
Occurrence of Human Defensins and S100 Proteins in Head and Neck Basal Cell Carcinoma (BCC) Entities: hBD3 and S100A4 as Potential Biomarkers to Evaluate Successful Surgical Therapy


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Tissue Sampling
2.2. Immunohistology
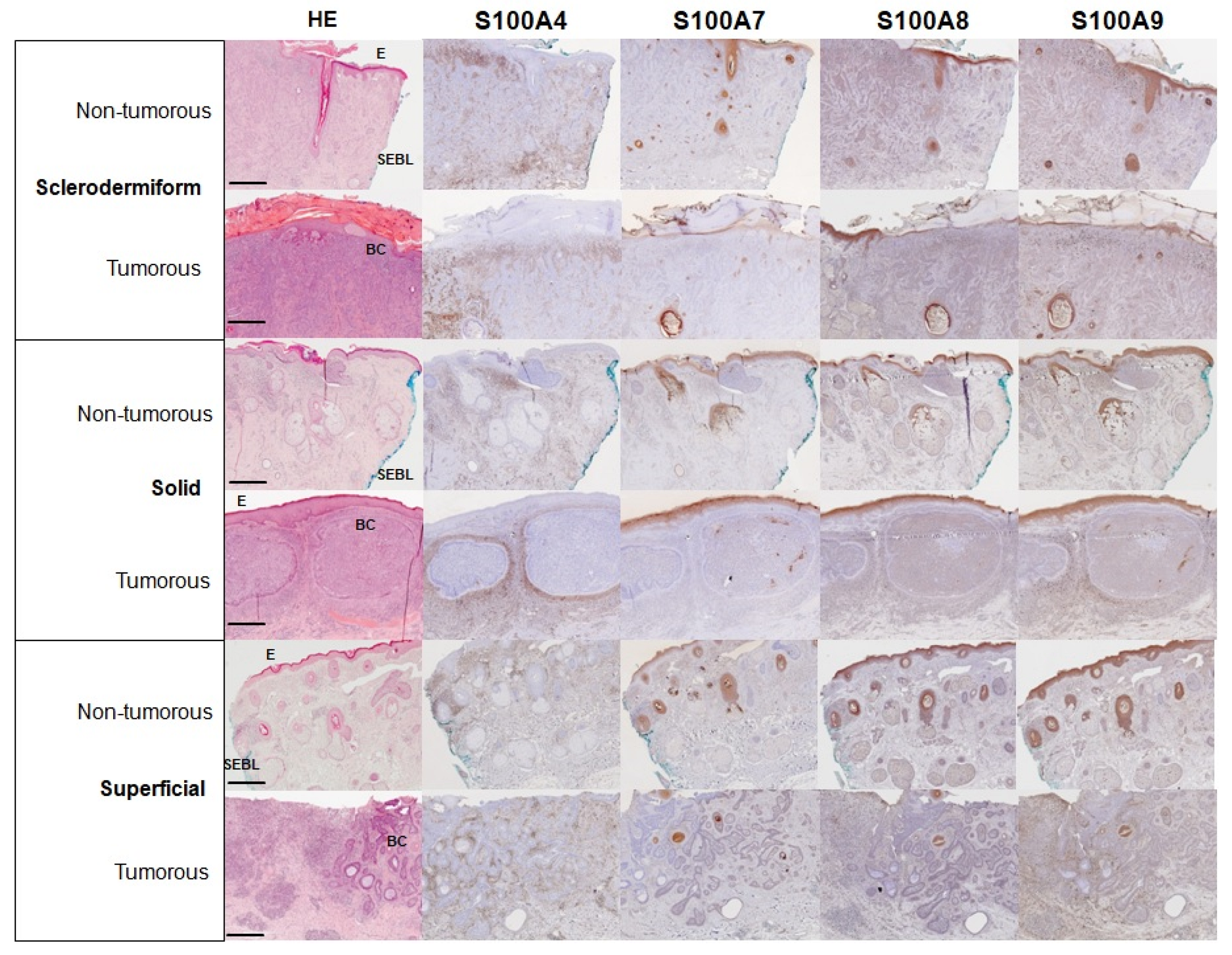
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marzuka, A.G.; Book, S.E. Basal cell carcinoma: Pathogenesis, epidemiology, clinical features, diagnosis, histopathology, and management. Yale J. Biol. Med. 2015, 88, 167–179. [Google Scholar]
- Cancer, Facts & Figures. 2010. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2010.html (accessed on 15 May 2022).
