Ammonium Cycling and Nitrification Stimulation during Oil Sludge Remediation by Gram-Positive Bacteria Lysinibacillus sphaericus Using Red Wiggler Earthworm Eisenia fetida


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Summary
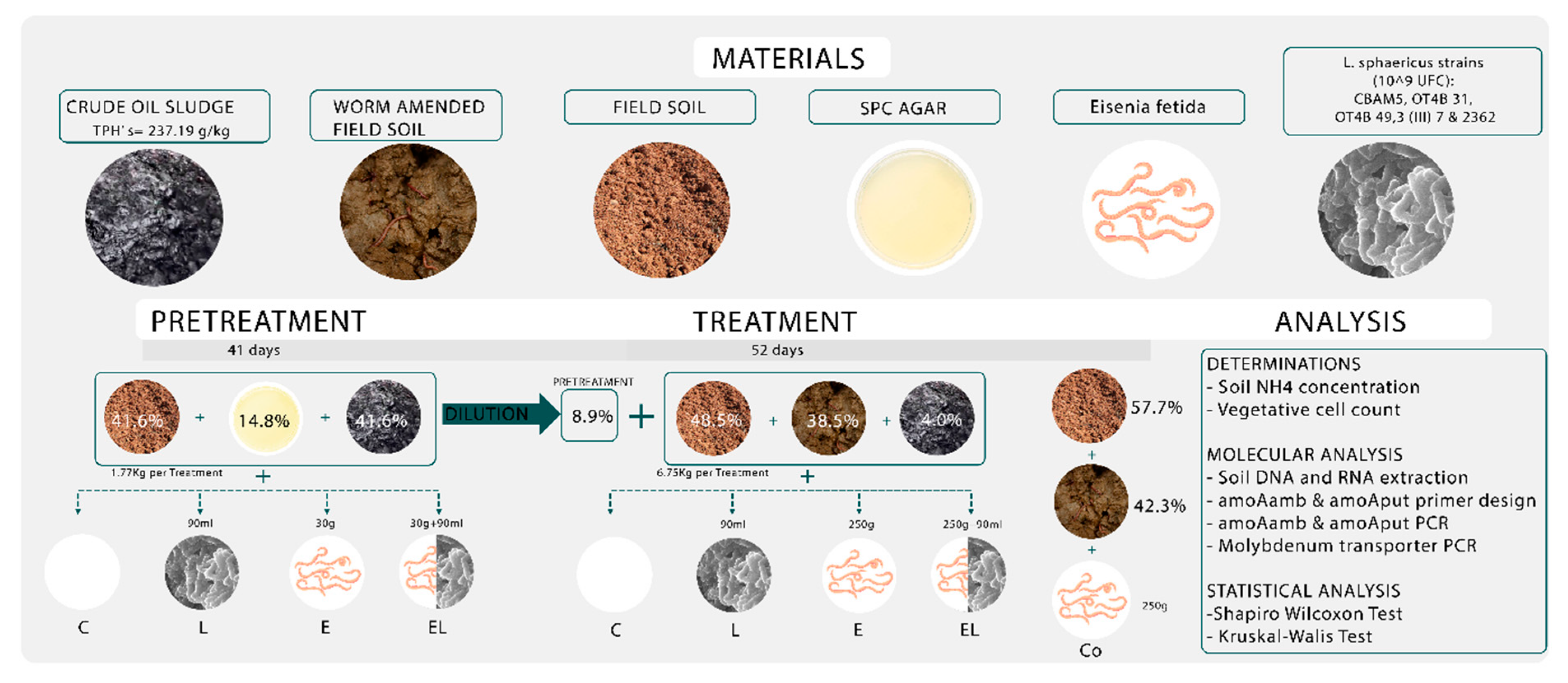
2.2. Materials
2.3. Composted Soil, Worm Castings and Bacteria Growth
2.4. Pretreatment
2.5. Treatment
2.6. Sampling and Analysis
2.7. Ammonium and Hydrocarbon Determination and Microbial Count
2.8. DNA/RNA Extraction and PCR
3. Results and Discussion
3.1. Environmental Conditions and TPH’s Concentration
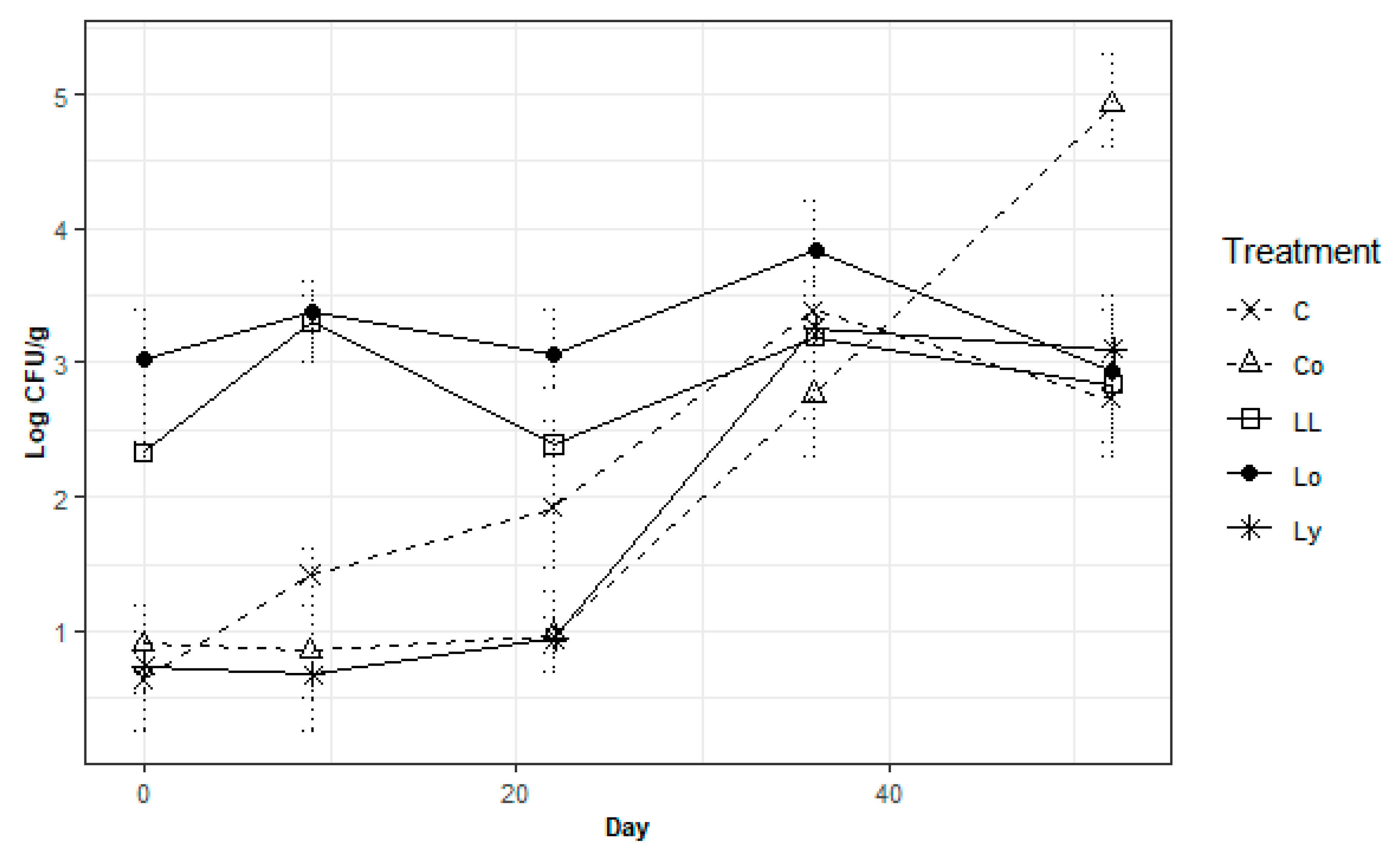
3.2. Bacterial Growth
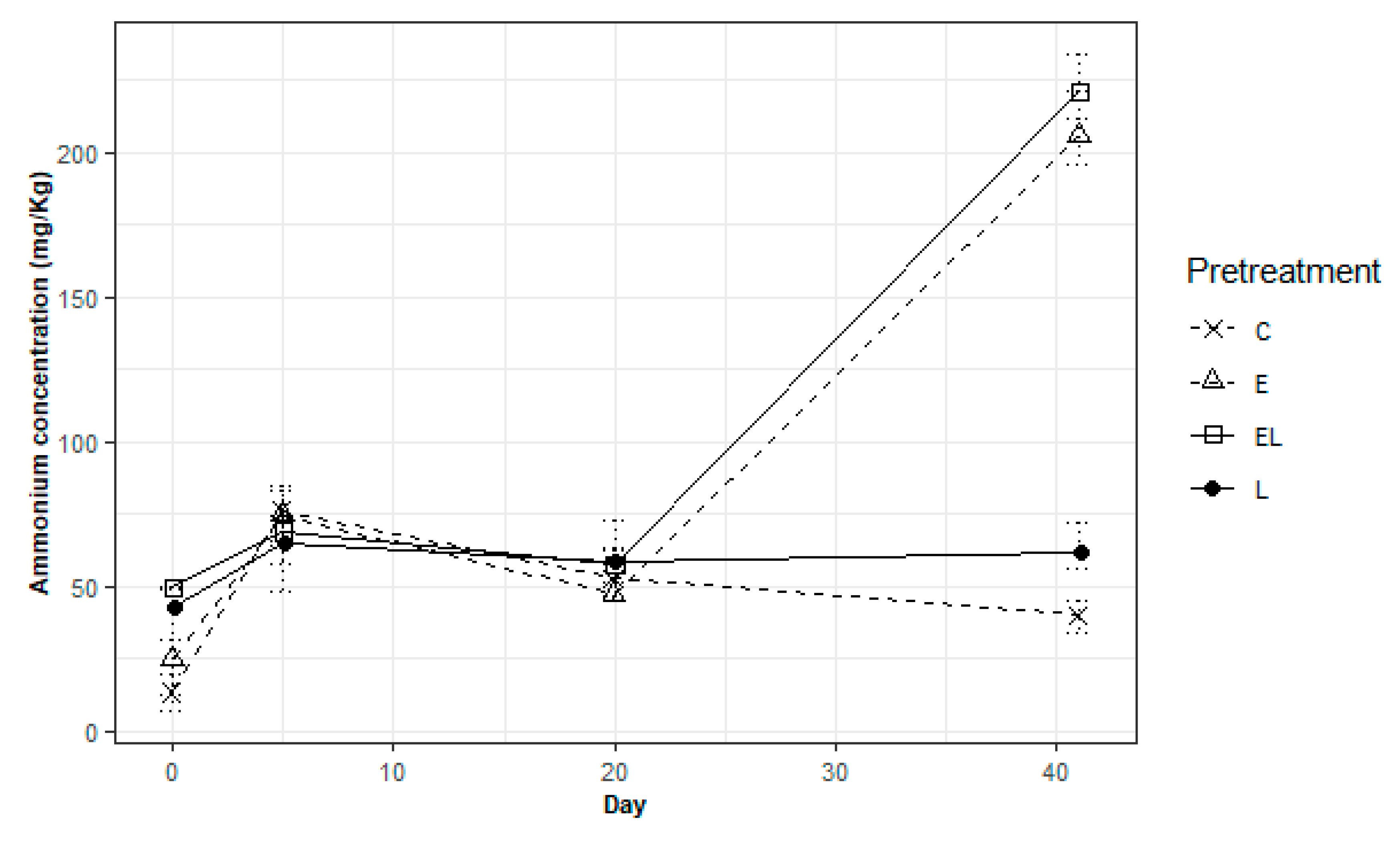
3.3. Ammonium Concentrations
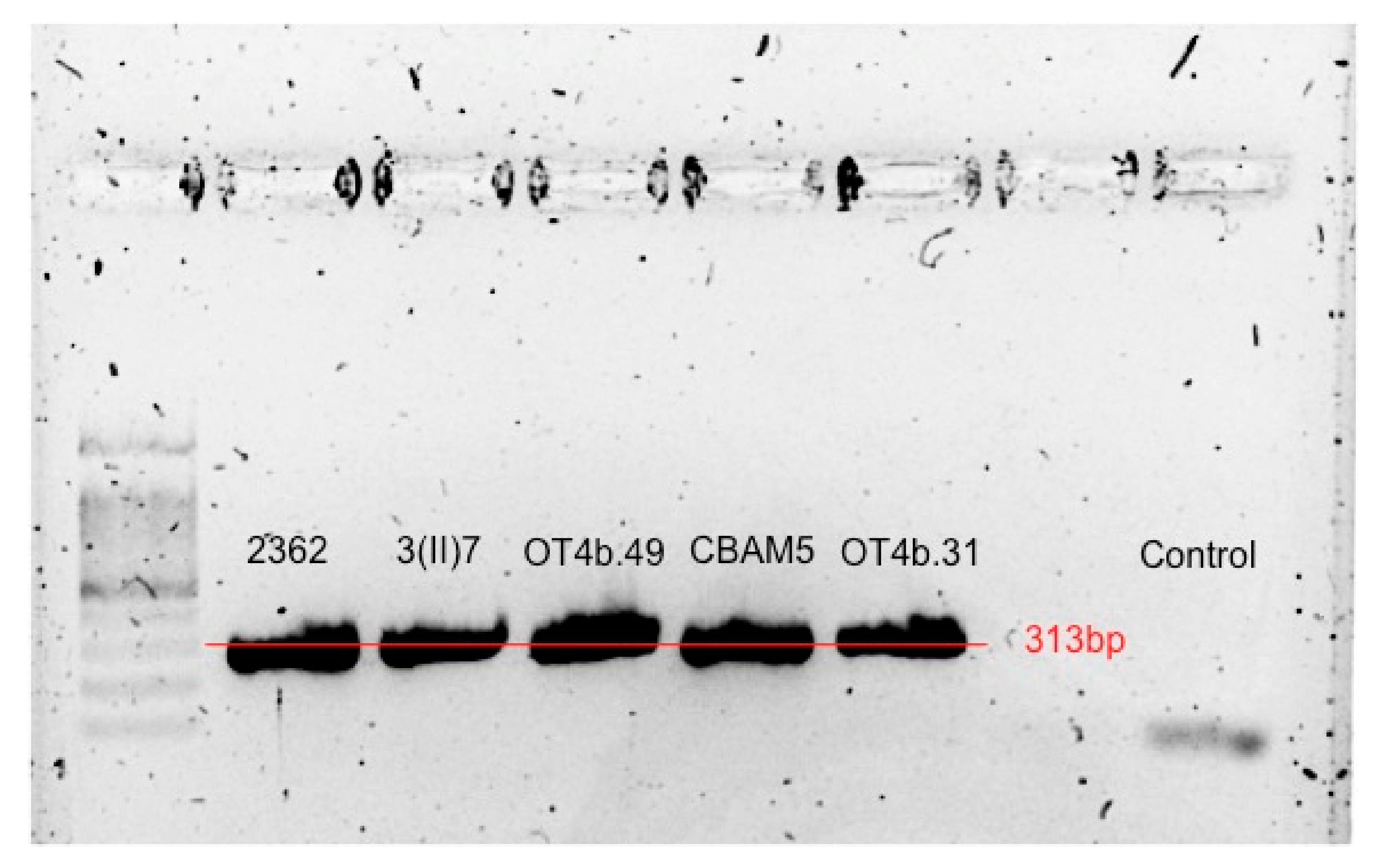
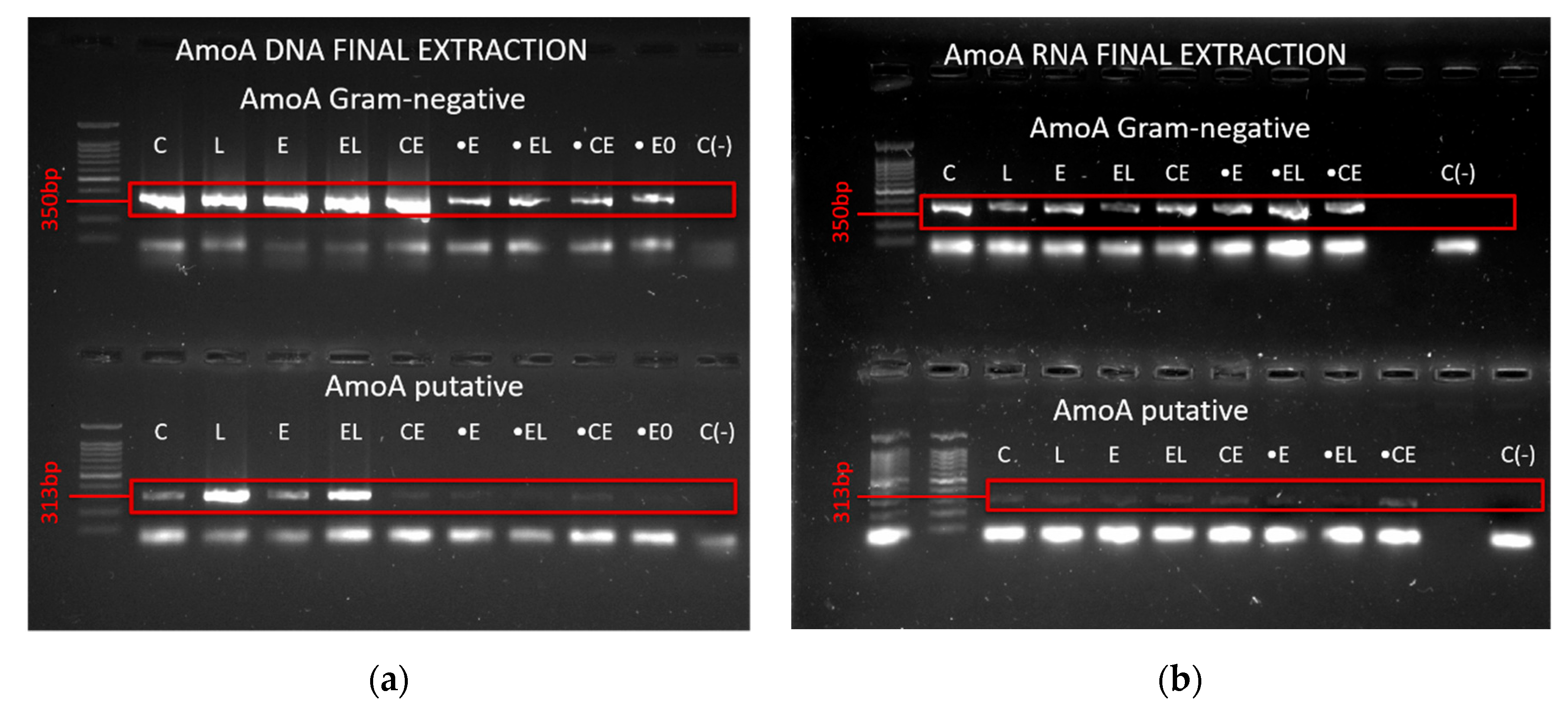
3.4. Presence and Expression of AmoA and Molybdenum Transporter Genes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Finer, M.; Jenkins, C.N.; Pimm, S.L.; Keane, B.; Ross, C. Oil and gas projects in the western Amazon: Threats to wilderness, biodiversity, and indigenous peoples. PLoS ONE 2008, 3, e2932. [Google Scholar] [CrossRef] [PubMed]
- Ron, E.Z.; Rosenberg, E. Biosurfactants and oil bioremediation. Curr. Opin. iBiotech. 2002, 13, 249–252. [Google Scholar] [CrossRef]
