
An Update on Phytochemicals in Redox Homeostasis: “Virtuous or Evil” in Cancer Chemoprevention?



Abstract
:
1. Introduction
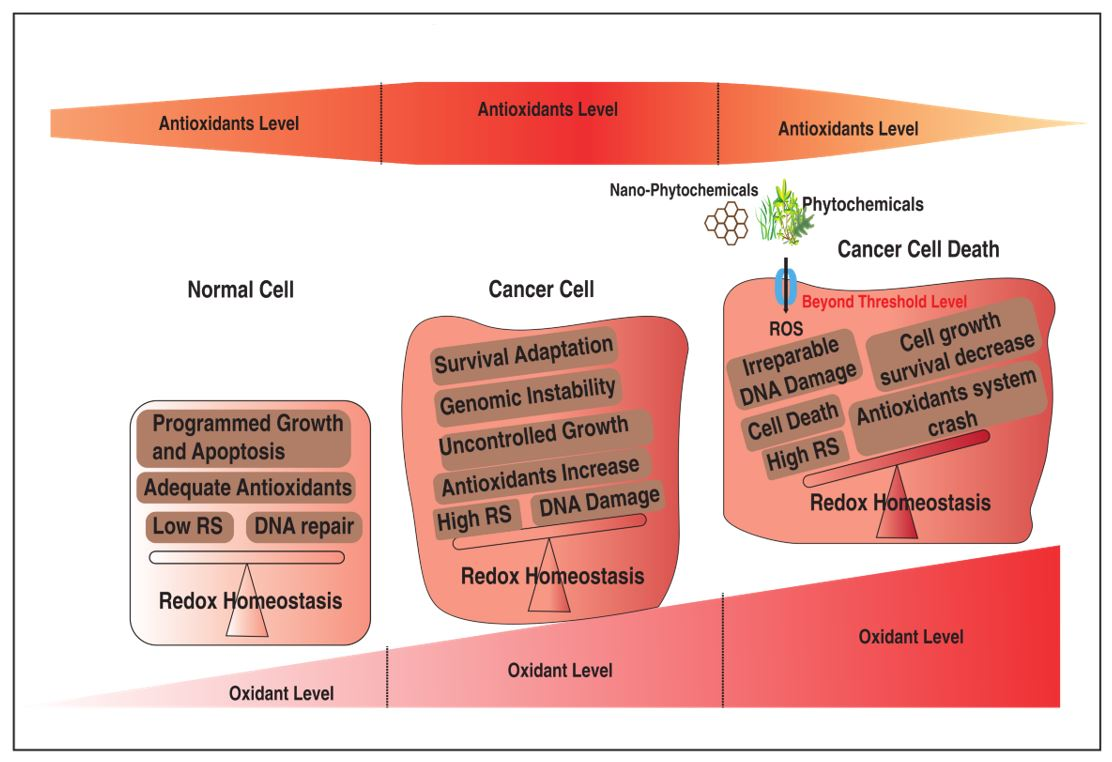
2. Cellular Redox Dynamics in Cancer Pathogenesis
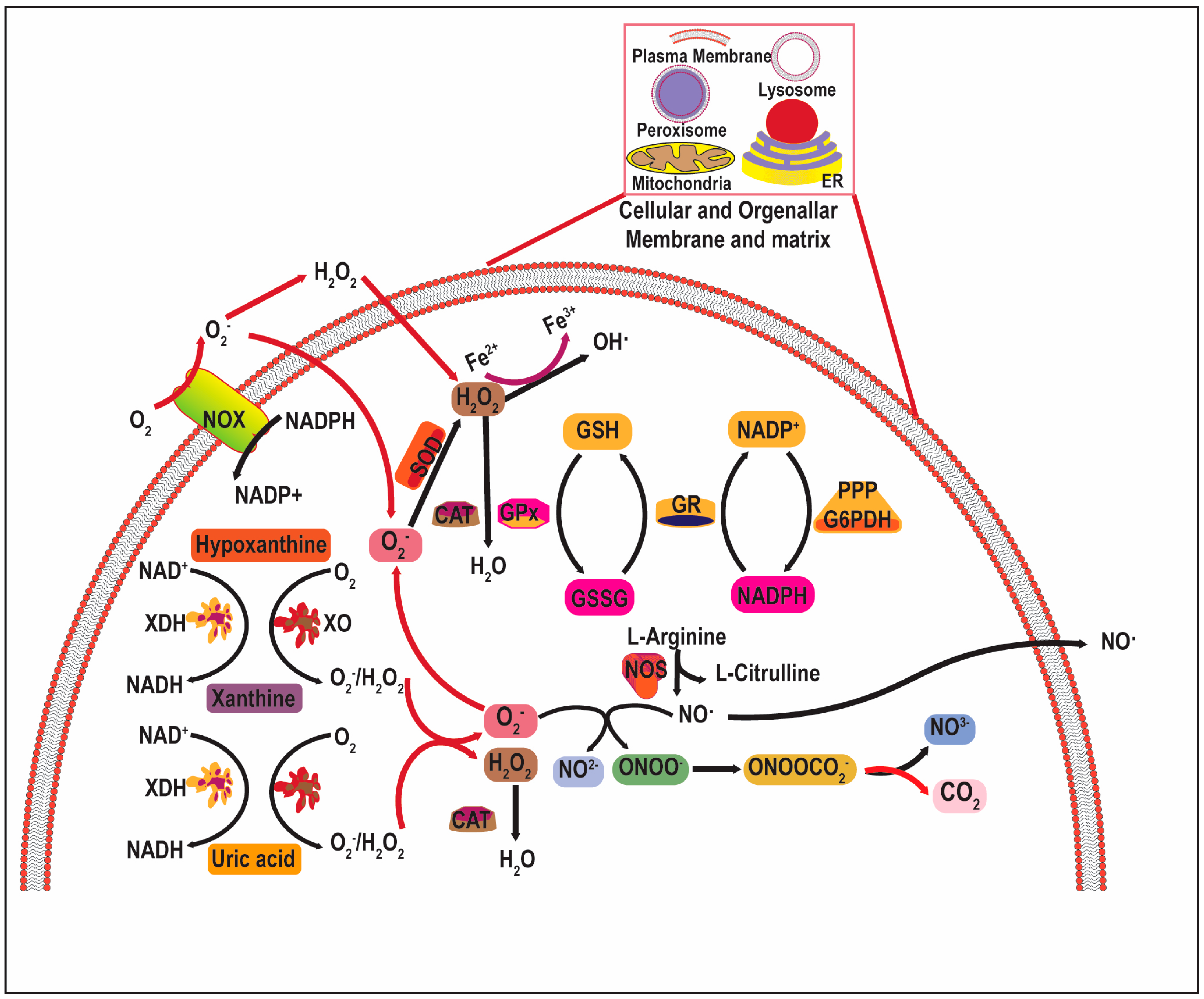
2.1. Source and Chemistry of Cellular Redox Dynamics
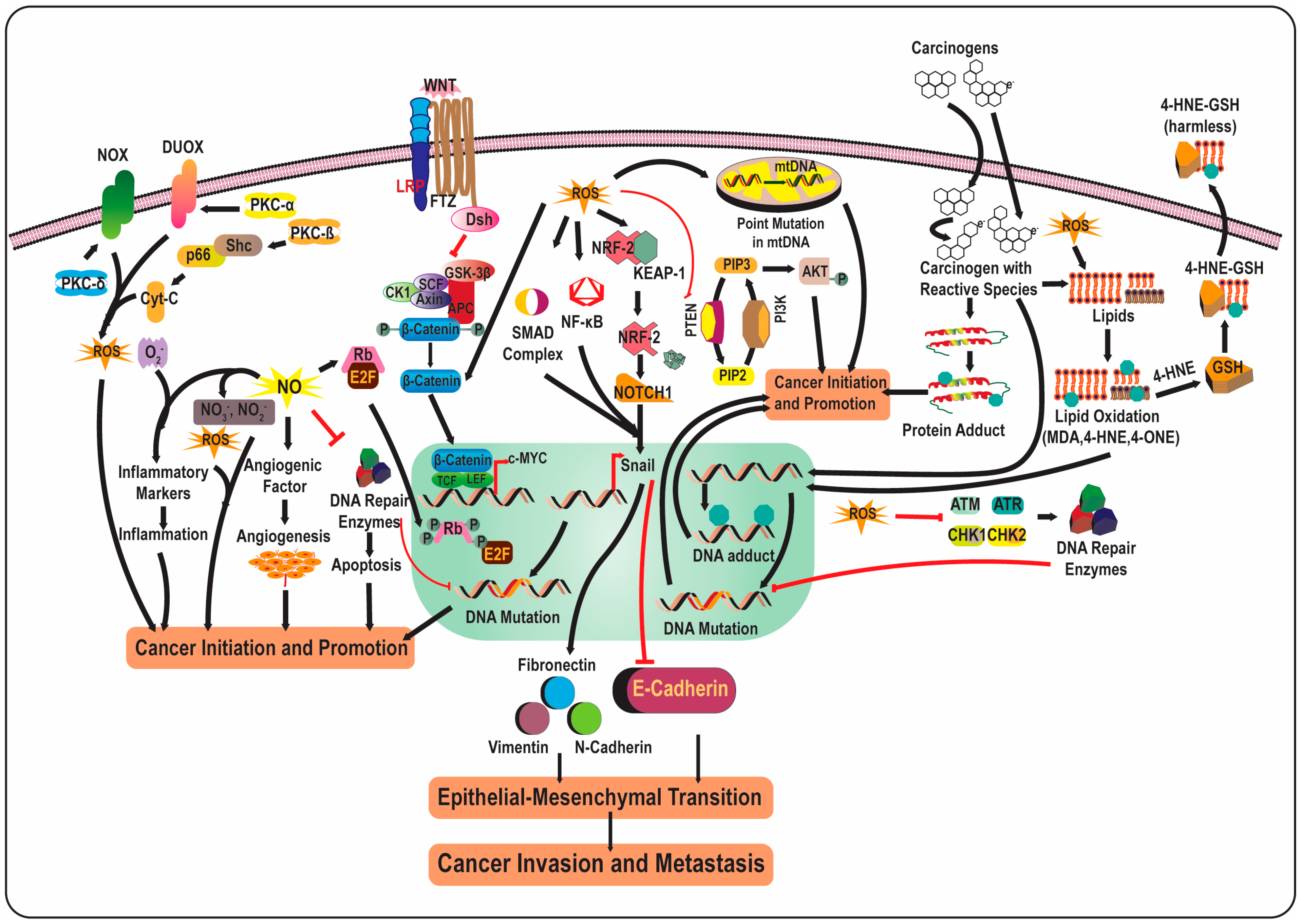
2.2. Free Radicals in Cancer Biology
3. Limitations to the Conventional Cancer Therapy and Role of Alternative Medicine
4. Phytochemicals in Redox Homeostasis: Friend or Foe in Cancer Pathogenesis?
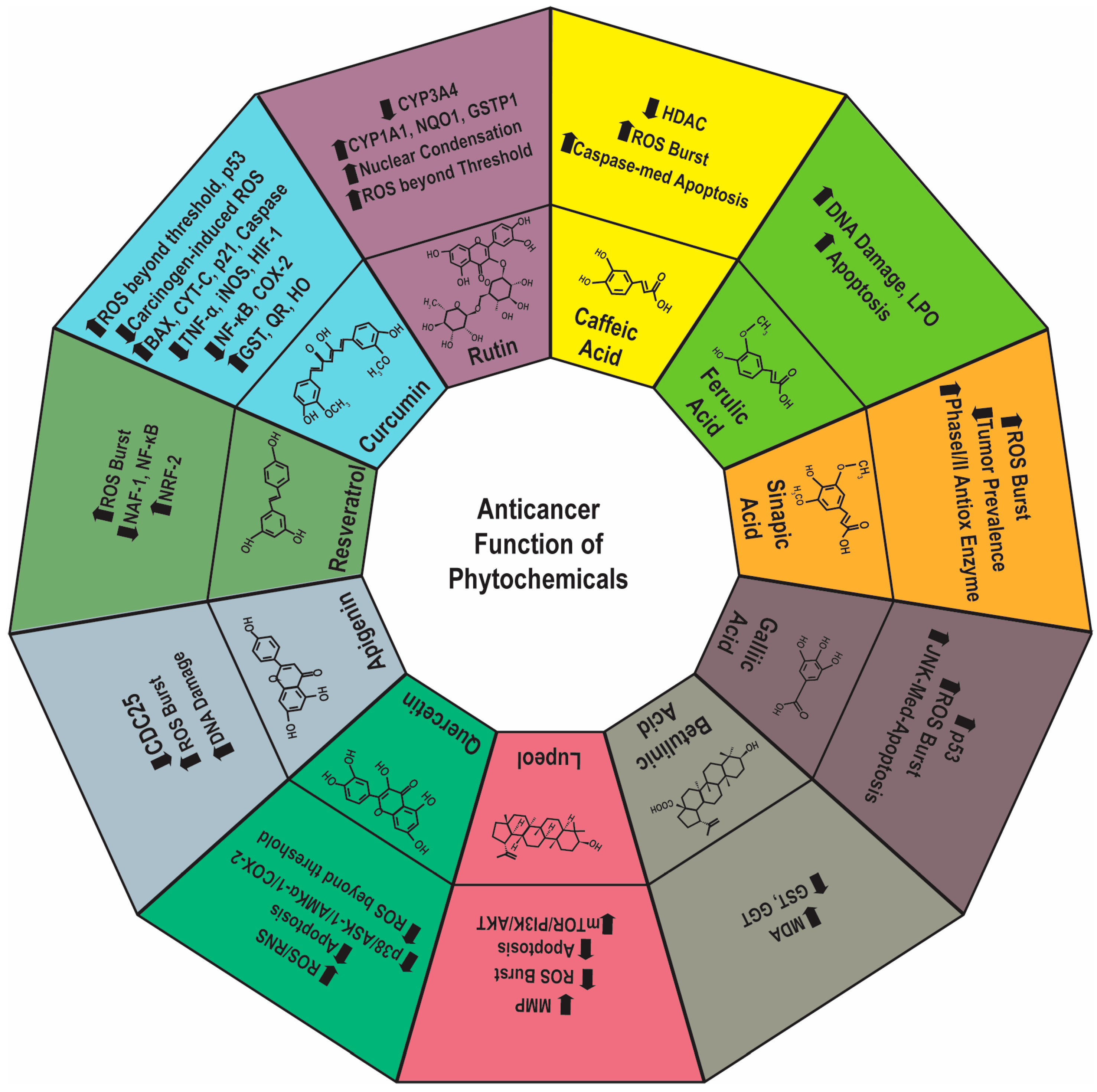
4.1. Phytochemicals as a Chemopreventive Agent

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Found in | Function | Role in Redox Balance | Ref. |
|---|---|---|---|---|
| Curcumin | Turmeric | Anticancer | Induce Glutathione S-transferase, Quinine reductase, and Hemeoxygenase, induce apoptosis by upregulating the expression of ROS beyond a threshold level, Ca2+, BAX, Cyt C, p53, p2, Caspase 3 and reducing MMP, inhibit iNOS and TNF-α, HIF-1, hypoxia-induced ROS | [61,62,63,64] |
| Resveratrol | Grapes | Anticancer | Facilitate Nrf-2 expression, overproduction of ROS, suppresses NAF-1 and iNOS expression, and post-translation modification and translocation of NF-κB | [65] |
| Apigenin | Parsley, Chamomile, Celery, Vine-Spinach, Artichokes, and Oregano | Organ protective and Anticancer | Inhibiting the expression of Cdc25c, overproduction of ROS, DNA damage | [66,67,68] |
| Quercetin | Citrus fruits, Apples, Onions, Parsley, Sage, Tea, Red wine, Olive oil, Grapes, Cherries, Blueberries, Blackberries, and Bilberries | Chemopreventive | Scavenge ROS and RNS, enhance Paclitaxel efficacy, ER-stress and increase ROS beyond a threshold level, induce free radical-mediated apoptosis by p38/ASK1/AMPKα1/COX2. | [69,70,71] |
| Rutin | Passionflower, Buckwheat, Tea, and Apples | Anticancer | Alter the expression of CYP3A4 and CYP1A1, NQO1, and GST variant P1. Enhances ROS beyond threshold level and nuclear condensation | [72,73] |
