
Spatial and Temporal Distribution of Rodents during the Epizootic and Enzootic Periods of Plague, with a Focus on Exu, Northeastern Brazil

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
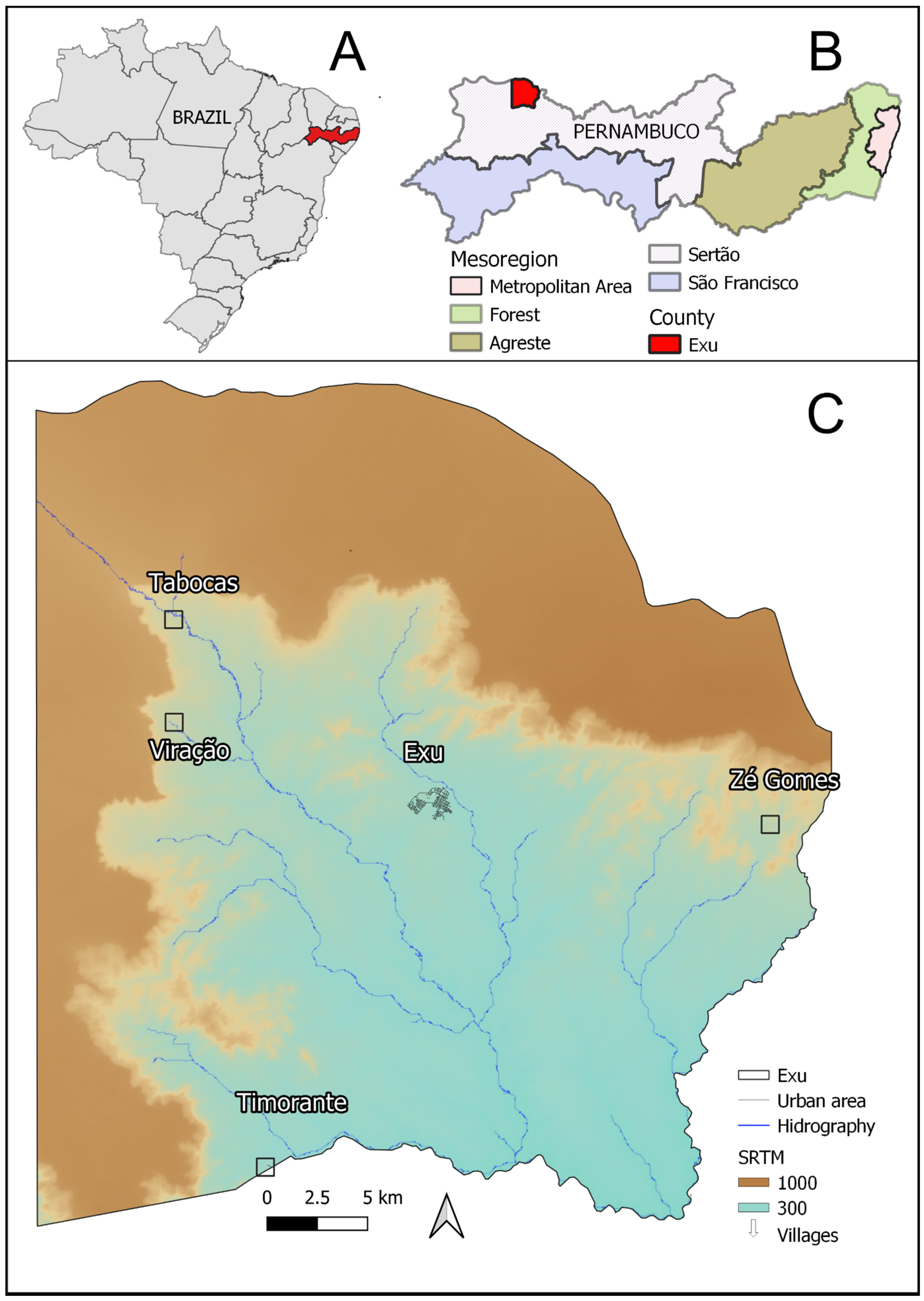
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
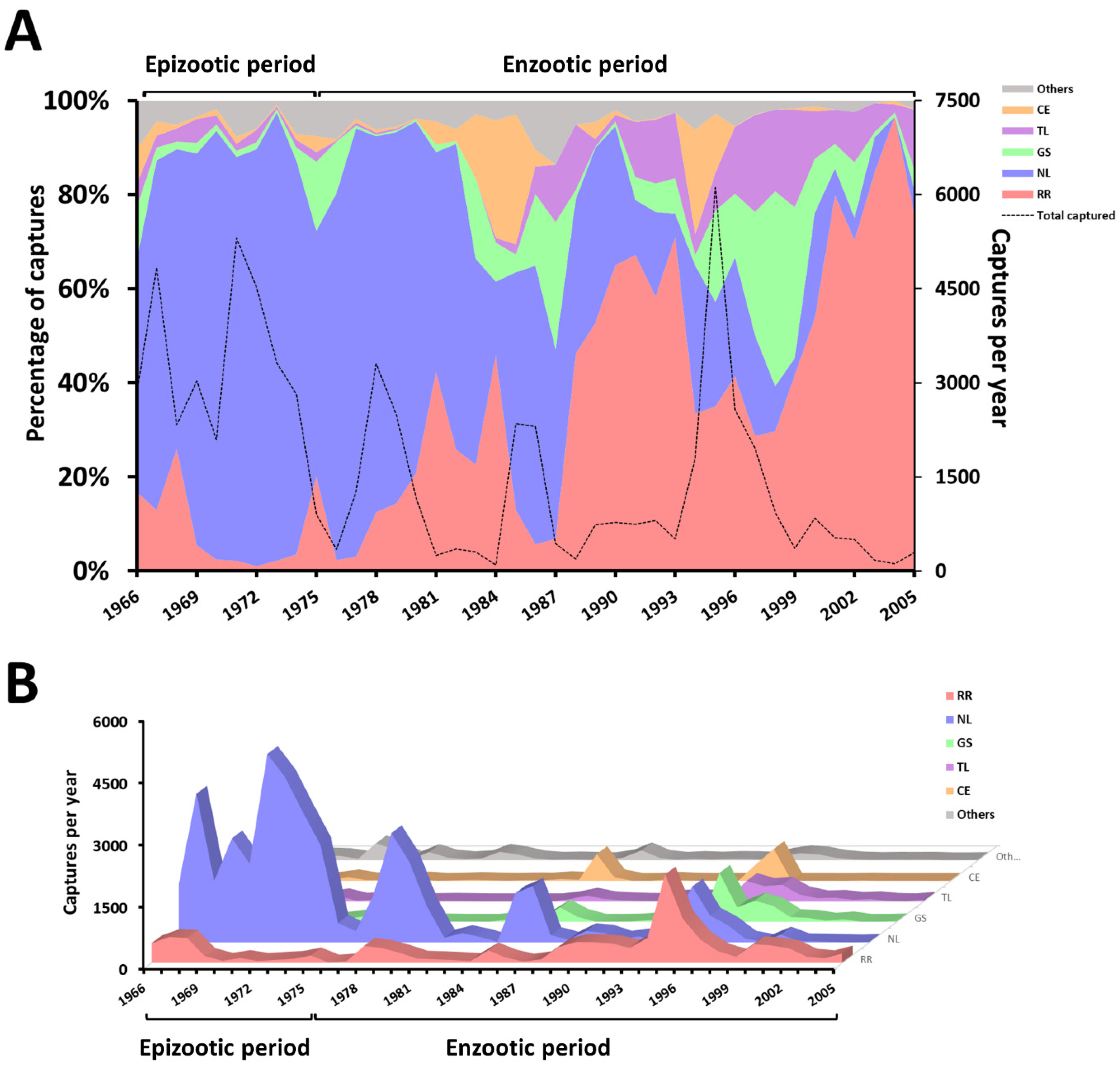
3.1. Fluctuation in the Abundance of Rodent Species from 1966 to 2005
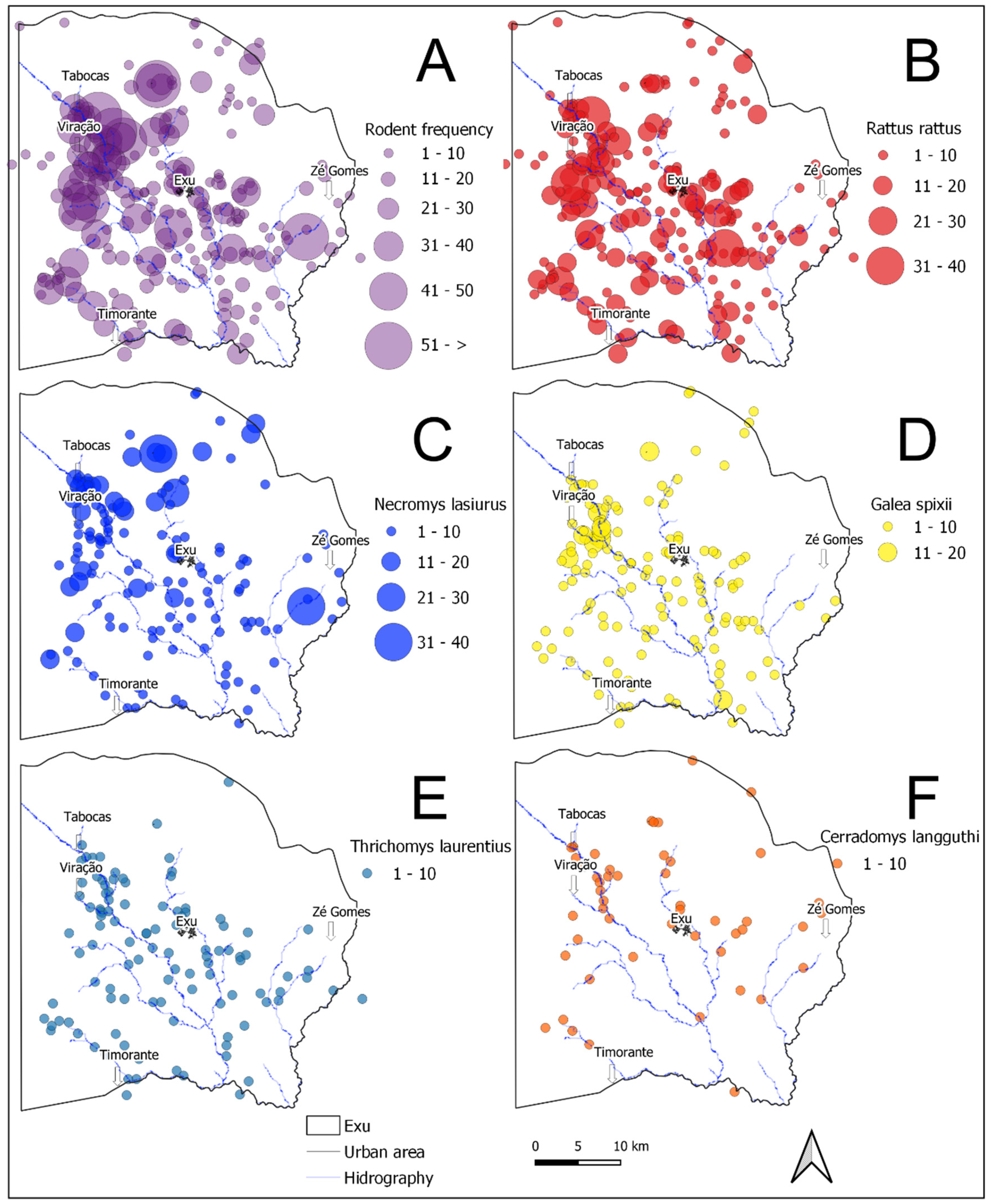
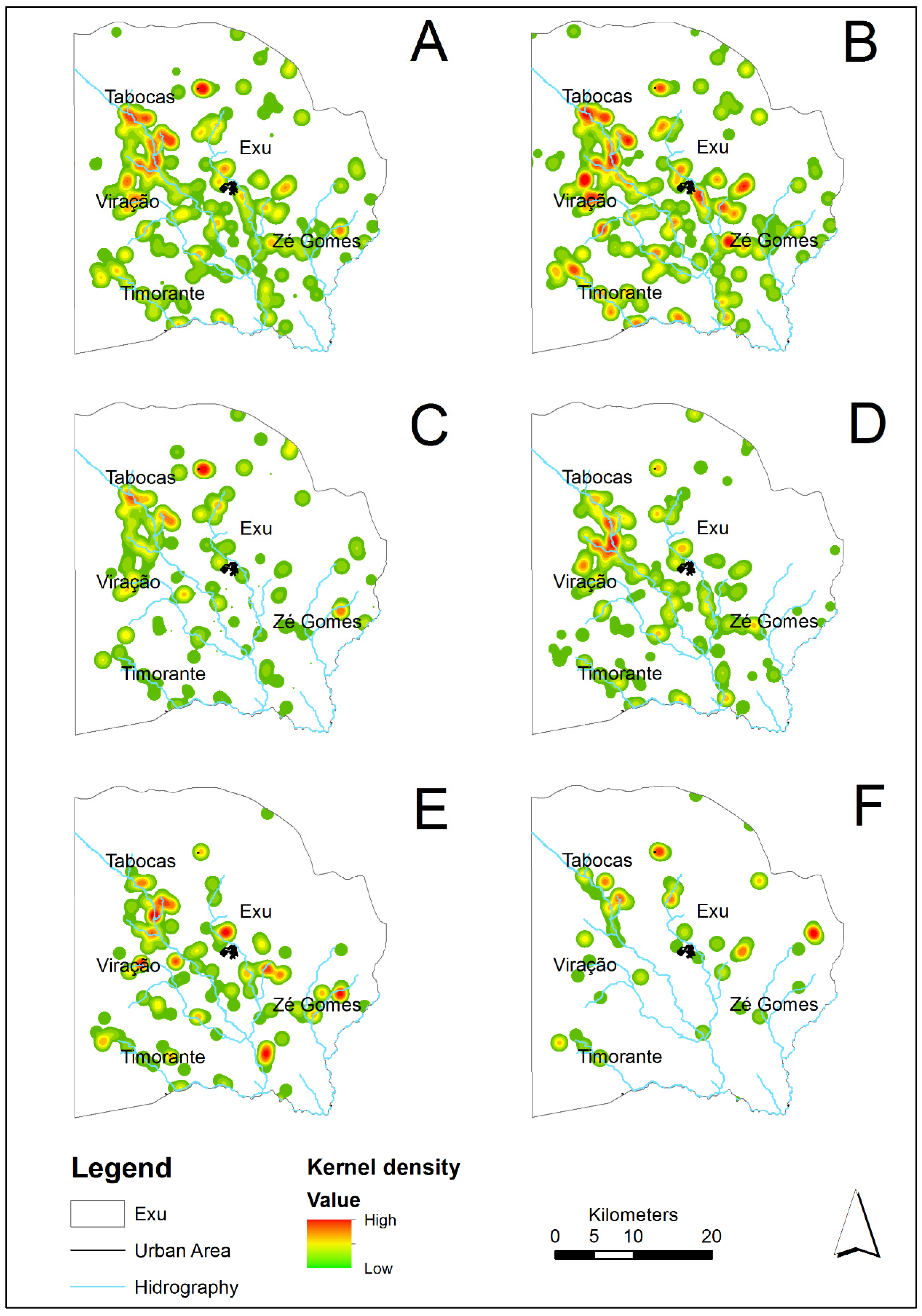
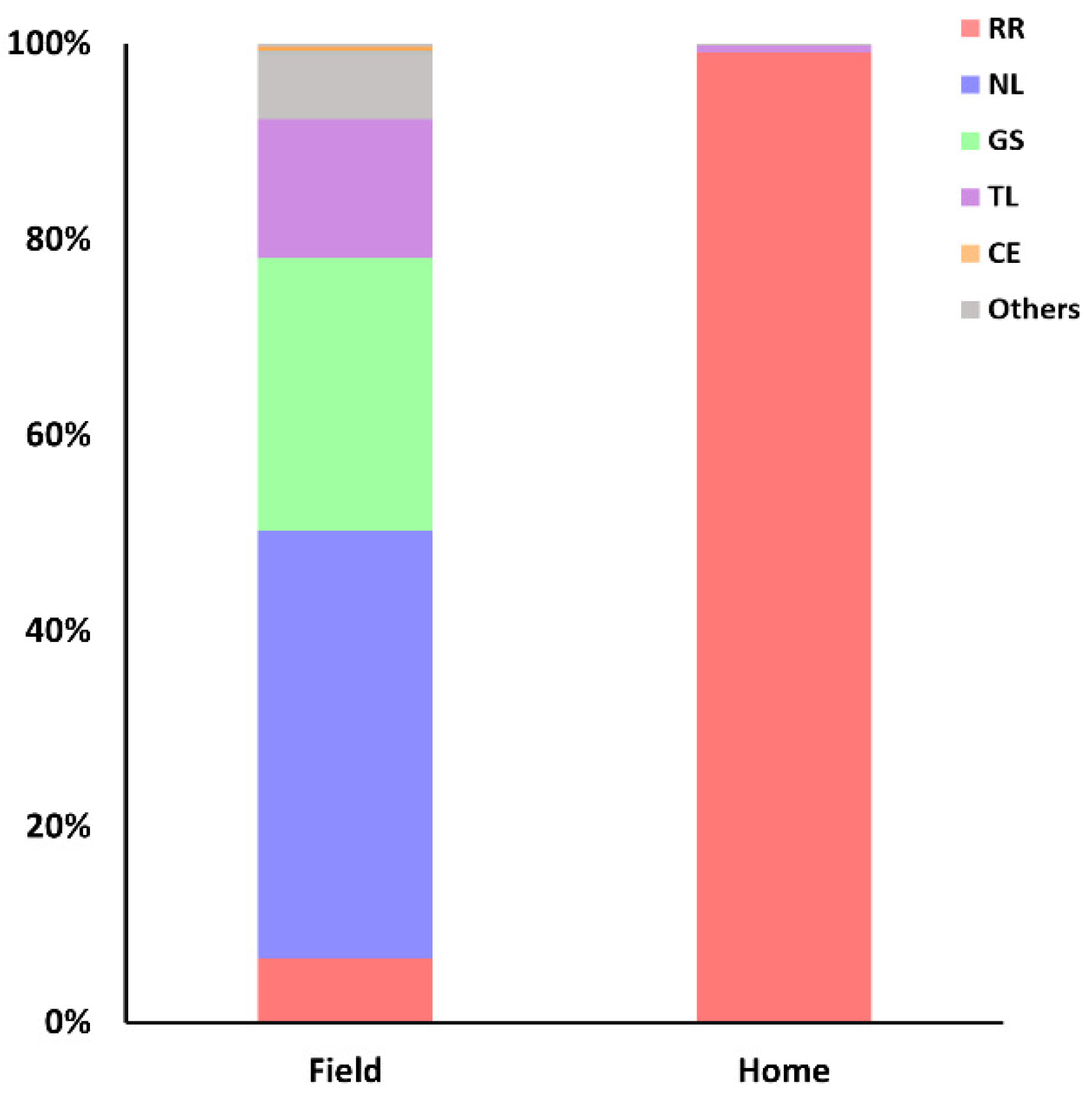
3.2. Spatial Distribution of the Rodents’ Populations in the Period Analyzed (1996–2005)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbieri, R.; Signoli, M.; Chevé, D.; Costedoat, C.; Tzortzis, S.; Aboudharam, G.; Raoult, D.; Drancourt, M. Yersinia pestis: The Natural History of Plague. Clin. Microbiol. Rev. 2020, 34, e00044-19. [Google Scholar] [CrossRef]
