathogen infection and a hindered ability to suppress pathogen growth [
12].
ZO-1 is involved, in its function as a scaffolding protein, by determining the tightness of the epithelial barrier and contributing to the polarity of the epithelial cells [
5]. The extracellular loop of
Occludin goes straight into the TJ complex and interacts with the transmembrane domain to alter the permeability of extracellular selectivity [
27]. In the present study, the mRNA transcript levels of
ZO-1 were up-regulated, but the relative expression levels of the
Occludin gene in the CSM25 and FM groups were not significantly different.
Claudin-3 is a tightening TJ protein that aids in sealing the paracellular intestinal barrier [
28]. In this study, the
Claudin-3 gene had the highest relative expression in the CSM25 group, indicating that the intestinal barrier closure was strongest at the 25% substitution level.
Claudin-15 is widely distributed in the tight junctions of the villi and crypt cells of the small and large intestines. In this study, the relative expression of
Claudin-15 in the CSM12.5, CSM25, and CSM100 groups was significantly higher than that in the FM group, suggesting that the structural integrity of intestinal intercellular structures was better in the CSM12.5, CSM25, and CSM100 groups. The results of the above studies suggest that appropriate substitution ratios can ameliorate golden pompano intestinal physical barrier damage induced by FCSM. This is similar to the results of a study on the genes related to the improvement of intestinal tight junction in yellow catfish by fermented rapeseed meal [