- Wilson, S.C.; Jones, K.C. Bioremediation of soil contaminated with polynuclear aromatic hydrocarbons (PAHs): A review. Environ. Pollut. 1993, 81, 229–249. [Google Scholar] [CrossRef]
- Manchola, L.; Dussán, J. Lysinibacillus sphaericus and Geobacillus sp. biodegradation of petroleum hydrocarbons and biosurfactant production. Remediat. J. 2014, 25, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Santana, A.; Dussán, J. Lysinibacillus sphaericus proved to have potential for the remediation of petroleum hydrocarbons. Soil Sediment Contam. 2018, 27, 538–549. [Google Scholar]
- Gómez-Garzón, C.; Hernández-Santana, A.; Dussán, J. A genome-scale metabolic reconstruction of Lysinibacillus sphaericus unveils unexploited biotechnological potentials. PLoS ONE 2017, 12, e0179666. [Google Scholar] [CrossRef] [PubMed]
- Martínez, S.A.; Dussán, J. Lysinibacillus sphaericus plant growth promoter bacteria and lead phytoremediation enhancer with Canavalia ensiformis. Environ. Prog. Sustain. Energy 2017, 37, 276–282. [Google Scholar] [CrossRef]
- Shabanamol, S.; Divya, K.; George, T.K.; Rishad, K.S.; Sreekumar, T.S.; Jisha, M.S. Characterization and in planta nitrogen fixation of plant growth promoting endophytic diazotrophic Lysinibacillus sphaericus isolated from rice (Oryza sativa). Physiol. Mol. Plant Pathol. 2018, 102, 46–54. [Google Scholar] [CrossRef]