- D’Elía, G.; Fabre, P.H.; Lessa, E.P. Rodent systematics in an age of discovery: Recent advances and prospects. J. Mammal. 2019, 100, 852–871. [Google Scholar] [CrossRef]
- Mahmoudi, A.; Krytufek, B.; Sludsky, A.; Schmid, B.V.; Almeida, A.M.P.; Lei, X.; Bertherat, E.; Ramasindrazana, B.; Yeszhanov, A.; Stenseth, N.C.; et al. Plague reservoir species throughout the world. Integr. Zool. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, B.; Stenseth, N.C.; Walløe, L.; Lei, X. Plague: A disease which changed the path of human civilization. In Yersinia Pestis: Retrospective and Perspective, 1st ed.; Yang, R., Anisimov, A., Eds.; Springer: Dordrecht, The Netherlands, 2016; Volume 918, pp. 1–26. [Google Scholar]
- WHO. Plague around the world in 2019. Wkly. Epidemiol. Rec. 2019, 94, 289–292. [Google Scholar]
- Schneider, M.C.; Najera, P.; Aldighieri, S.; Galan, D.I.; Bertherat, E.; Ruiz, A.; Dumit, E.; Gabastou, J.M.; Espinal, M.A. Where Does Human Plague Still Persist in Latin America? PLoS Negl. Trop. Dis. 2014, 8, e2680. [Google Scholar] [CrossRef] [PubMed]
- Pollitzer, R.; Meyer, K.F. Plague in the Americas; Scient Publ Nº. 115; PAHO: Washington, DC, USA, 1965. [Google Scholar]
- Almeida, A.M.P.; Sobreira, M.; Leal, N.C.; Tavares, C. Does the plague still threaten us? Rev. Soc. Bras. Med. Trop. 2020, 53, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Giles, J.; Peterson, A.T.; Almeida, A.M.P. Ecology and Geography of Plague Transmission Areas in Northeastern Brazil. PLoS Negl. Trop. Dis. 2011, 5, e925. [Google Scholar] [CrossRef] [PubMed]
- Baltazard, M. La démarche exemplaire d’un épidémiologiste de terrain: M. Baltazard et les foyers de peste du nordest brésilien. Bull. Soc. Pathol. Exot. 2004, 97, 93–118. [Google Scholar]
