
Stochastic Modeling of Three-Species Prey–Predator Model Driven by Lévy Jump with Mixed Holling-II and Beddington–DeAngelis Functional Responses




Abstract
:1. Introduction
2. The Well-Posedness of the Solution
3. Stochastic Extinction
4. Stochastic Extinction of Predator
5. Stochastic Persistence
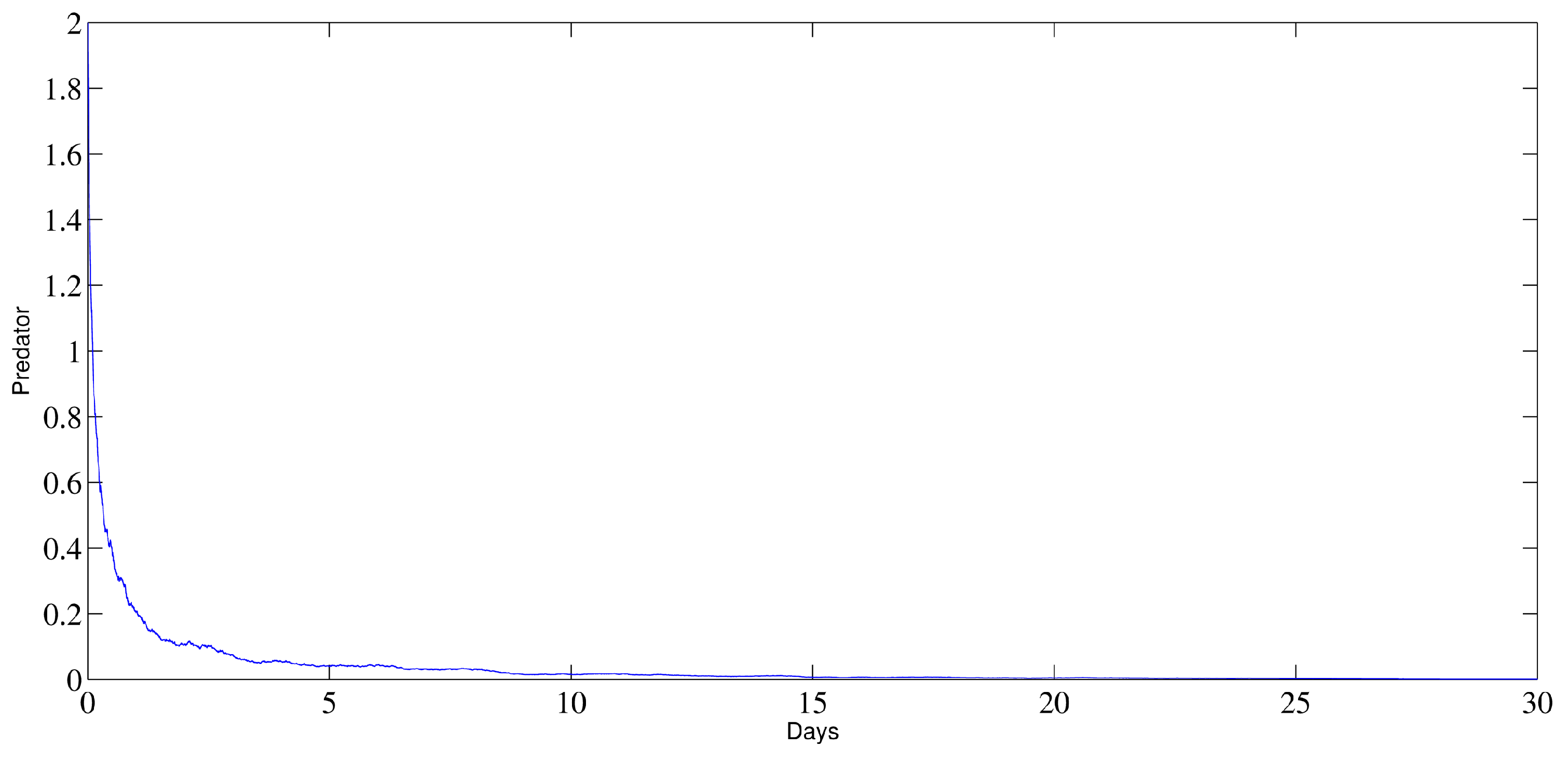
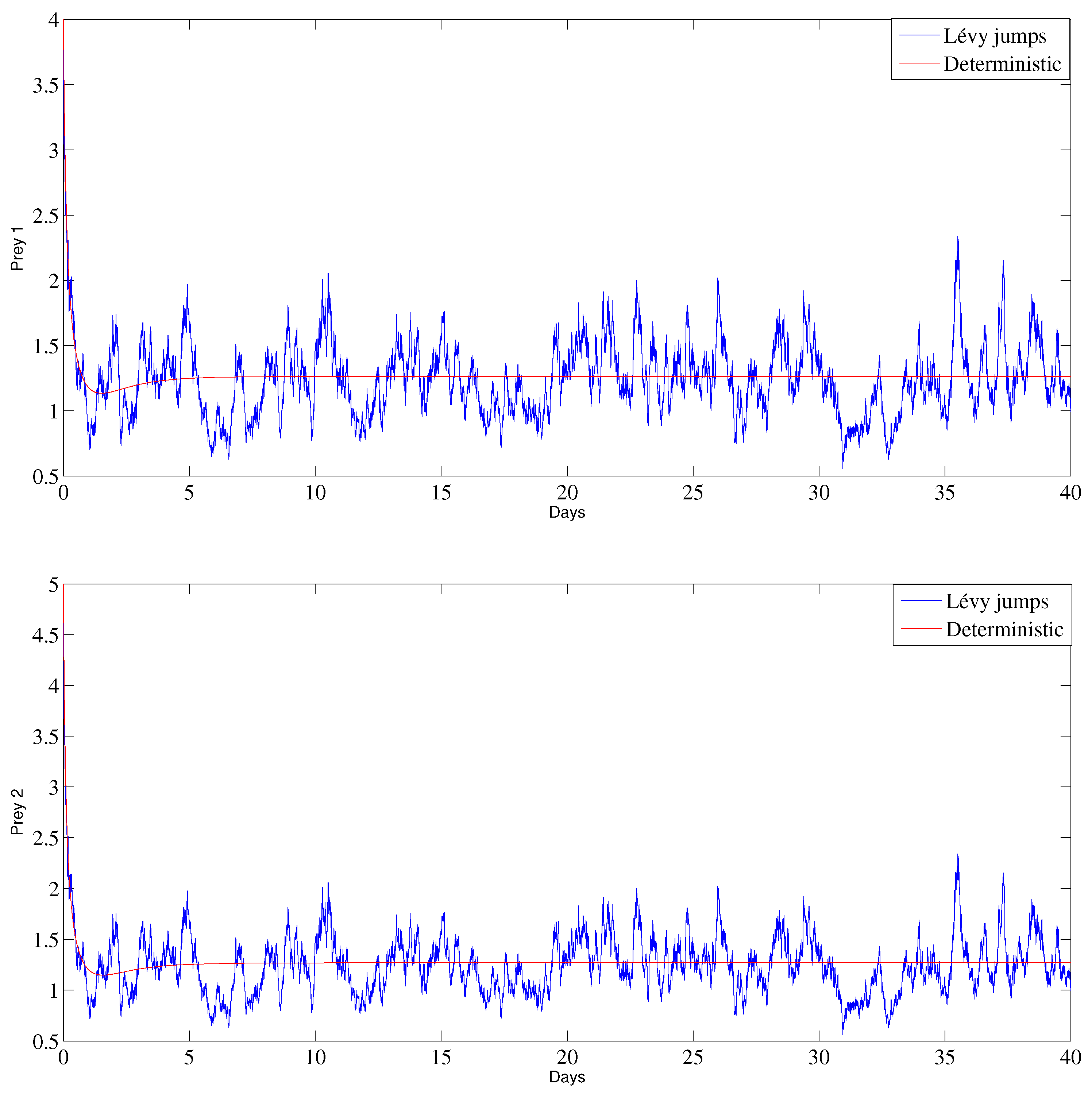
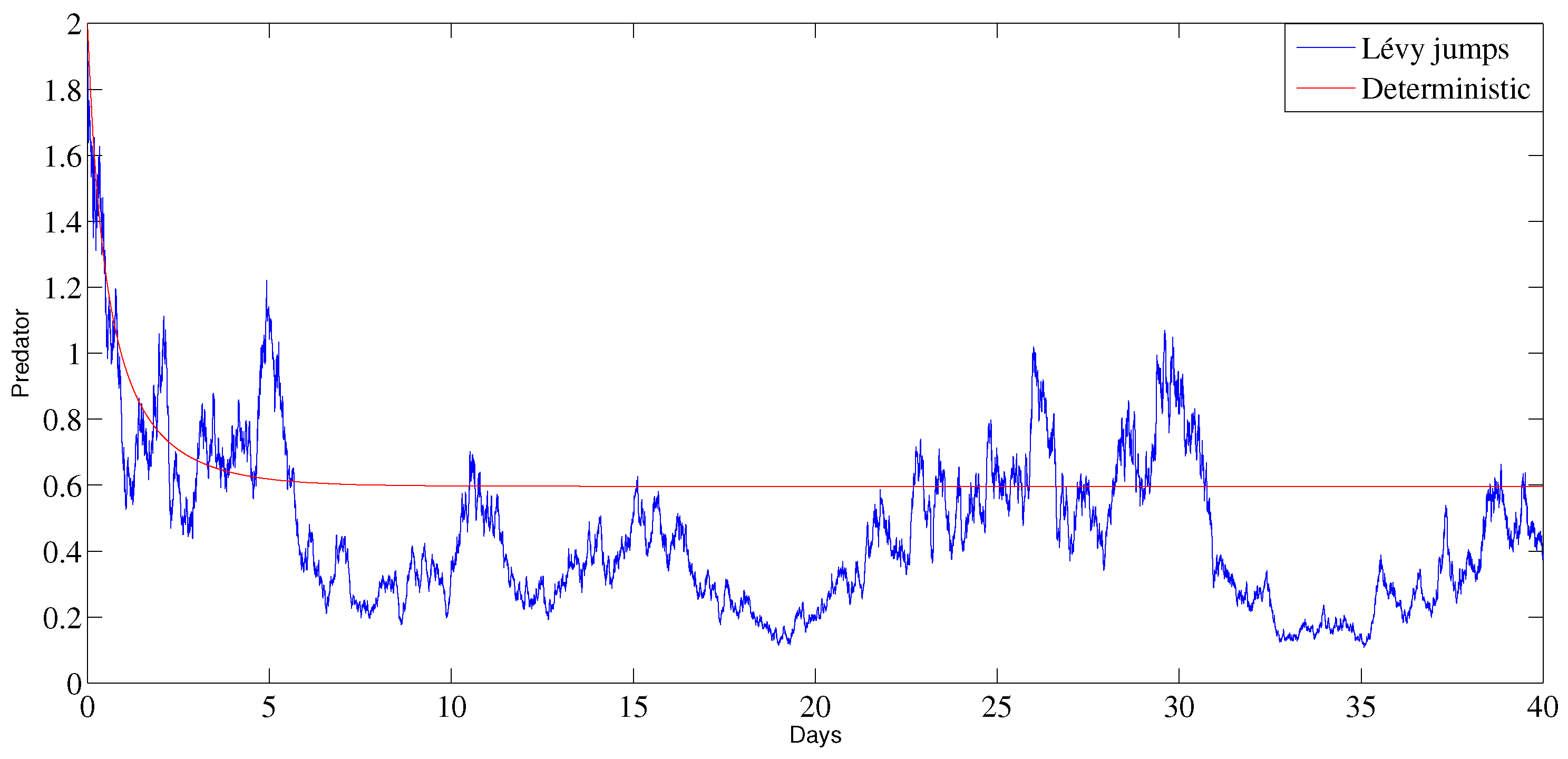
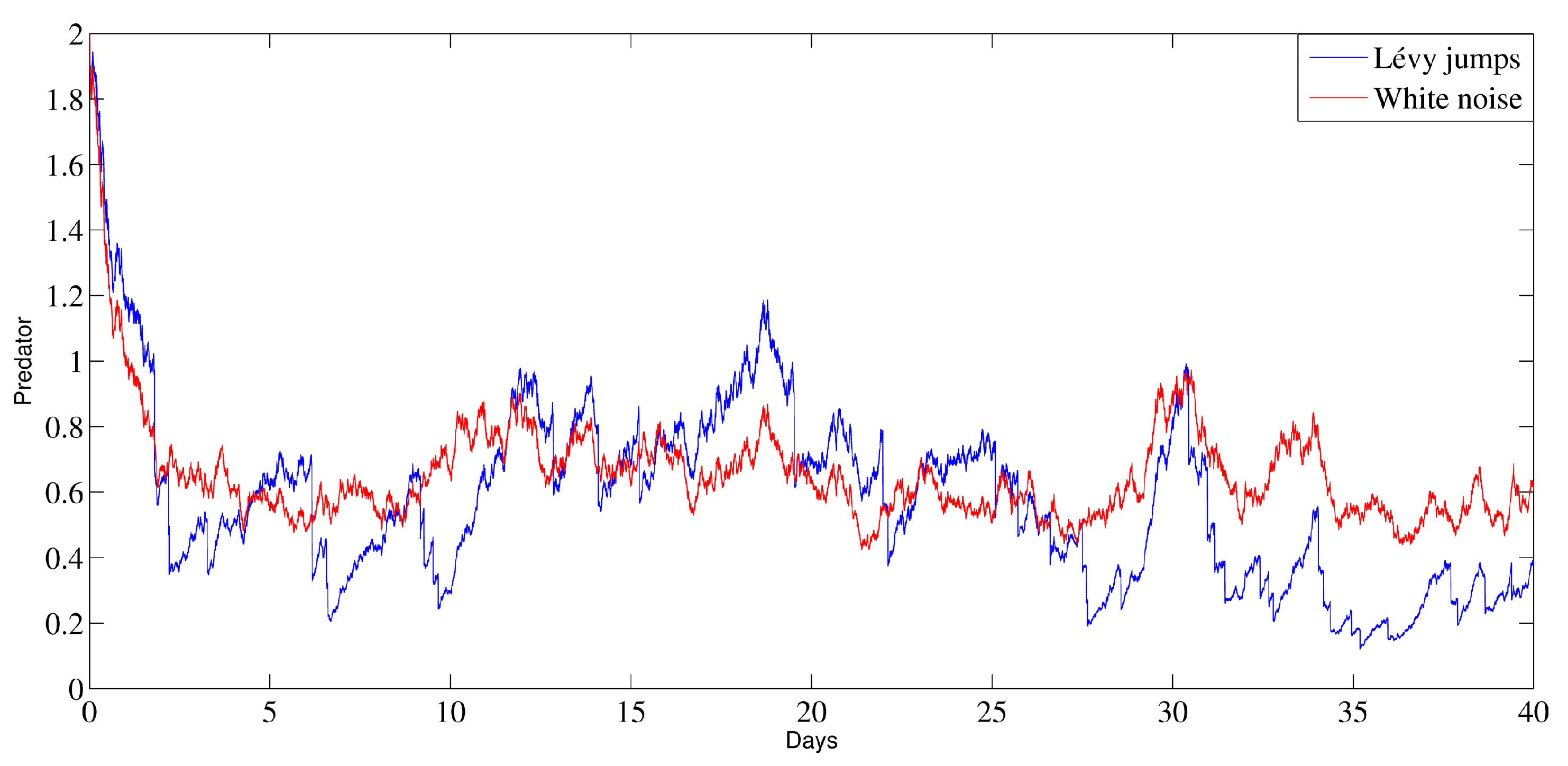
6. Numerical Analysis
7. Conclusions and Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Naik, P.A.; Eskandari, Z.; Shahkari, H.E.; Owolabi, K.M. Bifurcation analysis of a discrete-time prey–predator model. Bull. Biomath. 2023, 1, 111–123. [Google Scholar] [CrossRef]
- Chatterjee, A.; Pal, S. A predator–prey model for the optimal control of fish harvesting through the imposition of a tax. Int. J. Optim. Control Theor. Appl. 2023, 13, 68–80. [Google Scholar] [CrossRef]
- Tripathi, J.P.; Tyagi, S.; Abbas, S. Global analysis of a delayed density dependent predator–prey model with Crowley-Martin functional response. Commun. Nonlinear Sci. Numer. Simul. 2016, 30, 45–69. [Google Scholar] [CrossRef]
- Naik, P.A.; Eskandari, Z.; Yavuz, M.; Zu, J. Complex dynamics of a discrete-time Bazykin–Berezovskaya prey-predator model with a strong Allee effect. J. Comput. Appl. Math. 2022, 413, 114401. [Google Scholar] [CrossRef]
- Lotka, A. Elements of Mathematical Biology; Dover Publications: New York, NY, USA, 1956. [Google Scholar]