- Schaefer, M.; Juliane, F. The influence of earthworms and organic additives on the biodegradation of oil contaminated soil. Appl. Soil Ecol. 2007, 36, 53–62. [Google Scholar] [CrossRef]
- Njoku, K.L.; Akinola, M.O.; Anigbogu, C.C. Vermiremediation of soils contaminated with mixture of petroleum products using Eisenia fetida. J. Appl. Sci. Environ. Manag. 2016, 20, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Martinkosky, L.; Barkley, J.; Sabadell, G.; Gough, H.; Davidson, S. Earthworms (Eisenia fetida) demonstrate potential for use in soil bioremediation by increasing the degradation rates of heavy crude oil hydrocarbons. Sci. Total Environ. 2017, 580, 734–743. [Google Scholar] [CrossRef]
- Contreras-Ramos, S.M.; Alvarez-Bernal, D.; Dendooven, L. Removal of polycyclic aromatic hydrocarbons from soil amended with biosolid or vermicompost in the presence of earthworms (Eisenia fetida). Soil Biol. Biochem. 2008, 40, 1954–1959. [Google Scholar] [CrossRef]
- Zhang, B.G.; Li, G.T.; Shen, T.S.; Wang, J.K.; Sun, Z. Changes in microbial biomass C, N, and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida. Soil Biol. Biochem. 2000, 32, 2055–2062. [Google Scholar] [CrossRef]
- Huang, K.; Xia, H.; Cui, G.; Li, F. Effects of earthworms on nitrification and ammonia oxidizers in vermicomposting systems for recycling of fruit and vegetable wastes. Sci. Total Environ. 