
Immunological Characteristics of Hepatic Dendritic Cells in Patients and Mouse Model with Liver Echinococcus multilocularis Infection

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. Mice
2.3. Hepatic Mouse Model of E. multilocularis Infection
2.4. Flow Cytometric Analysis
2.5. Statistical Analysis
3. Results
3.1. scRNA-seq Reveals the Distribution and Molecular Expression of DC Subsets in the Hepatic AE Tissue Microenvironment
3.2. Changes in the Hepatic DC Composition in Mouse Livers during E. multilocularis Infection
3.3. Expression of Costimulatory Molecules on Hepatic DC Subsets in Mouse Livers during E. multilocularis Infection
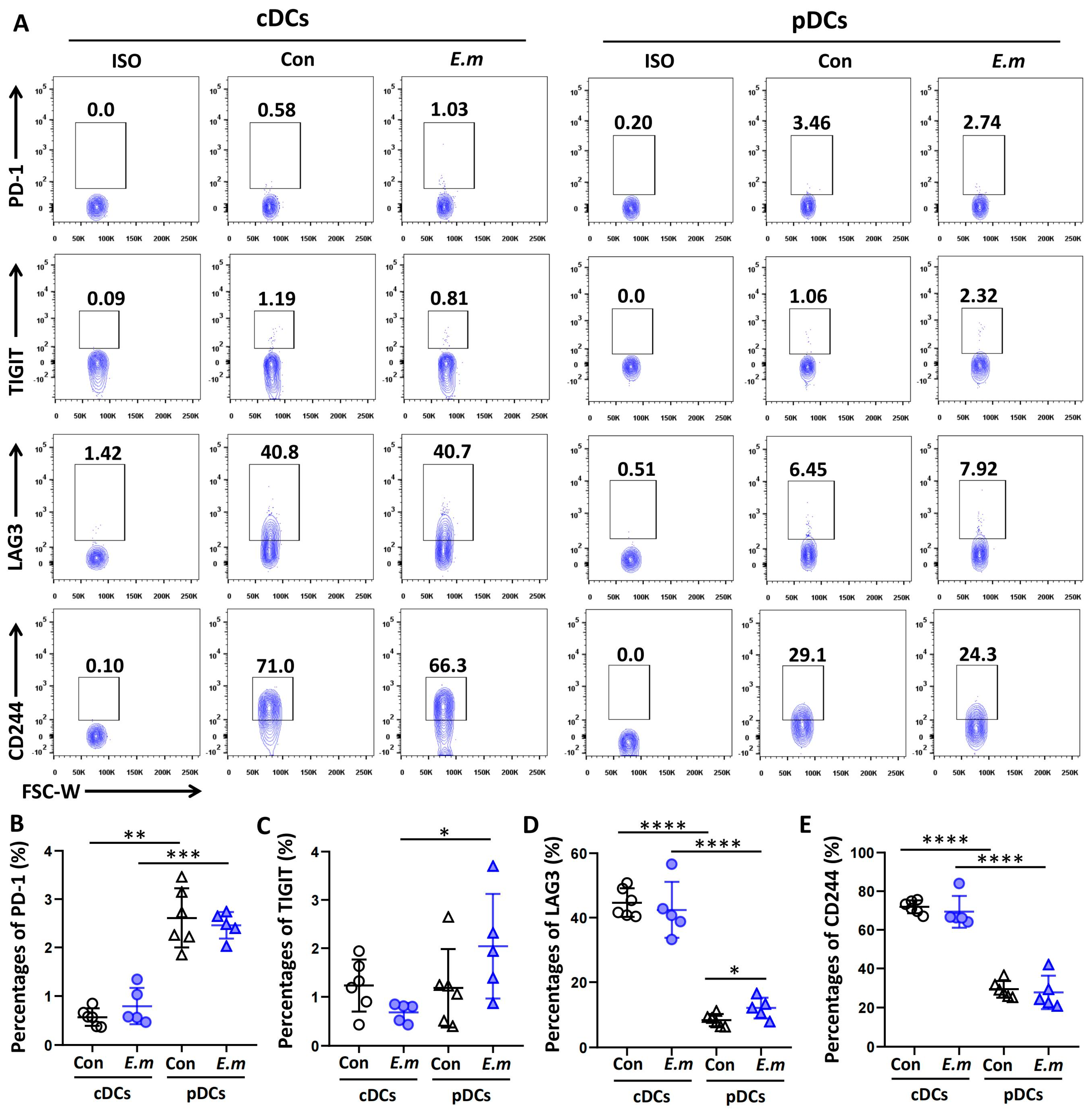
3.4. Expression of Immune Checkpoint Molecules on Hepatic DC Subsets in E. multilocularis-Infected Mice
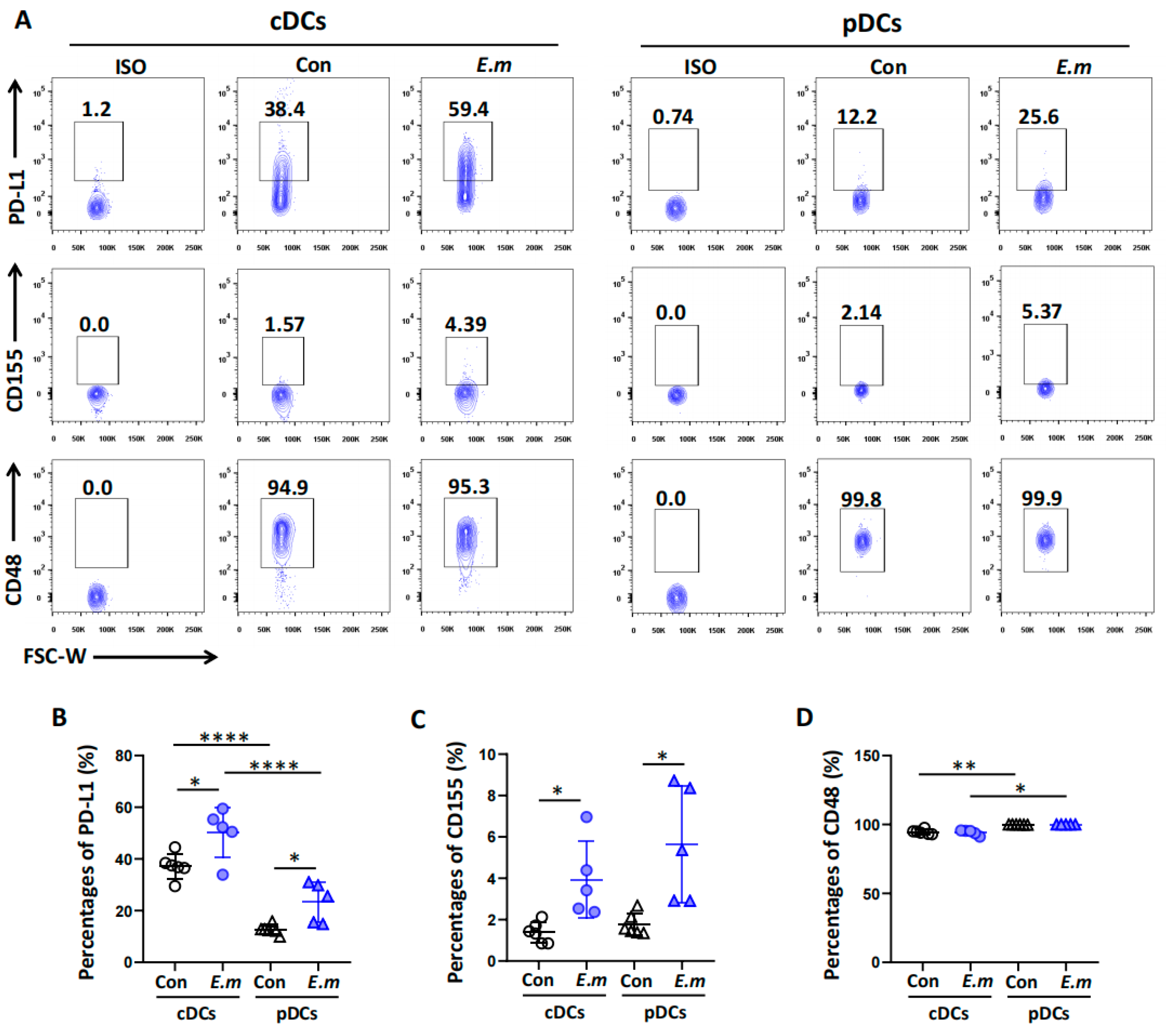
3.5. Expression of Immune Checkpoint Ligands on Hepatic DC Subsets in E. multilocularis-Infected Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wen, H.; Vuitton, L.; Tuxun, T.; Li, J.; Vuitton, D.A.; Zhang, W.; McManus, D.P. Echinococcosis: Advances in the 21st Century. Clin. Microbiol. Rev. 2019, 32, e00075-18. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Autier, B.; Gottstein, B.; Millon, L.; Ramharter, M.; Gruener, B.; Bresson-Hadni, S.; Dion, S.; Robert-Gangneux, F. Alveolar echinococcosis in immunocompromised hosts. Clin. Microbiol. Infect. 2023, 29, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Gottstein, B.; Soboslay, P.; Ortona, E.; Wang, J.; Siracusano, A.; Vuitton, D. Immunology of Alveolar and Cystic Echinococcosis (AE and CE). Adv. Parasitol. 2017, 96, 1–54. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, P.R.; Keller, K.; Magnotta, M.; Ragland, N. The global burden of alveolar echinococcosis. PLoS Negl. Trop. Dis. 2010, 4, e722. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Aji, T.; Dong, J.H.; Shao, Y.M.; Zhao, J.M.; Li, T.; Tuxun, T.E.H.J.; Shalayiadang, P.; Ran, B.; Jiang, T.-M.; Zhang, R.-Q.; et al. Ex vivo liver resection and autotransplantation as alternative to allotransplantation for end-stage hepatic alveolar echinococcosis. J. Hepatol. 2018, 69, 1037–1046. [Google Scholar] [CrossRef]