- Spencer, P.D.; Collie, J.S. A simple predator–prey model of exploited marine fish populations incorporating alternative prey. ICES J. Mar. Sci. 1996, 53, 615–628. [Google Scholar] [CrossRef]
- Fay, T.H.; Greeff, J.C. Lion, wildebeest and zebra: A predator–prey model. Ecol. Model. 2006, 196, 237–244. [Google Scholar] [CrossRef]
- Verhulst, P.F. Notice sur la loi que la population suit dans son accroissement. Corresp. MathéMatique Phys. 1838, 10, 113–121. [Google Scholar]
- Pearl, R.; Reed, L.J. The logistic curve and the census count of i930. Science 1930, 72, 399–401. [Google Scholar] [CrossRef] [PubMed]
- MacLean, M.; Willard, A. The logistic curve applied to Canada’s population. Can. J. Econ. Polit. Sci. 1937, 3, 241–248. [Google Scholar] [CrossRef]
- Kot, M. Elements of Mathematical Ecology; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Kingsland, S. The refractory model: The logistic curve and the history of population ecology. Q. Rev. Biol. 1982, 57, 29–52. [Google Scholar] [CrossRef]
- Keshet, E.L. Mathematical Models in Biology; McGrawHill: New York, NY, USA, 1988. [Google Scholar]
- Ghosh, D.; Santra, P.K.; Mahapara, G.S. A three-component prey-predator system with interval number. Math. Model. Numer. Simul. Appl. 2023, 3, 1–16. [Google Scholar] [CrossRef]
- Yavuz, M.; Sene, N. Stability analysis and numerical computation of the fractional predator–prey model with the harvesting rate. Fractal Fract. 2020, 4, 35. [Google Scholar] [CrossRef]