29]. Therefore, FCSM may alleviate the damage, associated with cottonseed meal, to the intestinal structure of golden pompano by degrading anti-nutritional factors, thereby improving the intestinal health of fish.
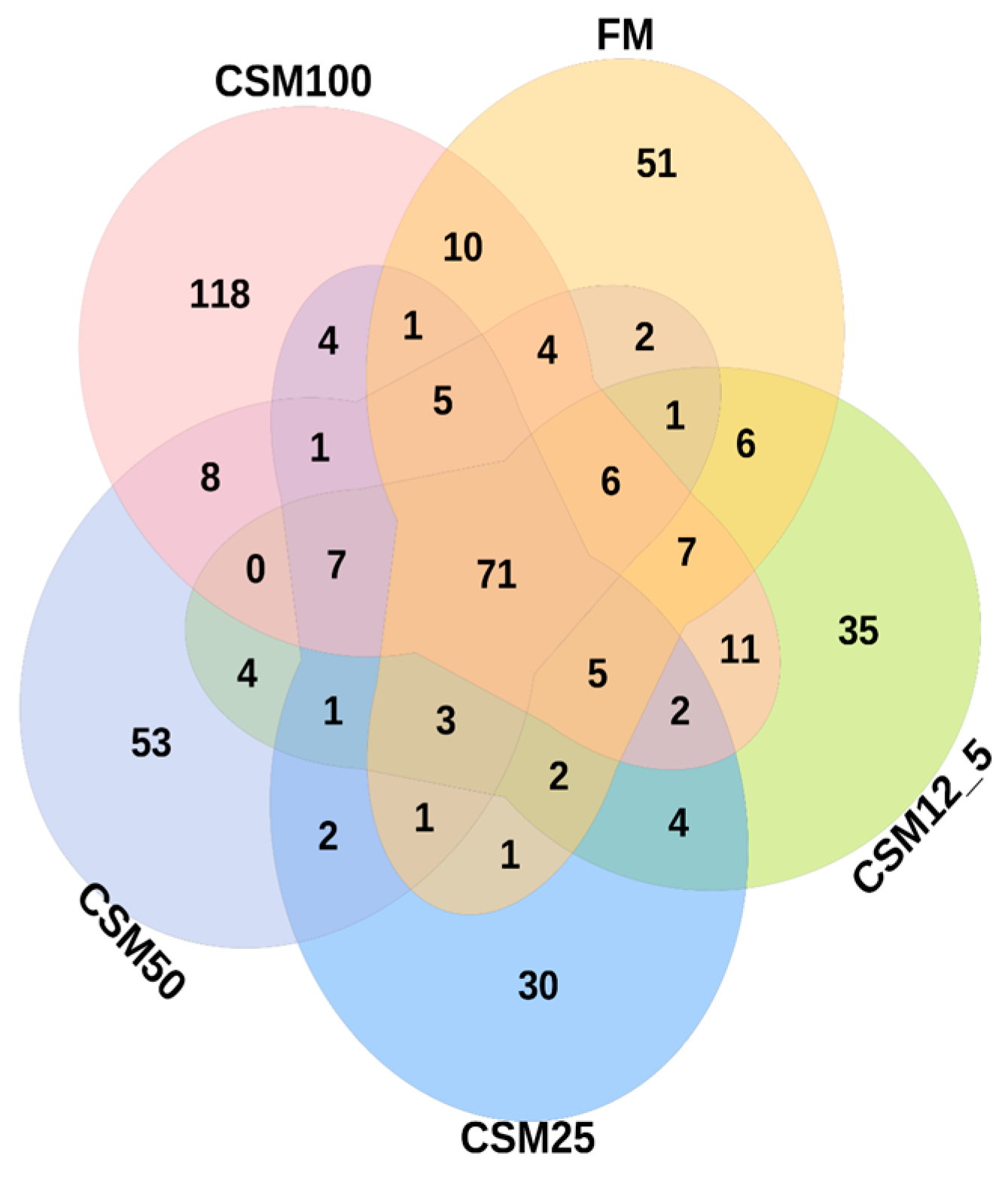
The microbial ecosystem of animals consists of microbial communities and their host microenvironment. The intestinal microenvironment consists of intestinal microorganisms and is the most critical microenvironment of the animal body [
30]. The intestinal microbiota play a crucial role in various physiological processes, including metabolism, nutrient absorption, growth and development, and immune function. Therefore, it is very important for body health to investigate changes in fish intestinal flora. In the present study, the OTU Wayne diagram found that 71 OTU were common in all groups, indicating that there are still inherent core flora at different levels of FCSM addition in the same culture environment. These results suggested that the tested variations in diet did not exert large, long-term alterations on the intestinal microbiota of golden pompano.
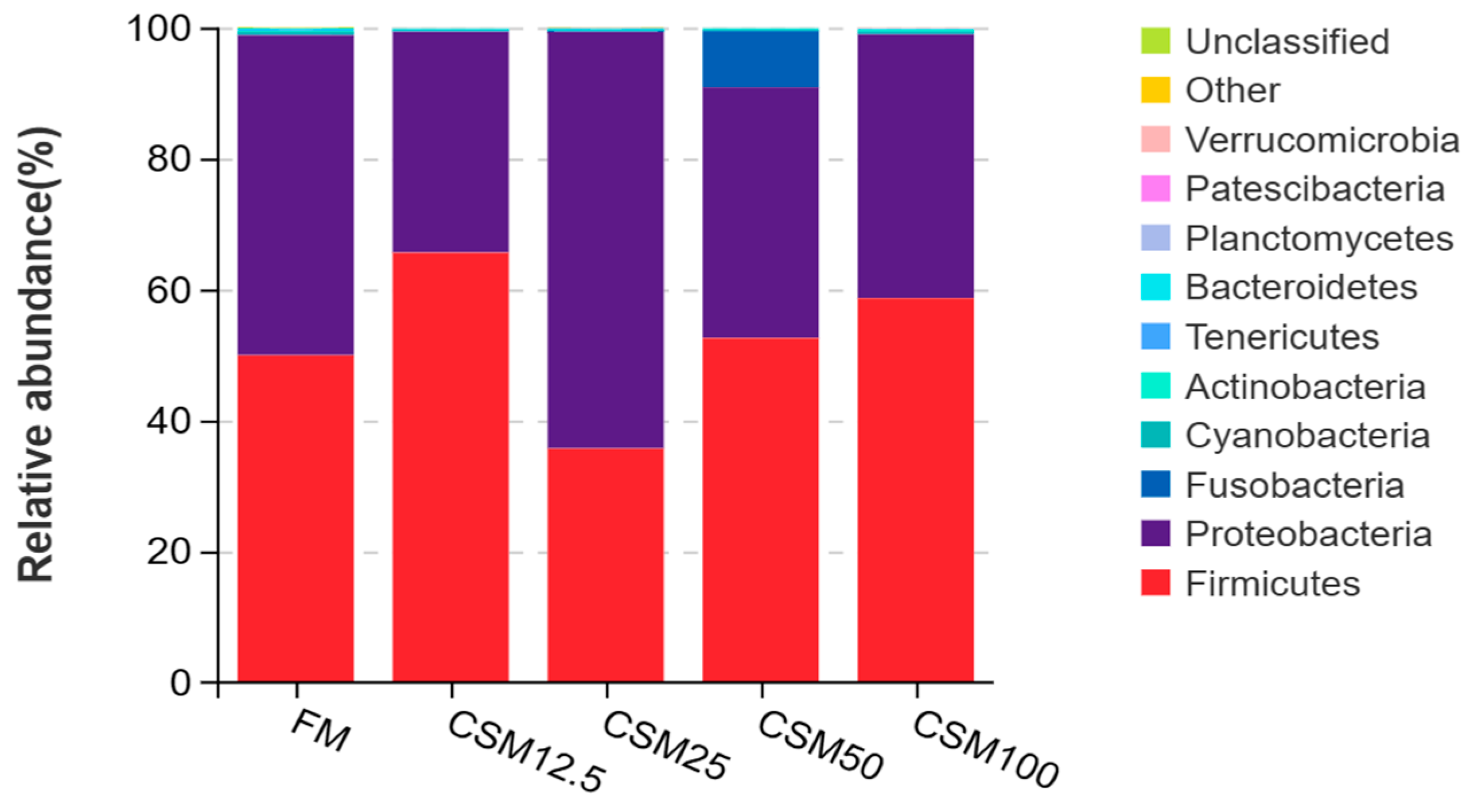
Intestinal bacterial microflora level is closely related to digestive function and overall gastrointestinal health [
31]. In terms of community composition, we found that the dominant intestinal bacteria of the golden pompano belonged to three phyla: Proteobacteria, Firmicutes, and Tenercutes [
32]; this has also been reported for the turbot [
33].
Proteobacteria can catabolize feed ingredients [
34]. The increased abundance may allow