- National Comprehensive Cancer Network. Basal Cell Skin Cancer. Available online: https://www.nccn.org/professionals/physician_gls/default.aspx (accessed on 8 August 2019).
- Rigel, D.S.; Russak, J.; Friedman, R. The evolution of melanoma diagnosis: 25 years beyond the ABCDs. CA Cancer J. Clin. 2010, 60, 301–316. [Google Scholar] [CrossRef]
- Rogers, H.W.; Weinstock, M.A.; Feldman, S.R.; Coldiron, B.M. Incidence Estimate of Nonmelanoma Skin Cancer (Keratinocyte Carcinomas) in the U.S. Population, 2012. JAMA Dermatol. 2015, 151, 1081–1086. [Google Scholar] [CrossRef]
- Scrivener, Y.; Grosshans, E.; Cribier, B. Variations of basal cell carcinomas according to gender, age, location and histopathological subtype. Br. J. Dermatol. 2002, 147, 41–47. [Google Scholar] [CrossRef]
- Soyer, H.P.; Rigel, D.; Wurm, E.M.T. Actinic Keratosis, Basal Cell Carcinoma and Squamous Cell Carcinoma. In Dermatology; Bolognia, J.L., Jorizzo, J.L., Schaffer, J.V., Eds.; Elsevier Saunders: Edinburgh, Scotland, 2012; pp. 1773–1794. [Google Scholar]
- Epstein, E.H. Basal cell carcinomas: Attack of the hedgehog. Nat. Rev. Cancer 2008, 8, 743–754. [Google Scholar] [CrossRef]
- Baxter, J.M.; Patel, A.N.; Varma, S. Facial basal cell carcinoma. BMJ 2012, 345, e5342. [Google Scholar] [CrossRef]
- Dika, E.; Scarfi, F.; Ferracin, M.; Broseghini, E.; Marcelli, E.; Bartolani, B.; Campione, E.; Riefolo, M.; Ricci, C.; Lambertini, M. Basal cell carcinoma: A comprehensive review. Int. J. Mol. Sci. 2020, 21, 5572. [Google Scholar] [CrossRef]
- Sexton, M.; Jones, D.B.; Maloney, M.E. Histologic pattern analysis of basal cell carcinoma: Study of a series of 1039 consecutive neoplasms. J. Am. Acad. Dermatol. 1990, 23, 1118–1126. [Google Scholar] [CrossRef]
- Crowson, N.A. Basal cell carcinoma: Biology, morphology and clinical implications. Mod. Pathol. 2006, 19 (Suppl. S2), S127–S147. [Google Scholar] [CrossRef] [Green Version]
- Geisse, J.; Caro, I.; Lindholm, J.; Golitz, L.; Stampone, P.; Owens, M. Imiquimod 5% cream for the treatment of superficial basal cell carcinoma: Results from two phase III, randomized, vehicle-controlled studies. J. Am. Acad. Dermatol. 2004, 50, 722–733. [Google Scholar] [CrossRef]
- Arits, A.H.H.M.; Mosterd, K.; Essers, B.A.; Spoorenberg, E.; Sommer, A.; De Rooij, M.J.M.; van Pelt, H.P.A.; Quaedvlieg, P.J.F.; Krekels, G.A.M.; van Neer, P.A.F.A.; et al. Photodynamic therapy versus topical imiquimod versus topical fluorouracil for treatment of superficial basal-cell carcinoma: A single blind, non-inferiority, randomised controlled trial. Lancet Oncol. 2013, 14, 647–654. [Google Scholar] [CrossRef]
- Roozeboom, M.H.; Arits, A.H.H.M.; Nelemans, P.J.; Kelleners-Smeets, N.W.J. Overall treatment success after treatment of primary superficial basal cell carcinoma: A systematic review and meta-analysis of randomized and nonrandomized trials. Br. J. Dermatol. 2012, 167, 733–756. [Google Scholar] [CrossRef]