- Meng, X.Y.; Huo, H.F.; Xiang, H.; Yin, Q.Y. Stability in a predator–prey model with Crowley-Martin function and stage structure for prey. Appl. Math. Comput. 2014, 232, 810–819. [Google Scholar] [CrossRef]
- Ko, W.; Ryu, K. Qualitative analysis of a predator—Prey model with Holling type II functional response incorporating a prey refuge. J. Differ. Equ. 2006, 231, 534–550. [Google Scholar] [CrossRef]
- Sugie, J.; Kohno, R.; Miyazaki, R. On a predator-prey system of Holling type. Proc. Am. Math. Soc. 1997, 125, 2041–2050. [Google Scholar] [CrossRef]
- Baek, H.; Dongseok, K. Dynamics of a predator-prey system with mixed functional responses. J. Appl. Math. 2014, 2014, 536019. [Google Scholar] [CrossRef]
- Zou, X.; Li, Q.; Cao, W.; Lv, J. Thresholds and critical states for a stochastic predator–prey model with mixed functional responses. Math. Comput. Simul. 2023, 206, 780–795. [Google Scholar] [CrossRef]
- Liu, Q.; Jiang, D.; Hayat, T.; Alsaedi, A. Dynamics of a stochastic predator–prey model with stage structure for predator and Holling type II functional response. J. Nonlinear Sci. 2018, 28, 1151–1187. [Google Scholar] [CrossRef]
- Rihan, F.A.; Alsakaji, H.J. Stochastic delay differential equations of three-species prey-predator system with cooperation among prey species. Discret. Contin. Dyn.-Syst. 2020, 15, 245–263. [Google Scholar] [CrossRef]
- Danane, J.; Allali, K.; Hammouch, Z.; Nisar, K.S. Mathematical analysis and simulation of a stochastic COVID-19 Lévy jump model with isolation strategy. Results Phys. 2021, 23, 103994. [Google Scholar] [CrossRef] [PubMed]