2017, 578, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Singh, D.P.; Tiwari, R.; Kumar, K.; Singh, R.V.; Singh, S.; Prassana, R.; Saxena, A.K.; Nain, L. Taxonomic and functional annotation of gut bacterial communities of Eisenia foetida and Perionyx excavatus. Microbiol. Res. 2015, 175, 48–56. [Google Scholar] [CrossRef]
- Pathma, J.; Sakthivel, N. Microbial diversity ofvermicompost bacteria that exhibit useful agricultural traits and waste management potential. SpringerPlus 2012, 1, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Hickman, Z.; Reid, B. Earthworm Assisted Bioremediation of Organic Contaminants. Environ. Int. 2008, 34, 1072–1081. [Google Scholar] [CrossRef]
- Prosser, J.I.; Nicol, G.W. Archaeal and bacterial ammonia-oxidisers in soil: The quest for niche specialisation and differentiation. Trends Microbiol. 2012, 20, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junier, P.; Molina, V.; Dorador, C.; Hadas, O.; Kim, O.S.; Junier, T.; Witzel, K.P.; Imhoff, J.F. Phylogenetic and functional marker genes to study ammonia-oxidizing microorganisms (AOM) in the environment. Appl. Microbiol. Biotechnol. 2010, 85, 425–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish Health Management Division, Central Institute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar, India; National Bureau of Fish Genetics Resources, Canal Ring Road, Dilkusha, Lucknow, India. Evaluation and efficacy study of Nitrifying bacteria in freshwater Aquaculture system. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 962–969. [Google Scholar]