- Sailer, M.; Soelder, B.; Allerberger, F.; Zaknun, D.; Feichtinger, H.; Gottstein, B. Alveolar echinococcosis of the liver in a six-year-old girl with acquired immunodeficiency syndrome. J. Pediatr. 1997, 130, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Chauchet, A.; Grenouillet, F.; Knapp, J.; Richou, C.; Delabrousse, E.; Dentan, C.; Millon, L.; Di Martino, V.; Contreras, R.; Deconinck, E.; et al. Increased Incidence and Characteristics of Alveolar Echinococcosis in Patients with Immunosuppression-Associated Conditions. Clin. Infect. Dis. 2014, 59, 1095–1104. [Google Scholar] [CrossRef]
- Mejri, N.; Muller, N.; Hemphill, A.; Gottstein, B. Intraperitoneal Echinococcus multilocularis infection in mice modulates peritoneal CD4+ and CD8+ regulatory T cell development. Parasitol. Int. 2011, 60, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lin, R.; Li, Z.; Yang, S.; Bi, X.; Wang, H.; Aini, A.; Zhang, N.; Abulizi, A.; Sun, C.; et al. Immune Exhaustion of T Cells in Alveolar Echinococcosis Patients and Its Reversal by Blocking Checkpoint Receptor TIGIT in a Murine Model. Hepatology 2020, 71, 1297–1315. [Google Scholar] [CrossRef] [PubMed]
- Manfras, B.J.; Reuter, S.; Wendland, T.; Kern, P. Increased activation and oligoclonality of peripheral CD8(+) T cells in the chronic human helminth infection alveolar echinococcosis. Infect. Immun. 2002, 70, 1168–1174. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, C.S.; Shao, Y.M.; Yang, S.T.; Bi, X.J.; Li, L.; Wang, H.; Yang, N.; Li, Z.; Sun, C.; Lü, G.; et al. T cell tolerance and exhaustion in the clearance of role of inoculum size in a quantitative hepatic experimental model. Sci. Rep. 2017, 7, 11153. [Google Scholar] [CrossRef]
- Li, D.; Ainiwaer, A.; Zheng, X.; Wang, M.; Shi, Y.; Rousu, Z.; Hou, X.; Kang, X.; Maimaiti, M.; Wang, H.; et al. Upregulation of LAG3 modulates the immune imbalance of CD4+ T cell subsets and exacerbates disease progression in patients with alveolar echinococcosis and a mouse model. PLoS Pathog. 2023, 19, e1011396. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shi, Q.; Zhuang, F.; Liu, J.T.; Li, N.; Chen, Y.X.; Su, X.B.; Yao, A.-H.; Yao, Q.-P.; Han, Y.; Li, S.-S.; et al. Single-cell analyses reveal functional classification of dendritic cells and their potential roles in inflammatory disease. FASEB J. 2019, 33, 3784–3794. [Google Scholar] [CrossRef]