- Akdim, K.; Ez-zetouni, A.; Danane, J.; Allali, K. Stochastic viral infection model with lytic and nonlytic immune responses driven by Lévy noise. Phys. Stat. Mech. Its Appl. 2020, 549, 124367. [Google Scholar] [CrossRef]
- Danane, J. Stochastic predator–prey Lévy jump model with Crowley–Martin functional response and stage structure. J. Appl. Math. Comput. 2021, 67, 41–67. [Google Scholar] [CrossRef]
- Zhao, S.; Song, M. A stochastic predator-prey system with stage structure for predator. Abstr. Appl. Anal. 2014, 2014, 518695. [Google Scholar] [CrossRef]
- Danane, J. Stochastic Capital–Labor Lévy Jump Model with the Precariat Labor Force. Math. Comput. Appl. 2022, 27, 93. [Google Scholar] [CrossRef]
- Choo, S.; Kim, Y.H. Global stability in stochastic difference equations for predator-prey models. J. Comput. Anal. Appl. 2017, 23, 3. [Google Scholar]
- Yang, L.; Zhong, S. Global stability of a stage-structured predator-prey model with stochastic perturbation. Discret. Dyn. Nat. Soc. 2014, 2014, 512817. [Google Scholar] [CrossRef]
- Zhao, Y.; Yuan, S. Stability in distribution of a stochastic hybrid competitive Lotka–Volterra model with Lévy jumps. Chaos Solitons Fractals 2016, 85, 98–109. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, Q.M. Analysis of a stochastic delay predator-prey system with jumps in a polluted environment. Appl. Math. Comput. 2014, 242, 90–100. [Google Scholar] [CrossRef]