| Caffeic acid | Coffee, Red wine, Berries, and Apples | Anticancer | Inhibit HDAC2, overproduction of ROS, cell cycle arrest, Caspase-3-mediated apoptosis | [75] |
| Ferulic acid | Rice, Wheat, Oats, Pineapple, Grasses, Grains, Beans, Coffee Beans, Artichokes, and Peanuts | Cytoprotective | Scavenge ROS, inhibit DNA damage, inflammation, LPO, stimulate apoptosis | [77] |
| Sinapic acid | Spices, Citrus, Berries, Fruits, Vegetables, Cereals, and Oilseed crops | Chemopreventive | Decrease tumor prevalence, modulate LPO markers, increase phase I and phase II detoxifying enzymes. Increase ROS, oxidative stress, mitochondrial-dependent apoptosis | [78,79] |
| Gallic acid | Hazel, Tea Leaves, and Oak Barks | Anticancer | Increases ROS, decreases GSH, MMP loss, activates p53, facilitates JNK-mediated apoptosis | [80] |
| Betulinic acid | Birch, Eucalyptus, and Plane trees | Anticancer | Neutralizes ROS, upregulates GST, γ-glutamyl transpeptidase, and DT-diaphorase, reduces MDA levels | [81] |
| Lupeol | White cabbage, Green pepper, Strawberry, Olive, Mangoes, and Grapes | Anticancer | Excessive ROS generation, apoptosis, downregulation of m-TOR/PI3K/AKT axis, loss of MMP | [82] |
| Capsaicin | Chilli pepper, Oregano, Cinnamon, and Cilantro | Carcinogenic | Increase tumoral load and prevalence, histone modification by HDAC and TLR4 dysregulation | [83,84] |
| Cycasin | Cycad nuts | Carcinogenic | Promotes neoplasia | [85] |
| β-myrcene | Verbane, Lemongrass, Bay, Rosemary, Basil, Cardamom | Carcinogenic | Promote adenomas and carcinoma | |
| Alkylbenzenes | Artemisia dranunculus, Nutmeg | Carcinogenic | Form DNA adducts, micronuclei, malignant tumors | [86] |
| Coumarin | Cinnamon, Tonka Beans, and Sweet Clover. | Carcinogenic | Adenomas and carcinomas | |
| Safrole and Methyleugenol | Artemisia dranunculus, Nutmeg | Carcinogenic | Genotoxicity, mutagenicity, chromosomal aberrations | [87,88] |