- Kwapisz, E.; Wszelaka, J.; Marchut, O.; Bielecki, S. The effect of nitrate and ammonium ions on kinetics of diesel oil degradation by Gordonia alkanivorans S7. Int. Biodeterior. Biodegrad. 2008, 61, 214–222. [Google Scholar] [CrossRef]
- Rasche, M.E.; Hyman, M.R.; Arp, D.J. Biodegradation of halogenated hydrocarbon fumigants by nitrifying bacteria. Appl. Environ. Microbiol. 1990, 56, 2568–2571. [Google Scholar] [CrossRef] [Green Version]
- John, R.C.; Okpokwasili, G.C. Crude oil-degradation and plasmid profile of nitrifying bacteria isolated from oil-impacted mangrove sediment in the Niger Delta of Nigeria. Bull. Environ. Contam. Toxicol. 2012, 88, 1020–1026. [Google Scholar] [CrossRef] [Green Version]
- García-Rivero, M.; Peralta-Pérez, M.R. Cometabolism in de Biodegradation of Hydrocarbons. Rev. Mex. Ing. Quim. 2008, 7, 1–12. [Google Scholar]
- Fann Instrument Company. Oil and Water Retort Instruction Manual; Fann Instrument Company: Houston TX, USA, 2019; pp. 4–6. [Google Scholar]
- Presley, M.L.; McElroy, T.C.; Diehl, W.J. Soil moisture and temperature interact to affect growth, survivorship, fecundity, and fitness in the earthworm Eisenia fetida. Comp. Biochem. Physiol. 1996, 114, 319–326. [Google Scholar] [CrossRef]