- MacDonald, A.S.; Maizels, R.M. Alarming dendritic cells for Th2 induction. J. Exp. Med. 2008, 205, 13–17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vuitton, D.A.; Gottstein, B. Echinococcus multilocularis and its intermediate host: A model of parasite-host interplay. J. Biomed. Biotechnol. 2010, 2010, 923193. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nutt, S.L.; Chopin, M. Transcriptional Networks Driving Dendritic Cell Differentiation and Function. Immunity 2020, 52, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Bruno, L. Differentiation of dendritic cell subsets from mouse bone marrow. Methods Mol. Biol. 2007, 380, 47–57. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhu, X.; Shi, Z.; Wu, T.; Wu, L. Metabolic Regulation of Dendritic Cell Differentiation. Front. Immunol. 2019, 10, 410. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Amon, L.; Lehmann, C.H.K.; Baranska, A.; Schoen, J.; Heger, L.; Dudziak, D. Transcriptional control of dendritic cell development and functions. Int. Rev. Cell Mol. Biol. 2019, 349, 55–151. [Google Scholar] [CrossRef] [PubMed]
- Tel, J.; Schreibelt, G.; Sittig, S.P.; Mathan, T.S.; Buschow, S.I.; Cruz, L.J.; Lambeck, A.J.A.; Figdor, C.G.; de Vries, I.J.M. Human plasmacytoid dendritic cells efficiently cross-present exogenous Ags to CD8+ T cells despite lower Ag uptake than myeloid dendritic cell subsets. Blood 2013, 121, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Hawiger, D.; Nussenzweig, M.C. Tolerogenic dendritic cells. Annu. Rev. Immunol. 2003, 21, 685–711. [Google Scholar] [CrossRef] [PubMed]
- Garcia Cruz, D.; Giri, R.R.; Gamiotea Turro, D.; Balsbaugh, J.L.; Adler, A.J.; Rodriguez, A. Lymphocyte Activation Gene-3 Regulates Dendritic Cell Metabolic Programing and T Cell Priming Function. J. Immunol. 2021, 207, 2374–2384. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nono, J.K.; Pletinckx, K.; Lutz, M.B.; Brehm, K. Excretory/secretory-products of Echinococcus multilocularis larvae induce apoptosis and tolerogenic properties in dendritic cells in vitro. PLoS Negl. Trop. Dis. 2012, 6, e1516. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, H.; Li, J.; Guo, B.; Zhao, L.; Zhang, Z.; McManus, D.P.; Wen, H.; Zhang, W. In vitro culture of Echinococcus multilocularis producing protoscoleces and mouse infection with the cultured vesicles. Parasit. Vectors 2016, 9, 411. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, H.; Zhang, C.S.; Fang, B.B.; Hou, J.; Li, W.D.; Li, Z.D.; Li, L.; Bi, X.-J.; Abulizi, A.; Shao, Y.