- Zhang, X.; Li, W.; Liu, M.; Wang, K. Dynamics of a stochastic Holling II one-predator two-prey system with jumps. Phys. Stat. Mech. Its Appl. 2015, 421, 571–582. [Google Scholar] [CrossRef]
- Liu, M.; Bai, C.; Deng, M.; Du, B. Analysis of stochastic two-prey one-predator model with Lévy jumps. Phys. A Stat. Mech. Its Appl. 2016, 445, 176–188. [Google Scholar] [CrossRef]
- Sabbar, Y. Asymptotic extinction and persistence of a perturbed epidemic model with different intervention measures and standard lévy jumps. Bull. Biomath. 2023, 1, 58–77. [Google Scholar] [CrossRef]
- Mouhcine, N.; Sabbar, Y.; Anwar, Z. Stability characterization of a fractional-order viral system with the non-cytolytic immune assumption. Math. Model. Numer. Simul. Appl. 2022, 2, 164–176. [Google Scholar]
- Wu, J. Stability of a three-species stochastic delay predator–prey system with Lévy noise. Phys. Stat. Mech. Appl. 2018, 502, 492–505. [Google Scholar] [CrossRef]
- Hudson, R.L.; Parthasarathy, K.R. Quantum Ito’s formula and stochastic evolutions. Commun. Math. Phys. 1984, 93, 301–323. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, J.; Wang, W.; Zhang, H. A Predictor–Corrector Compact Difference Scheme for a Nonlinear Fractional Differential Equation. Fractal Fract. 2023, 7, 521. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Description |
|---|---|
| The intrinsic growth rate of prey | |
| The intrinsic growth rate for | |
| The intra-specific competition for prey | |
| The intra-specific competition for prey | |
| The half-saturation due to | |