- Fernandes, D.L.R.; Gomes, E.C.S.; Filgueira, M.B.; Guimarães, R.J.P.S.; Almeida, A.M.P. Spatiotemporal analysis of bubonic plague in Pernambuco, Northeast of Brazil: Case study in the Municipality of Exu. PLoS ONE 2021, 16, e0249464. [Google Scholar] [CrossRef]
- Almeida, C.R.; Almeida, A.M.P.; Brasil, D.P. Observations sur le comportament de fouissement de Zygodontomys lasiurus pixuna Moojen, 1943. Reproduction au laboratoire (Rongeurs, Cricétidés). Mammalia 1981, 45, 415–422. [Google Scholar] [CrossRef]
- Almeida, C.R.; Almeida, A.M.P.; Brasil, D.P.; Sobrinho, J.D.; Leal, N.C. Estudo do roedor Akodon arviculoides, Wagner, 1842 (Cricetidae). Importância nos focos pestosos do Brasil. Memórias do Instituto Oswaldo Cruz 1986, 81, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Bonvicino, C.R.; Oliveira, J.A.; Estrela, P.C.; D’Andrea, P.S.; Almeida, A.M.P. A taxonomic update of small mammal plague reservoirs in South America. Vector-Borne Zoonotic Dis. 2015, 15, 571–579. [Google Scholar] [CrossRef]