| Aristocholic acid | Birthworts or pipevines and Asarum | Carcinogenic | DNA damage, DNA adduct, premalignant alterations | [88] |
| Isothiocyanates | Cruciferous, Watercress, and Radish | Carcinogenic | Papillary of nodular hyperplasia and carcinoma | [89] |
4.2. Toxic Effect of Phytochemicals
5. Nano-Phytomedicine Is a Hope for Improved Cancer Therapeutics: Evidence from Preclinical Studies
6. Ascendency of Phytochemicals Mediated Combination Therapy in Chemoprevention
7. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-HNE | 4-Hydroxy-2-Nonenal |
| 4-ONE | 4-Oxo-2-Nonenal |
| ATM | Ataxia-Telengiectasia-Mutated |
| ATR | ATM and Rad-3 Related |
| CHK1/2 | Checkpoint Kinase |
| DMBA | 7,12-Dimethylbenz[a]anthracene |
| GPx | Glutathione Peroxidase |
| MMP | Matrix Metalloproteinase |
| NOS | Nitric Oxide Synthase |
| NOX | NADPH Oxidase |
| NRF-2 | Nuclear Factor Related-Erythroid 2 |
| PI3K | Phosphoinositide 3-Kinase |
| PIP2/3 | Phosphatidylinositol di/trisphosphate |
| PTEN | Phosphatase and Tensin Homolog |
| PUFA | Polyunsaturated Fatty Acid |
| RNS | Reactive Nitrogen Species |
| ROS | Reactive Oxygen Species |
| SOD | Superoxide Dismutase |
| TPA | 12-O-Tetradecanoylphorbol-13-Acetate |
| TR | Thioredoxin Reductase |
References
- Parvez, S.; Long, M.J.C.; Poganik, J.R.; Aye, Y. Redox Signaling by Reactive Electrophiles and Oxidants. Chem. Rev. 2018, 118, 8798–8888. [Google Scholar] [CrossRef] [PubMed]
- Harris, I.S.; DeNicola, G.M. The Complex Interplay between Antioxidants and ROS in Cancer. Trends Cell. Biol. 2020, 30, 440–451. [Google Scholar] [CrossRef]
- Klaunig, J.E. Oxidative Stress and Cancer: Ingenta Connect. Curr. Pharm. Des. 2018, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Singh, D.; Fatma, H.; Akhtar, K.; Arjmand, F.; Maurya, S.K.; Siddique, H.R. Antiandrogen enzalutamide induced genetic, cellular, and hepatic damages: Amelioration by triterpene Lupeol. Drug Chem. Toxicol. 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Tania, M.; Zhang, D.Z.; Chen, C.H. Antioxidant enzymes and cancer. Chin. J. Cancer Res. 2010, 22, 87–92. [Google Scholar] [CrossRef]