T. ovatus to absorb nutrients from the feed. Proteobacteria, a class of bacteria characterized by their Gram-negative cell wall structure, serves a significant function in the breakdown and fermentation of polysaccharides, proteins, and various organic substances. Moreover, they constitute the prevailing microbial population within the intestinal ecosystem of numerous fish species [
35]. It is well known that many kinds of Vibrio, Photobacterium, and Mycoplasma are pathogenic bacteria [
19,
36]. These microorganisms are found within the gastrointestinal tract. The degradation of the aquatic environment leads to an escalation in nutrient deficits and physiological dysfunctions, hence heightening the vulnerability of fish to these infections [
37]. Research shows that the balance of the intestinal microflora was altered primarily by probiotic supplementation and, to a lesser extent, by the energy content of the diet [
33]. The majority of Fusobacteriales strains identified in fish intestinal samples are attributed to Cetobacterium, a prevalent and extensively distributed species found within the gastrointestinal tracts of fish [
38]. In this experiment, the dominant intestinal flora of the golden pompano was composed of
Firmicutes and
proteobacteria. However, with the change in the proportion of FCSM instead of fishmeal in feed, the absolute dominant flora between the groups also changed. At the phylum level, the dominant phylum of the control group for the CSM12.5 group, CSM50 group, and CSM100 group was firmicutes, while the dominant phylum of the CSM25 group was proteobacteria. The results of the above studies showed that the substitution of FM by FCSM did not lead to changes in the dominant strains of the intestinal flora of golden pompano.