-M.; et al. Dual Role of Hepatic Macrophages in the Establishment of the Echinococcus multilocularis Metacestode in Mice. Front. Immunol. 2020, 11, 600635. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jiang, T.; Sun, W.; Aji, T.; Shao, Y.; Guo, C.; Zhang, C.; Ran, B.; Hou, J.; Yasen, A.; Guo, Q.; et al. Single-Cell Heterogeneity of the Liver-Infiltrating Lymphocytes in Individuals with Chronic Echinococcus multilocularis Infection. Infect. Immun. 2022, 90, e0017722. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nehar-Belaid, D.; Hong, S.; Marches, R.; Chen, G.; Bolisetty, M.; Baisch, J.; Walters, L.; Punaro, M.; Rossi, R.J.; Chung, C.-H.; et al. Mapping systemic lupus erythematosus heterogeneity at the single-cell level. Nat. Immunol. 2020, 21, 1094–1106. [Google Scholar] [CrossRef]
- Zhang, Q.M.; He, Y.; Luo, N.; Patel, S.J.; Han, Y.J.; Gao, R.R.; Modak, M.; Carotta, S.; Haslinger, C.; Kind, D.; et al. Landscape and Dynamics of Single Immune Cells in Hepatocellular Carcinoma. Cell 2019, 179, 829–845.e20. [Google Scholar] [CrossRef]
- Lundie, R.J.; Webb, L.M.; Marley, A.K.; Phythian-Adams, A.T.; Cook, P.C.; Jackson-Jones, L.H.; Brown, S.; Maizels, R.M.; Boon, L.; O’Keeffe, M.; et al. A central role for hepatic conventional dendritic cells in supporting Th2 responses during helminth infection. Immunol. Cell Biol. 2016, 94, 400–410. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jenne, C.N.; Kubes, P. Immune surveillance by the liver. Nat. Immunol. 2013, 14, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Samonigg, H.; Stoger, H.; Auer, H.; Sternthal, M.H.; Wilders-Truschnig, M.; Reisinger, E.C. Use of interferon gamma and mebendazole to stop the progression of alveolar hydatid disease: Case report. Clin. Infect. Dis. 1995, 20, 1543–1546. [Google Scholar] [CrossRef] [PubMed]
- Fresnay, S.; Garnache-Ottou, F.; Plumas, J.; Seilles, E.; Tiberghien, P.; Saas, P. Can tolerogenic dendritic cells help to modulate allo-immune responses in the setting of hematopoietic cell transplantation? Transpl. Immunol. 2003, 11, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Semnani, R.T.; Liu, A.Y.; Sabzevari, H.; Kubofcik, J.; Zhou, J.; Gilden, J.K.; Nutman, T.B. Brugia malayi microfilariae induce cell death in human dendritic cells, inhibit their ability to make IL-12 and IL-10, and reduce their capacity to activate CD4+ T cells. J. Immunol. 