- Karimi, Y.; Almeida, C.R.; Almeida, A.M.P. La peste expérimentale chez les rongeurs du Brésil. Déductions Épidémiologiques. Bull. Soc. Pathol. Exot. 1974, 67, 591–601. [Google Scholar]
- Karimi, Y.; Eftekhari, M.; Almeida, C.R. Sur l’écologie des puces impliquées dans l’epidémiologie de la peste et le rôle éventuel de certains insectes hématophages dans son processus au Nord-Est du Brésil. Bull. Soc. Pathol. Exot. 1974, 67, 583–591. [Google Scholar]
- Tavares, C.; Aragão, A.I.; Leal, N.C.; Leal-Balbino, T.C.A.; Oliveira, M.B.M.; Oliveira, G.M.; Almeida, A.M.P. Plague in Brazil: From now and then. Adv. Exp. Med. Biol. 2012, 954, 69–77. [Google Scholar] [PubMed]
- Baltazard, M.; Davis, D.H.; Devignat, R.; Girard, G.; Gohar, M.A.; Kartman, L.; Meyer, K.F.; Parker, M.T.; Pollitzer, R.; Prince, F.M.; et al. Recommended laboratory methods for the diagnosis of plague. Bull. World Health Organ. 1956, 14, 457–509. [Google Scholar] [PubMed]