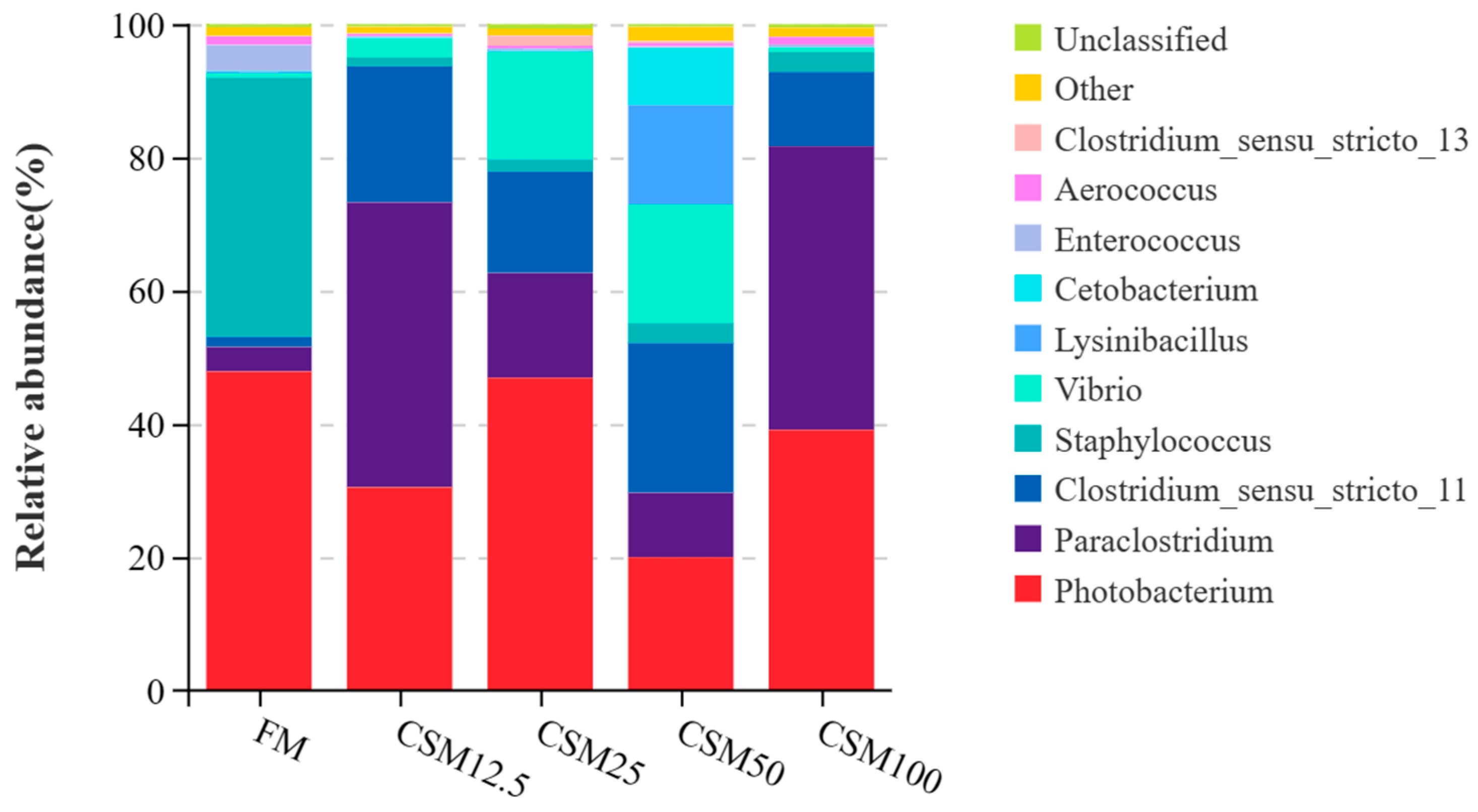
At the genus level, the dominant genera in the intestinal flora of the
T. ovatus are
Photobacterium and
Paraclostridium.
Photobacterium, a bacterium classified as Gram-negative, has a combination of anaerobic and aerobic characteristics, displaying both respiratory and fermentative metabolic pathways. The inclusion of Bacillus probiotics in the diet has been found to promote the proliferation of lactic acid bacteria within the gastrointestinal system, leading to a decrease in pH levels. Consequently, this creates an unfavorable environment for the growth and survival of pathogenic bacteria, such as
E. coli and
Salmonella spp. [
39]. Photobacterium, a Gram-negative bacterium, is partly anaerobic and has respiratory and fermentative metabolism types. Photobacterium can infect the shrimp Litopenaeus vannamei to cause its mortality, and may be a common pathogen of shrimps [
19]. The Photobacterium levels in the CSM25 group and CSM50 group were significantly lower than that in the control group, indicating that the abundance of Photobacterium decreased when the amount of FCSM added to the feed was 25–50% [
40].