- Sakanyan, V. Reactive Chemicals and Electrophilic Stress in Cancer: A Minireview. High Throughput 2018, 7, 12. [Google Scholar] [CrossRef]
- Nirgude, S.; Choudhary, B. Insights into the role of GPX3, a highly efficient plasma antioxidant, in cancer. Biochem. Pharmacol. 2021, 184, 114365. [Google Scholar] [CrossRef]
- Gaikwad, S.; Srivastava, S.K. Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer. Antioxidants 2021, 10, 83. [Google Scholar] [CrossRef]
- Fatma, H.; Siddique, H.R. Research and Patents Status of Selected Phytochemicals Against Cancer: How Close and How Far? Recent Pat. Anticancer Drug Discov. 2022, 18. [Google Scholar] [CrossRef]
- Georgescu, S.R.; Sârbu, M.I.; Matei, C.; Ilie, M.A.; Caruntu, C.; Constantin, C.; Neagu, M.; Tampa, M. Capsaicin: Friend or Foe in Skin Cancer and Other Related Malignancies? Nutrients 2017, 9, 1365. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Y.; Loscalzo, J. Real-Time Assessment of the Metabolic Profile of Living Cells with Genetically Encoded NADH Sensors. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2014; Volume 542, pp. 349–367. [Google Scholar]
- Held, J.M. Redox Systems Biology: Harnessing the Sentinels of the Cysteine Redoxome. Antioxid. Redox Signal. 2020, 32, 659–676. [Google Scholar] [CrossRef]
- Bayr, H. Reactive oxygen species. Crit. Care Med. 2005, 33, S498–S501. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, H.; Liu, Y.; Duan, C.; Liu, X.; Xia, T.; Chen, D.; Piao, H.L.; Liu, H.X. The double-edged roles of ROS in cancer prevention and therapy. Theranostics 2021, 11, 4839–4857. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, A.; Ogun, M.; Ozcan, A.; Ogun, M. Biochemistry of Reactive Oxygen and Nitrogen Species. In Basic Principles and Clinical Significance of Oxidative Stress; Gowder, S.J.T., Ed.; IntechOpen: London, UK, 2015; Volume 3, pp. 37–58. [Google Scholar]