- Gómez-Garzón, C.; Dussán, J. Evidence-based validation of quorum quenching from Lysinibacillus sphaericus and Geobacillus sp. in bioremediation of oil sludge. Can. J. Microbiol. 2016, 63, 74–82. [Google Scholar] [CrossRef]
- Schuch, R.; Pelzek, A.J.; Kan, S.; Fischetti, V.A. Prevalence of Bacillus anthracis-like organisms and bacteriophages in the intestinal tract of the earthworm Eisenia fetida. Appl. Environ. Microbiol. 2010, 76, 2286–2294. [Google Scholar] [CrossRef] [Green Version]
- Burns, R.C.; Hardy, R.W. Nitrogenase: The Catalyst. In Nitrogen Fixation in Bacteria and Higher Plants, 1st ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1975; Volume 21, pp. 74–105. [Google Scholar]
- Ihssen, J.; Horn, M.A.; Matthies, C.; Gößner, A.; Schramm, A.; Drake, H.L. N2O-producing microorganisms in the gut of the earthworm Aporrectodea caliginosa are indicative of ingested soil bacteria. Appl. Environ. Microbiol. 2003, 69, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Wertz, J.T.; Kim, E.; Breznak, J.A.; Schmidt, T.M.; Rodrigues, J.L. Genomic and physiological characterization of the Verrucomicrobia isolate Diplosphaera colitermitum gen. nov., sp. nov., reveals microaerophily and nitrogen fixation genes. Appl. Environ. Microbiol. 