- Silverman, M.K.; Kopf, A.W.; Gladstein, A.H.; Bart, R.S.; Grin, C.M.; Levenstein, M.J. Recurrence rates of treated basal cell carcinomas. Part 4: X-ray therapy. J. Dermatol. Surg. Oncol. 1992, 18, 549–554. [Google Scholar] [CrossRef]
- Kim, D.P.; Kus, K.J.B.; Ruiz, E. Basal cell carcinoma review. Hematol. Oncol. Clin. N. Am. 2019, 33, 13–24. [Google Scholar] [CrossRef]
- Basset-Seguin, N.; Herms, F. Update in the Management of Basal Cell Carcinoma. Acta Derm. Venereol. 2020, 100, adv00140. [Google Scholar] [CrossRef]
- Wenghoefer, M.; Pantelis, A.; Dommisch, H.; Götz, W.; Reich, R.; Bergé, S.; Martini, M.; Allam, J.P.; Jepsen, S.; Merkelbach-Bruse, S.; et al. Nuclear hBD-1 accumulation in malignant salivary gland tumours. BMC Cancer 2008, 8, 290. [Google Scholar] [CrossRef] [Green Version]
- Wenghoefer, M.; Pantelis, A.; Dommisch, H.; Reich, R.; Martini, M.; Allam, J.P.; Novak, N.; Bergé, S.; Jepsen, S.; Winter, J. Decreased gene expression of human beta-defensin-1 in the development of squamous cell carcinoma of the oral cavity. Int. J. Oral Maxillofac. Surg. 2008, 37, 660–663. [Google Scholar] [CrossRef]
- Kesting, M.R.; Loeffelbein, D.J.; Hasler, R.J.; Wolff, K.D.; Rittig, A.; Schulte, M.; Hirsch, T.; Wagenpfeil, S.; Jacobsen, F.; Steinstraesser, L. Expression profile of human beta-defensin 3 in oral squamous cell carcinoma. Cancer Investig. 2009, 27, 575–581. [Google Scholar] [CrossRef]
- Kesting, M.R.; Sudhoff, H.; Hasler, R.J.; Nieberler, M.; Pautke, C.; Wolff, K.D.; Wagenpfeil, S.; Al-Benna, S.; Jacobsen, F.; Steinstraesser, L. Psoriasin (S100A7) up-regulation in oral squamous cell carcinoma and its relation to clinicopathologic features. Oral Oncol. 2009, 45, 731–736. [Google Scholar] [CrossRef]
- Pantelis, A.; Wenghoefer, M.; Haas, S.; Merkelbach-Bruse, S.; Pantelis, D.; Jepsen, S.; Bootz, F.; Winter, J. Down regulation and nuclear localization of human beta-defensin-1 in pleomorphic adenomas of salivary glands. Oral Oncol. 2009, 45, 526–530. [Google Scholar] [CrossRef]
- Winter, J.; Pantelis, A.; Reich, R.; Jepsen, S.; Allam, J.P.; Novak, N.; Wenghoefer, M. Risk estimation for a malignant transformation of oral lesions by S100A7 and Doc-1 gene expression. Cancer Investig. 2011, 29, 478–484. [Google Scholar]
- Winter, J.; Pantelis, A.; Reich, R.; Martini, M.; Kraus, D.; Jepsen, S.; Allam, J.P.; Novak, N.; Wenghoefer, M. Human beta-defensin-1, -2, and -3 exhibit opposite effects on oral squamous cell carcinoma cell proliferation. Cancer Investig. 2011, 29, 196–201. [Google Scholar] [CrossRef]
- Winter, J.; Pantelis, A.; Kraus, D.; Reckenbeil, J.; Reich, R.; Jepsen, S.; Fischer, H.P.; Allam, J.P.; Novak, N.; Wenghoefer, M. Human α-defensin (DEFA) gene expression helps to characterise benign and malignant salivary gland tumours. BMC Cancer 2012, 12, 465. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.; Mohr, S.; Pantelis, A.; Kraus, D.; Allam, J.P.; Novak, N.; Reich, R.; Martini, M.; Jepsen, S.; Götz, W.; et al. IGF-1 deficiency in combination with a low basic hBD-2 and hBD-3 gene expression might counteract malignant transformation in pleomorphic adenomas in vitro. Cancer Investig. 2012, 30, 106–113. [Google Scholar] [CrossRef]