- Bahmanyar, M.; Cavanaugh, D.C. Plague Manual; World Health Organization: Geneva, Switzerland, 1976. [Google Scholar]
- Dennis, D.T.; Gage, K.L.; Gratz, N.G.; Poland, J.D.; Tikhomirov, E. Plague Manual: Epidemiology, Distribution, Surveillance and Control; World Health Organization: Geneva, Switzerland, 1999; pp. 2–171. [Google Scholar]
- Mills, J.N.; Yates, T.L.; Childs, J.E.; Parmenter, R.R.; Ksiazek, T.G.; Rollin, P.E.; Peters, C.J. Guidelines for working with rodents potentially infected with hantavirus. J. Mammal. 1995, 76, 716–722. [Google Scholar] [CrossRef] [Green Version]
- da Costa, E.C.V.; Sobreira, M.; Leal, N.C.; Almeida, A.M.P. Rodents and other small mammal reservoirs in plague foci in northeastern Brazil. J. Infect. Dev. Ctries. 2017, 11, 426–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeppelini, C.G.; Almeida, A.M.P.; Estrela, P.C. Ongoing quiescence in the Borborema Plateau Plague focus (Paraiba, Brazil). An. Acad. Bras. Ciênc. 2018, 90, 3007–3015. [Google Scholar] [CrossRef] [PubMed]
- IBGE. Brazilian Institute of Geography and Statistics. Available online: https://www.ibge.gov.br/geociencias/organizacao-do-territorio/15774-malhas.html?=&t=downloads (accessed on 1 April 2021).
- CPRM. Mineral Resources Research Company. Available online: https://www.cprm.gov.br/en/Hydrology-83 (accessed on 1 April 2021).
- SRTM. Shuttle Radar Topography Mission. Available online: https://code.earthengine.google.com/ccf3b9ff46eb845e1b88f68550e9a22a (accessed on 1 April 2021).
- Qgis Desktop Software Version 3.16.5. Available online: https://www.qgis.org (accessed on 24 May 2021).
- ArcGIS Software Version 10. Available online: http://www.arcgis.com (accessed on 24 May 2021).