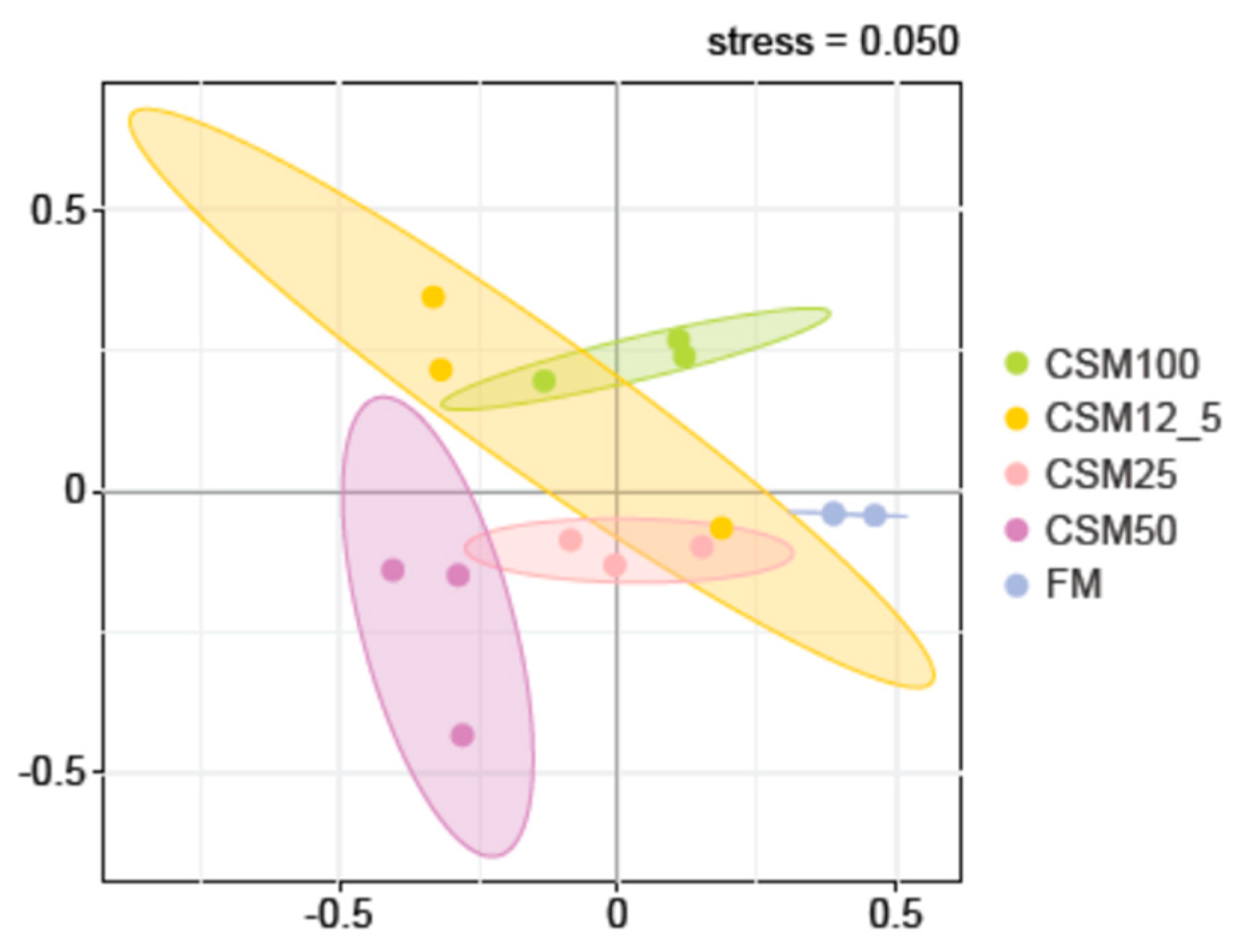
Beta diversity analysis is mainly used to compare differences in the overall structure of microbial communities in each sample. Non-metric multidimensional scaling (NMDS) is a data analysis method that simplifies the research objects of multidimensional space to low-dimensional space for locating, analysis, and classification while retaining the original relationship between objects [
41]. In this experiment, we found that the gut microbial composition of the FM group was significantly different from that of the CSM12.5, CSM25, CSM50, and CSM100 groups, and that there were significant differences in the composition of the CSM12.5 and CSM50 groups and the CSM25 and CSM100 groups, indicating that varying levels of fermented cotton meal substitution in the feeds resulted in significant differences in gut microbial composition. Complex microbiota interactions may explain the fact that fermented fishmeal increased the diversity of the gut microbiota of the vanabin carp, a finding which is similar to the results of our study [
42].


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}