- Ahmad, R. Introductory chapter: Basics of free radicals and antioxidants. In Free Radicals, Antioxidants and Diseases; Ahmad, R., Ed.; IntechOpen: London, UK, 2018; Volume 1, pp. 18–21. [Google Scholar]
- Porter, N.A.; Xu, L.; Pratt, D.A. Reactive Sterol Electrophiles: Mechanisms of Formation and Reactions with Proteins and Amino Acid Nucleophiles. Chemistry 2020, 2, 390–417. [Google Scholar] [CrossRef]
- Dantas, L.S.; Viviani, L.G.; Inague, A.; Piccirillo, E.; de Rezende, L.; Ronsein, G.E.; Augusto, O.; de Medeiros, M.H.; Antonia, T.; Miyamoto, S. Lipid-derived electrophiles induce covalent modification and aggregation of Cu, Zn-superoxide dismutase in a hydrophobicity-dependent manner. Free Radic. Biol. Med. 2016, 100, S65. [Google Scholar] [CrossRef]
- Martinez, K.B.; Mackert, J.D.; McIntosh, M.K. Polyphenols and Intestinal Health. In Nutrition and Functional Foods for Healthy Aging; Watson, R.R., Ed.; Academic Press: Cambridge, UK, 2017; pp. 191–210. [Google Scholar]
- Adams, L.; Franco, M.C.; Estevez, A.G. Reactive nitrogen species in cellular signaling. Exp. Biol. Med. 2015, 240, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.C.; Andriantsitohaina, R. Reactive Nitrogen Species: Molecular Mechanisms and Potential Significance in Health and Disease. Antioxid. Redox Signal. 2009, 11, 669–702. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Slusarenko, A.J. The biology of reactive sulfur species (RSS). Plant Physiol. Biochem. 2012, 1, 98–107. [Google Scholar] [CrossRef]
- Olson, K.R. Reactive oxygen species or reactive sulfur species: Why we should consider the latter. J. Exp. Biol. 2020, 223 Pt 4, jeb19635226. [Google Scholar] [CrossRef]