- Almeida, A.M.P.; Leal, N.C.; Carvalho, F.G.; Sobrinho, J.D.; Almeida, C.R. Plague surveillance in Brazil: 1983–1992. Rev. Inst. Med. Trop. Sao Paulo 1995, 37, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Biggins, D.E.; Eads, D.A. Prairie Dogs, Persistent Plague, Flocking Fleas, and Pernicious Positive Feedback. Front. Vet. Sci 2019, 6, 75. [Google Scholar] [CrossRef]
- Davis, S.; Calvet, E.; Leirs, H. Fluctuating Rodent Populations and Risk to Humans from Rodent-Borne Zoonoses. Vector-Borne Zoonotic Dis. 2005, 5, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Gage, K.; Kosoy, M.K. Natural history of plague: Perspectives from more than a Century of Research. Annu. Rev. Entomol. 2005, 50, 505–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Instituto Chico Mendes de Conservação da Biodiversidade. Brazil Red Book of Threatened Species of Fauna (Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume II—Mamíferos), 1st ed.; Instituto Chico Mendes de Conservação da Biodiversidade: Brasília, Brazil, 2018; pp. 15–622. [Google Scholar]
- Fernandes, D.L.R.; Filgueira, M.B.; Sobreira, M.; Leal, N.C.; Reis, C.R.S.; Almeida, A.M.P. Rodent hosts and flea vectors in Brazilian plague foci: A review. Int. Zool. 2020, 16, 1–10. [Google Scholar]
- Biggins, D.E.; Kosoy, M.Y. Influences of introduced plague on North American mammals: Implications from ecology of plague in Asia. J. Mammal. 2001, 82, 906–916. [Google Scholar] [CrossRef] [Green Version]
- Hotez, P.J. Global urbanization and the neglected tropical diseases. PLoS Negl. Trop. Dis. 2017, 11, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Himsworth, C.G.; Parsons, K.L.; Jardine, C.; Patrick, D.M. Rats, Cities, People, and Pathogens: A Systematic Review and Narrative Synthesis of Literature Regarding the Ecology of Rat-Associated Zoonoses in Urban Centers. Vector-Borne Zoonotic Dis. 2013, 13, 349–359. [Google Scholar] [CrossRef]
- Butler, T.; Fu, Y.S.; Furman, I.L.; Almeida, C.; Almeida, A. Experimental Yersinia pestis infection in rodents after intragastric inoculation and ingestion of bacteria. Infect. Immun. 1982, 36, 1160–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutinho, E.M.; Almeida, A.M.P.; Almeida, C.R. Histopatologia da infecção por Yersinia pestis em roedores de focos de peste do nordeste brasileiro. Mem. Inst. Oswaldo Cruz 1982, 77, 139–151. [Google Scholar] [CrossRef]
- Petter, F. Les rongeurs et la peste en Iran et au Brésil. Nouvelles donnés. Bull. Soc. Pathol. Exot. 1999, 92, 411–413. [Google Scholar] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, D.L.R.d.S.; Bezerra, M.F.; Duarte, B.M.; de França Silva, M.P.; de Almeida Souza, H.; de Souza Gomes, E.C.; de Almeida, A.M.P. Spatial and Temporal Distribution of Rodents during the Epizootic and Enzootic Periods of Plague, with a Focus on Exu, Northeastern Brazil. Trop. Med. Infect. Dis. 2021, 6, 195. https://doi.org/10.3390/tropicalmed6040195
Fernandes DLRdS, Bezerra MF, Duarte BM, de França Silva MP, de Almeida Souza H, de Souza Gomes EC, de Almeida AMP. Spatial and Temporal Distribution of Rodents during the Epizootic and Enzootic Periods of Plague, with a Focus on Exu, Northeastern Brazil. Tropical Medicine and Infectious Disease. 2021; 6(4):195. https://doi.org/10.3390/tropicalmed6040195
Chicago/Turabian StyleFernandes, Diego Leandro Reis da Silva, Matheus Filgueira Bezerra, Bruna Mendes Duarte, Mayara Paes de França Silva, Hadassa de Almeida Souza, Elainne Christine de Souza Gomes, and Alzira Maria Paiva de Almeida. 2021. "Spatial and Temporal Distribution of Rodents during the Epizootic and Enzootic Periods of Plague, with a Focus on Exu, Northeastern Brazil" Tropical Medicine and Infectious Disease 6, no. 4: 195. https://doi.org/10.3390/tropicalmed6040195
APA StyleFernandes, D. L. R. d. S., Bezerra, M. F., Duarte, B. M., de França Silva, M. P., de Almeida Souza, H., de Souza Gomes, E. C., & de Almeida, A. M. P. (2021). Spatial and Temporal Distribution of Rodents during the Epizootic and Enzootic Periods of Plague, with a Focus on Exu, Northeastern Brazil. Tropical Medicine and Infectious Disease, 6(4), 195. https://doi.org/10.3390/tropicalmed6040195