2003, 171, 1950–1960. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, R.; Hu, J. Dendritic cell apoptosis: Regulation of tolerance versus immunity. J. Immunol. 2010, 185, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.L.; Terrazas, C.A.; Vera-Arias, L.; Terrazas, L.I. Differential response of antigen presenting cells from susceptible and resistant strains of mice to Taenia crassiceps infection. Infect. Genet. Evol. 2009, 9, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Casaravilla, C.; Pittini, A.; Ruckerl, D.; Seoane, P.I.; Jenkins, S.J.; MacDonald, A.S.; Ferreira, A.M.; Allen, J.E.; Díaz, Á. Unconventional maturation of dendritic cells induced by particles from the laminated layer of larval Echinococcus granulosus. Infect. Immun. 2014, 82, 3164–3176. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, Y.; Wang, Q.; Lv, S.; Zhang, S. Different protein of Echinococcus granulosus stimulates dendritic induced immune response. Parasitology 2015, 142, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Jenne, L.; Arrighi, J.F.; Sauter, B.; Kern, P. Dendritic cells pulsed with unfractionated helminthic proteins to generate antiparasitic cytotoxic T lymphocyte. Parasite Immunol. 2001, 23, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Andreae, S.; Piras, F.; Burdin, N.; Triebel, F. Maturation and activation of dendritic cells induced by lymphocyte activation gene-3 (CD223). J. Immunol. 2002, 168, 3874–3880. [Google Scholar] [CrossRef] [PubMed]
- Voisine, C.; Mastelic, B.; Sponaas, A.M.; Langhorne, J. Classical CD11c+ dendritic cells, not plasmacytoid dendritic cells, induce T cell responses to Plasmodium chabaudi malaria. Int. J. Parasitol. 2010, 40, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Carenza, C.; Calcaterra, F.; Oriolo, F.; Di Vito, C.; Ubezio, M.; Della Porta, M.G.; Mavilio, D.; Della Bella, S. Costimulatory Molecules and Immune Checkpoints Are Differentially Expressed on Different Subsets of Dendritic Cells. Front. Immunol. 2019, 10, 1325. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Teng, C.F.; Wang, T.; Shih, F.Y.; Shyu, W.C.; Jeng, L.B. Therapeutic efficacy of dendritic cell vaccine combined with programmed death 1 inhibitor for hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2021, 36, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Xie, S.; Zhang, Q.; Cao, L.; Niyazi, S.; Lu, X.; Sun, L.; Zhou, Y.; Zhang, Y.; Wang, K. Circulating Th1, Th2, Th17, Treg, and PD-1 Levels in Patients with Brucellosis. J. Immunol. Res. 2019, 2019, 3783209. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, J.; Lozano-Ruiz, B.; Yang, F.M.; Fan, D.D.; Shen, L.; González-Navajas, J.M. The Multifaceted Role of Th1, Th9, and Th17 Cells in Immune Checkpoint Inhibition Therapy. Front. Immunol. 2021, 12, 625667. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, X.; Teng, F.; Kong, L.; Yu, J. PD-L1 expression in human cancers and its association with clinical outcomes. Onco Targets Ther. 2016, 9, 5023–5039. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Luan, Y.Y.; Zhang, L.; Zhu, F.J.; Dong, N.; Lu, J.Y.; Yao, Y.M. Effect of TIPE1 on Immune Function of Dendritic Cells and Its Signaling Pathway in Septic Mice. J. Infect. Dis. 2019, 220, 699–709. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Li, Y.; Yu, Q.; Wang, M.; Ainiwaer, A.; Tang, N.; Zheng, X.; Duolikun, A.; Deng, B.; Li, J.; et al. Immunological Characteristics of Hepatic Dendritic Cells in Patients and Mouse Model with Liver Echinococcus multilocularis Infection. Trop. Med. Infect. Dis. 2024, 9, 95. https://doi.org/10.3390/tropicalmed9050095
Wang H, Li Y, Yu Q, Wang M, Ainiwaer A, Tang N, Zheng X, Duolikun A, Deng B, Li J, et al. Immunological Characteristics of Hepatic Dendritic Cells in Patients and Mouse Model with Liver Echinococcus multilocularis Infection. Tropical Medicine and Infectious Disease. 2024; 9(5):95. https://doi.org/10.3390/tropicalmed9050095
Chicago/Turabian StyleWang, Hui, Yinshi Li, Qian Yu, Mingkun Wang, Abidan Ainiwaer, Na Tang, Xuran Zheng, Adilai Duolikun, Bingqing Deng, Jing Li, and et al. 2024. "Immunological Characteristics of Hepatic Dendritic Cells in Patients and Mouse Model with Liver Echinococcus multilocularis Infection" Tropical Medicine and Infectious Disease 9, no. 5: 95. https://doi.org/10.3390/tropicalmed9050095
APA StyleWang, H., Li, Y., Yu, Q., Wang, M., Ainiwaer, A., Tang, N., Zheng, X., Duolikun, A., Deng, B., Li, J., Shen, Y., & Zhang, C. (2024). Immunological Characteristics of Hepatic Dendritic Cells in Patients and Mouse Model with Liver Echinococcus multilocularis Infection. Tropical Medicine and Infectious Disease, 9(5), 95. https://doi.org/10.3390/tropicalmed9050095