- Winter, J.; Wenghoefer, M. Human Defensins: Potential Tools for Clinical Applications. Polymers 2012, 4, 691–709. [Google Scholar] [CrossRef]
- Kesting, M.R.; Stoeckelhuber, M.; Kuppek, A.; Hasler, R.; Rohleder, N.; Wolff, K.D.; Nieberler, M. Human β-defensins and psoriasin/S100A7 expression in salivary glands: Anti-oncogenic molecules for potential therapeutic approaches. BioDrugs 2012, 26, 33–42. [Google Scholar] [CrossRef]
- Reckenbeil, J.; Kraus, D.; Probstmeier, R.; Allam, J.P.; Novak, N.; Frentzen, M.; Martini, M.; Wenghoefer, M.; Winter, J. Cellular Distribution and Gene Expression Pattern of Metastasin (S100A4), Calgranulin A (S100A8), and Calgranulin B (S100A9) in Oral Lesions as Markers for Molecular Pathology. Cancer Investig. 2016, 34, 246–254. [Google Scholar] [CrossRef]
- Probstmeier, R.; Kraus, D.; Wenghoefer, M.; Winter, J. S100 Proteins as Biomarkers in Risk Estimations for Malignant Transformation in Oral Lesions. Methods Mol. Biol. 2019, 1929, 763–771. [Google Scholar]
- Gambichler, T.; Skrygan, M.; Huyn, J.; Bechara, F.G.; Sand, M.; Altmeyer, P.; Kreuter, A. Pattern of mRNA expression of beta-defensins in basal cell carcinoma. BMC Cancer 2006, 6, 163. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Lehrer, R.I. Primate defensins. Nat. Rev. Microbiol. 2004, 2, 727–738. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Hazlett, L.; Wu, M. Defensins in innate immunity. Cell Tissue Res. 2011, 343, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Donald, C.D.; Sun, C.Q.; Lim, S.D.; Macoska, J.; Cohen, C.; Amin, M.B.; Young, A.N.; Ganz, T.A.; Marshall, F.F.; Petros, J.A. Cancer-specific loss of beta-defensin 1 in renal and prostatic carcinomas. Lab. Investig. 2003, 83, 501–505. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.Q.; Arnold, R.; Fernandez-Golarz, C.; Parrish, A.B.; Almekinder, T.; He, J.; Ho, S.M.; Svoboda, P.; Pohl, J.; Marshall, F.F.; et al. Human beta-defensin-1, a potential chromosome 8p tumor suppressor: Control of transcription and induction of apoptosis in renal cell carcinoma. Cancer Res. 2006, 66, 8542–8549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial peptides human beta-defensins stimulate epidermal keratinocyte migration, proliferation and production of proinflammatory cytokines and chemokines. J. Investig. Dermatol. 2007, 127, 594–604. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, T.; Kraus, D.; Novak, N.; Probstmeier, R.; Frentzen, M.; Wenghoefer, M.; Jepsen, S.; Winter, J. Oral pathogens change proliferation properties of oral tumor cells by affecting gene expression of human defensins. Tumour Biol. 2016, 37, 13789–13798. [Google Scholar] [CrossRef]