| The half-saturation due to | |
| The impact of the predator interference | |
| The death rates of predator | |
| The rate of intra-species competition for predator | |
| The average of the transformation of predator from prey | |
| The average of the transformation of predator from prey | |
| d | The natural mortality of the predator y |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danane, J.; Yavuz, M.; Yıldız, M. Stochastic Modeling of Three-Species Prey–Predator Model Driven by Lévy Jump with Mixed Holling-II and Beddington–DeAngelis Functional Responses. Fractal Fract. 2023, 7, 751. https://doi.org/10.3390/fractalfract7100751
Danane J, Yavuz M, Yıldız M. Stochastic Modeling of Three-Species Prey–Predator Model Driven by Lévy Jump with Mixed Holling-II and Beddington–DeAngelis Functional Responses. Fractal and Fractional. 2023; 7(10):751. https://doi.org/10.3390/fractalfract7100751
Chicago/Turabian StyleDanane, Jaouad, Mehmet Yavuz, and Mustafa Yıldız. 2023. "Stochastic Modeling of Three-Species Prey–Predator Model Driven by Lévy Jump with Mixed Holling-II and Beddington–DeAngelis Functional Responses" Fractal and Fractional 7, no. 10: 751. https://doi.org/10.3390/fractalfract7100751
APA StyleDanane, J., Yavuz, M., & Yıldız, M. (2023). Stochastic Modeling of Three-Species Prey–Predator Model Driven by Lévy Jump with Mixed Holling-II and Beddington–DeAngelis Functional Responses. Fractal and Fractional, 7(10), 751. https://doi.org/10.3390/fractalfract7100751