- Giles, G.I.; Jacob, C. Reactive Sulfur Species: An Emerging Concept in Oxidative Stress. Biol. Chem. 2002, 383, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Lau, N.; Pluth, M.D. Reactive sulfur species (RSS): Persulfides, polysulfides, potential, and problems. Curr. Opin. Chem. Biol. 2019, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug. Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Adwas, A.A.; Elsayed, A.; Azab, A.E.; Quwaydir, F.A. Oxidative stress and antioxidant mechanisms in human body. J. Appl. Biotechnol. Bioeng. 2019, 6, 43–47. [Google Scholar]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Tasdogan, A.; Ubellacker, J.M.; Morrison, S.J. Redox regulation in cancer cells during metastasis. Cancer Discov. 2021, 11, 2682. [Google Scholar] [CrossRef] [PubMed]
- Maurya, S.K.; Fatma, H.; Maurya, A.K.; Mishra, N.; Siddique, H.R. Role of lupeol in chemosensitizing therapy-resistant prostate cancer cells by targeting MYC, β-catenin and c-FLIP: In silico and in vitro studies. In Silico Pharmacol. 2022, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Mydin, R.B.S.; Okekpa, S.I. Reactive Oxygen Species, Cellular Redox Homeostasis and Cancer. In Homeostasis—An Integrated Vision; Lasakosvitsch, F., Garnes, S.D.A., Eds.; IntechOpen: London, UK, 2018. [Google Scholar]
- Yuan, X.; Wang, B.; Yang, L.; Zhang, Y. The role of ROS-induced autophagy in hepatocellular carcinoma. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 306–312. [Google Scholar] [CrossRef]
- Sullivan, L.B.; Chandel, N.S. Mitochondrial reactive oxygen species and cancer. Cancer Metab. 2014, 2, 17. [Google Scholar] [CrossRef]
- Saikolappan, S.; Kumar, B.; Shishodia, G.; Koul, S.; Koul, H.K. Reactive oxygen species and cancer: A complex interaction. Cancer Lett. 2019, 452, 132–143. [Google Scholar] [CrossRef]
- Liu, R.; Wang, H.L.; Deng, M.J.; Wen, X.J.; Mo, Y.Y.; Chen, F.M.; Daun, W.F.; Li, L.; Nei, X. Melatonin Inhibits Reactive Oxygen Species-Driven Proliferation, Epithelial-Mesenchymal Transition, and Vasculogenic Mimicry in Oral Cancer. Oxid. Med. Cell. Longev. 2018, 2018, e3510970. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Nie, X.; Yao, W.; Klinghammer, K.; Sudhoff, H.; Kaufmann, A.M.; Albers, A.E. Reactive oxygen species in cancer stem cells of head and neck squamous cancer. Semin. Cancer Biol. 2018, 53, 248–257. [Google Scholar] [CrossRef]