- Donato, R. Functional roles of S100 proteins, calcium-binding proteins of the EF-hand type. Biochim. Biophys. Acta 1999, 1450, 191–231. [Google Scholar] [CrossRef] [Green Version]
- Donato, R. S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int. J. Biochem. Cell Biol. 2001, 33, 637–668. [Google Scholar] [CrossRef]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xu, C.; Jin, Q.; Liu, Z. S100 protein family in human cancer. Am. J. Cancer Res. 2014, 4, 89–115. [Google Scholar]
- Halawi, A.; Abbas, O.; Mahalingam, M. S100 proteins and the skin: A review. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 405–414. [Google Scholar] [CrossRef]
- Bresnick, A.R.; Weber, D.J.; Zimmer, D.B. S100 proteins in cancer. Nat. Rev. Cancer 2015, 15, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Niyonsaba, F.; Kiatsurayanon, C.; Chieosilapatham, P.; Ogawa, H. Friends or Foes? Host defense (antimicrobial) peptides and proteins in human skin diseases. Exp. Dermatol. 2017, 26, 989–998. [Google Scholar] [CrossRef] [Green Version]
- Heizmann, C.W. S100 proteins: Diagnostic and prognostic biomarkers in laboratory medicine. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1197–1206. [Google Scholar] [CrossRef]
- Wenghoefer, M.; Pantelis, A.; Najafi, T.; Deschner, J.; Allam, J.P.; Novak, N.; Reich, R.; Martini, M.; Bergé, S.; Fischer, H.P.; et al. Gene expression of oncogenes, antimicrobial peptides, and cytokines in the development of oral leucoplakia. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 110, 351–356. [Google Scholar] [CrossRef]
- Winter, J.; Pantelis, A.; Allam, J.P.; Novak, N.; Reich, R.; Martini, M.; Bergé, S.; Deschner, J.; Jepsen, S.; Wenghoefer, M. High α-defensin and S100A7 expression and missing DOC-1 down-regulation characterize irritation fibromas of the oral cavity and may counteract malignant transformation. J. Craniofac. Surg. 2011, 22, 100–104. [Google Scholar] [CrossRef]
- Gläser, R.; Harder, J.; Lange, H.; Bartels, J.; Christophers, E.; Schröder, J.M. Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat. Immunol. 2005, 6, 57–64. [Google Scholar] [CrossRef]
- Pirr, S.; Richter, M.; Fehlhaber, B.; Pagel, J.; Härtel, C.; Roth, J.; Vogl, T.; Viemann, D. High amounts of S100-alarmins confer antimicrobial activity on human breast milk targeting pathogens relevant in neonatal sepsis. Front. Immunol. 2017, 8, 1822. [Google Scholar] [CrossRef] [Green Version]
- Vrekoussis, T.; Chaniotis, V.; Navrozoglou, I.; Dousias, V.; Pavlakis, K.; Stathopoulos, E.N.; Zoras, O. Image analysis of breast cancer immunohistochemistry-stained sections using ImageJ: An RGB-based model. Anticancer Res. 2009, 29, 4995–4998. [Google Scholar]
- Cizkova, K.; Foltynkova, T.; Gachechiladze, M.; Tauber, Z. Comparative Analysis of Immunohistochemical Staining Intensity Determined by Light Microscopy, ImageJ and QuPath in Placental Hofbauer Cells. Acta Histochem. Cytochem. 2021, 54, 21–29. [Google Scholar] [CrossRef]