2012, 78, 1544–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, J.C.; Caron, D.A.; Dennett, M.R. Regulation of gross growth efficiency and ammonium regeneration in bacteria by substrate C: N ratio. Limnol. Oceanogr. 1987, 32, 1239–1252. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acevedo, J.D.; Dussán, J. Ammonium Cycling and Nitrification Stimulation during Oil Sludge Remediation by Gram-Positive Bacteria Lysinibacillus sphaericus Using Red Wiggler Earthworm Eisenia fetida. AgriEngineering 2020, 2, 544-555. https://doi.org/10.3390/agriengineering2040036
Acevedo JD, Dussán J. Ammonium Cycling and Nitrification Stimulation during Oil Sludge Remediation by Gram-Positive Bacteria Lysinibacillus sphaericus Using Red Wiggler Earthworm Eisenia fetida. AgriEngineering. 2020; 2(4):544-555. https://doi.org/10.3390/agriengineering2040036
Chicago/Turabian StyleAcevedo, Juan Diego, and Jenny Dussán. 2020. "Ammonium Cycling and Nitrification Stimulation during Oil Sludge Remediation by Gram-Positive Bacteria Lysinibacillus sphaericus Using Red Wiggler Earthworm Eisenia fetida" AgriEngineering 2, no. 4: 544-555. https://doi.org/10.3390/agriengineering2040036
APA StyleAcevedo, J. D., & Dussán, J. (2020). Ammonium Cycling and Nitrification Stimulation during Oil Sludge Remediation by Gram-Positive Bacteria Lysinibacillus sphaericus Using Red Wiggler Earthworm Eisenia fetida. AgriEngineering, 2(4), 544-555. https://doi.org/10.3390/agriengineering2040036