- Liskova, A.; Stefanicka, P.; Samec, M.; Smejkal, K.; Zubor, P.; Bielik, T.; Biskupska-Bodova, K.; Kwon, T.K.; Danko, J.; Büsselberg, D.; et al. Dietary phytochemicals as the potential protectors against carcinogenesis and their role in cancer chemoprevention. Clin. Exp. Med. 2020, 20, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Chen, H.; Liang, J.; Li, Y.; Yang, J.; Luo, C.; Tang, Y.; Ding, Y.; Liu, X.; Yuan, Q.; et al. Dual Role of Reactive Oxygen Species and their Application in Cancer Therapy. J. Cancer 2021, 12, 5543–5561. [Google Scholar] [CrossRef] [PubMed]
- Viedma-Poyatos, Á.; González-Jiménez, P.; Langlois, O.; Company-Marín, I.; Spickett, C.M.; Pérez-Sala, D. Protein Lipoxidation: Basic Concepts and Emerging Roles. Antioxidants 2021, 10, 295. [Google Scholar] [CrossRef]
- Martín-Sierra, C.; Laranjeira, P.; Domingues, M.R.; Paiva, A. Lipoxidation and cancer immunity. Redox Biol. 2019, 23, 101103. [Google Scholar] [CrossRef]
- Bahar, G.; Feinmesser, R.; Shpitzer, T.; Popovtzer, A.; Nagler, R.M. Salivary analysis in oral cancer patients. Cancer 2007, 109, 54–59. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Lutsenko, S.V.; Terentiev, A.A. Reactive Oxygen and Nitrogen Species–Induced Protein Modifications: Implication in Carcinogenesis and Anticancer Therapy. Cancer Res. 2018, 78, 6040–6047. [Google Scholar] [CrossRef]
- O’Rourke, J.M.; Sagar, V.M.; Shah, T.; Shetty, S. Carcinogenesis on the background of liver fibrosis: Implications for the management of hepatocellular cancer. World J. Gastroenterol. 2018, 24, 4436–4447. [Google Scholar] [CrossRef]
- Chidambaram, M.; Manavalan, R.; Kathiresan, K. Nanotherapeutics to Overcome Conventional Cancer Chemotherapy Limitations. J. Pharm. Pharm. Sci. 2011, 14, 67–77. [Google Scholar] [CrossRef]