- Ishibashi, H.; Suzuki, T.; Suzuki, S.; Moriya, T.; Kaneko, C.; Takizawa, T.; Sunamori, M.; Handa, M.; Kondo, T.; Sasano, H. Sex steroid hormone receptors in human thymoma. J. Clin. Endocrinol. Metab. 2003, 88, 2309–2317. [Google Scholar] [CrossRef]
- Hoppe, T.; Kraus, D.; Probstmeier, R.; Jepsen, S.; Winter, J. Stimulation with Porphyromonas gingivalis enhances malignancy and initiates anoikis resistance in immortalized oral keratinocytes. J. Cell Physiol. 2019, 234, 21903–21914. [Google Scholar] [CrossRef]
- Kiatsurayanon, C.; Peng, G.; Niyonsaba, F. Opposing Roles of Antimicrobial Peptides in Skin Cancers. Curr. Pharm. Des. 2022, 28, 248–258. [Google Scholar] [CrossRef]
- Scola, N.; Gambichler, T.; Saklaoui, H.; Bechara, F.G.; Georgas, D.; Stücker, M.; Gläser, R.; Kreuter, A. The expression of antimicrobial peptides is significantly altered in cutaneous squamous cell carcinoma and precursor lesions. Br. J. Dermatol. 2012, 167, 591–597. [Google Scholar] [CrossRef]
- Kamino, Y.; Kurashige, Y.; Uehara, O.; Sato, J.; Nishimura, M.; Yoshida, K.; Arakawa, T.; Nagayasu, H.; Saitoh, M.; Abiko, Y. HBD-2 is downregulated in oral carcinoma cells by DNA hypermethylation, and increased expression of hBD-2 by DNA demethylation and gene transfection inhibits cell proliferation and invasion. Oncol. Rep. 2014, 32, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Kawsar, H.I.; Weinberg, A.; Hirsch, S.A.; Venizelos, A.; Howell, S.; Jiang, B.; Jin, G. Overexpression of human beta-defensin-3 in oral dysplasia: Potential role in macrophage trafficking. Oral Oncol. 2009, 45, 696–702. [Google Scholar] [CrossRef]
- Bick, R.J.; Poindexter, B.J.; Buja, L.M.; Lawyer, C.H.; Milner, S.M.; Bhat, S. Nuclear localization of HBD-1 in human keratino-cytes. J. Burns Wounds 2007, 7, e3. [Google Scholar]
- Böni, R.; Burg, G.; Doguoglu, A.; Ilg, E.C.; Schäfer, B.W.; Müller, B.; Heizmann, C.W. Immunohistochemical localization of the Ca2+ binding S100 proteins in normal human skin and melanocytic lesions. Br. J. Dermatol. 1997, 137, 39–43. [Google Scholar] [CrossRef]
- Zhu, L.; Kohda, F.; Nakahara, T.; Chiba, T.; Tsuji, G.; Hachisuka, J.; Ito, T.; Tu, Y.; Moroi, Y.; Uchi, H.; et al. Aberrant expression of S100A6 and matrix metalloproteinase 9, but not S100A2, S100A4, and S100A7, is associated with epidermal carcinogenesis. J. Dermatol. Sci. 2013, 72, 311–319. [Google Scholar] [CrossRef]
- Watson, P.H.; Leygue, E.R.; Murphy, L.C. Psoriasin (S100A7). Int. J. Biochem. Cell Biol. 1998, 30, 567–571. [Google Scholar] [CrossRef]
- Gebhardt, C.; Riehl, A.; Durchdewald, M.; Németh, J.; Fürstenberger, G.; Müller-Decker, K.; Enk, A.; Arnold, B.; Bierhaus, A.; Nawroth, P.P.; et al. RAGE signaling sustains inflammation and promotes tumor development. J. Exp. Med. 2008, 205, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Hattinger, E.; Zwicker, S.; Ruzicka, T.; Yuspa, S.H.; Wolf, R. Opposing functions of psoriasin (S100A7) and koebnerisin (S100A15) in epithelial carcinogenesis. Curr. Opin. Pharmacol. 2013, 13, 588–594. [Google Scholar] [CrossRef]
- Alowami, S.; Qing, G.; Emberley, E.; Snell, L.; Watson, P.H. Psoriasin (S100A7) expression is altered during skin tumorigenesis. BMC Dermatol. 2003, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Moubayed, N.; Weichenthal, M.; Harder, J.; Wandel, E.; Sticherling, M.; Gläser, R. Psoriasin (S100A7) is significantly up-regulated in human epithelial skin tumours. J. Cancer Res. Clin. Oncol. 2007, 133, 253–261. [Google Scholar] [CrossRef]
- Martinsson, H.; Yhr, M.; Enerbäck, C. Expression patterns of S100A7 (psoriasin) and S100A9 (calgranulin-B) in keratinocyte differentiation. Exp. Dermatol. 2005, 14, 161–168. [Google Scholar] [CrossRef]
- Lukanidin, E.; Sleeman, J.P. Building the niche: The role of the S100 proteins in metastatic growth. Semin. Cancer Biol. 2012, 22, 216–225. [Google Scholar] [CrossRef]
- Van Crombruggen, K.; Vogl, T.; Pérez-Novo, C.; Holtappels, G.; Bachert, C. Differential release and desposition of S100A8/A9 proteins in inflamed upper airway tissue. Eur. Respir. J. 2016, 47, 264–274. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age | BCC Type |
|---|---|---|---|
| 1 | m | 76 | sclerodermiform |
| 2 | m | 84 | sclerodermiform |
| 3 | m | 82 | sclerodermiform |
| 4 | f | 77 | sclerodermiform |
| 5 | f | 81 | sclerodermiform |
| 6 | m | 66 | sclerodermiform |
| 7 | f | 81 | sclerodermiform |
| 8 | m | 69 | sclerodermiform |
| 9 | f | 72 | sclerodermiform |
| 10 | f | 75 | sclerodermiform |
| 11 | m | 76 | solid |
| 12 | f | 84 | solid |
| 13 | m | 81 | solid |
| 14 | f | 89 | solid |
| 15 | f | 62 | solid |
| 16 | m | 73 | solid |
| 17 | f | 71 | solid |
| 18 | m | 71 | solid |
| 19 | f | 53 | solid |
| 20 | m | 65 | solid |
| 21 | f | 85 | superficial |
| 22 | m | 84 | superficial |
| 23 | m | 82 | superficial |
| 24 | f | 71 | superficial |
| 25 | f | 80 | superficial |
| 26 | m | 73 | superficial |
| 27 | f | 72 | superficial |
| 28 | m | 71 | superficial |
| 29 | f | 84 | superficial |
| 30 | m | 50 | superficial |
| Sclerodermiform | Solid | Superficial | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Non-Tumorous | Tumor | Non-Tumorous | Tumor | Non-Tumorous | Tumor | |||||||
| Epith. | Stroma | Center | Edge | Epith. | Stroma | Center | Edge | Epith. | Stroma | Center | Edge | |
| hBD1 | 98 (4) | 49 (5) | 99 (3) | 100 | 100 | 18 (6) | 98 (4) | 97 (6) | 100 | 22 (4) | 11 (3) | 99 (4) |
| hBD2 | 100 | 71 (5) | 53 (6) | 100 | 99 (3) | 31 (5) | 100 | 100 | 100 | 31 (3) | 22 (4) | 97 (6) |
| hBD3 | 100 | 99 (3) | 97 (6) | 100 n/52 (4) | 100 | 12 (4) | 97 (6) | 99 (3) | 100 | 98 (4) | 23 (8) | 97 (3) n/25 (5) |
| DEFA1/3 | 97 (6) | 48 (6) | 99 (3) | 100 | 98 (4) | 49 (7) | 100 | 79 (5) | 100 | 99 (3) | 12 (4) | 97 (6) |
| DEFA4 | 100 | 11 (3) | 100 | 98 (4) | 99 (3) | 13 (4) | 100 | 97 (6) | 100 | 12 (4) | 12 (4) | 98 (4) |
| S100A4 | 0 | 68 (6) | 0 | 0 | 0 | 53 (6) | 0 | 0 | 0 | 51 (6) n/48 (4) | 0 | 0 |
| S100A7 | 81 (5) | 0 | 0 | 0 | 89 (5) | 0 | 0 | 0 | 82 (4) | 0 | 0 | 0 |
| S100A8 | 88 (4) | 0 | 99 (3) | 100 | 90 (6) | 0 | 8 (6) | 100 | 81 (5) | 0 | 78 (6) | 99 (3) |
| S100A9 | 100 | 0 | 51 (5) | 99 (3) | 98 (4) | 0 | 99 (3) | 100 | 97 (6) | 0 | 0 | 0 |
| Sclerodermiform | Solid | Superficial | ||||
|---|---|---|---|---|---|---|
| Non-Tumorous | Tumor | Non-Tumorous | Tumor | Non-Tumorous | Tumor | |
| hBD1 | E/++/196S/++/98 | TC/++/198TE/++/200 | E/++/200S/++/36 | TC/++/196TE/++/194 | E/++/200S/++/44 | TC/+/11TE/++/198 |
| hBD2 | E/+++/300S/++/142 | TC/+/53TE/++/200 | E/+++/297S/+/31 | TC/++/200TE/++/200 | E/+++/300S/+/31 | TC/+/20TE/++/194 |
| hBD3 | E/++/200S/+/99 | TC/+/97TE/+/c/100TE/++/n/104 | E/++/200S/+/12 | TC/++/194TE/++/198 | E/++/200S/++/196 | TC/++/46TC/++/c/198TE/++/n/50 |
| DEFA1/3 | E/++/194S/+/48 | TC/++/198TE/++/200 | E/++/196S/+/49 | TC/++/200TE/++/158 | E/++/200S/+/198 | TC/++/24TE/++/194 |
| DEFA4 | E/++/200S/+/11 | TC/++/200TE/++/196 | E/+/99S/+/13 | TC/++/200TE/++/194 | E/++/200S/+/12 | TC/+/11TE/++/196 |
| S100A4 | E/-/0S/+/68 | TC/-/0TE/-/0 | E/-/0S/+/53 | TC/-/0TE/-/0 | E/-/0S/+/c/51S/+/n/48 | TC/-/0TE/-/0 |
| S100A7 | E/+/81S/-/0 | TC/-/0TE/-/0 | E/++/178S/-/0 | TC/-/0TE/-/0 | E/++/164S/-/0 | TC/-/0TE/-/0 |
| S100A8 | E/++/176S/-/0 | TC/+/198TE/+/100 | E/++/180S/-/0 | TC/+/88TE/+/100 | E/+++/243S/-/0 | TC/+/78TE/+/99 |
| S100A9 | E/+++/300S/-/0 | TC/+/51TE/+/99 | E/++/196S/-/0 | TC/+/99TE/+/100 | E/+++/291S/-/0 | TC/-/0TE/-/0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dröge, E.; Probstmeier, R.; Wenghoefer, M.; Winter, J. Occurrence of Human Defensins and S100 Proteins in Head and Neck Basal Cell Carcinoma (BCC) Entities: hBD3 and S100A4 as Potential Biomarkers to Evaluate Successful Surgical Therapy. J. Otorhinolaryngol. Hear. Balance Med. 2023, 4, 1. https://doi.org/10.3390/ohbm4010001
Dröge E, Probstmeier R, Wenghoefer M, Winter J. Occurrence of Human Defensins and S100 Proteins in Head and Neck Basal Cell Carcinoma (BCC) Entities: hBD3 and S100A4 as Potential Biomarkers to Evaluate Successful Surgical Therapy. Journal of Otorhinolaryngology, Hearing and Balance Medicine. 2023; 4(1):1. https://doi.org/10.3390/ohbm4010001
Chicago/Turabian StyleDröge, Eva, Rainer Probstmeier, Matthias Wenghoefer, and Jochen Winter. 2023. "Occurrence of Human Defensins and S100 Proteins in Head and Neck Basal Cell Carcinoma (BCC) Entities: hBD3 and S100A4 as Potential Biomarkers to Evaluate Successful Surgical Therapy" Journal of Otorhinolaryngology, Hearing and Balance Medicine 4, no. 1: 1. https://doi.org/10.3390/ohbm4010001
APA StyleDröge, E., Probstmeier, R., Wenghoefer, M., & Winter, J. (2023). Occurrence of Human Defensins and S100 Proteins in Head and Neck Basal Cell Carcinoma (BCC) Entities: hBD3 and S100A4 as Potential Biomarkers to Evaluate Successful Surgical Therapy. Journal of Otorhinolaryngology, Hearing and Balance Medicine, 4(1), 1. https://doi.org/10.3390/ohbm4010001