- Maurya, S.K.; Shadab, G.G.H.A.; Siddique, H.R. Chemosensitization of Therapy Resistant Tumors: Targeting Multiple Cell Signaling Pathways by Lupeol, A Pentacyclic Triterpene. Curr. Pharm. Des. 2020, 26, 455–465. [Google Scholar] [CrossRef]
- Fatma, H.; Siddique, H.R.; Maurya, S.K. The multiple faces of NANOG in cancer: A therapeutic target to chemosensitize therapy-resistant cancers. Epigenomics 2021, 13, 1885–1900. [Google Scholar] [CrossRef]
- Siddique, H.R.; Feldman, D.E.; Chen, C.L.; Punj, V.; Tokumitsu, H.; Machida, K. NUMB phosphorylation destabilizes p53 and promotes self-renewal of tumor-initiating cells by a NANOG-dependent mechanism in liver cancer. Hepatology 2015, 62, 1466–1479. [Google Scholar] [CrossRef] [PubMed]
- Fatma, H.; Siddique, H.R. Pluripotency inducing Yamanaka factors: Role in stemness and chemoresistance of liver cancer. Expert Rev. Anticancer Ther. 2021, 21, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Singh, D.; Arif, A.; Sodhi, K.K.; Singh, D.K.; Islam, S.N.; Ahmad, A.; Akhtar, K.; Siddique, H.R. Protective effect of green synthesized Selenium Nanoparticles against Doxorubicin induced multiple adverse effects in Swiss albino mice. Life Sci. 2022, 305, 120792. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Khan, M.A.; Akhtar, K.; Arjmand, F.; Siddique, H.R. Apigenin alleviates cancer drug Sorafenib induced multiple toxic effects in Swiss albino mice via antioxidative stress. Toxicol. Appl. Pharmacol. 2022, 447, 116072. [Google Scholar] [CrossRef]
- Wirsdörfer, F.; de Leve, S.; Jendrossek, V. Combining Radiotherapy and Immunotherapy in Lung Cancer: Can We Expect Limitations Due to Altered Normal Tissue Toxicity? Int. J. Mol. Sci. 2018, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Münstedt, K.; Momm, F.; Hübner, J. Honey in the management of side effects of radiotherapy- or radio/chemotherapy-induced oral mucositis: A systematic review. Complement. Ther. Clin. Pract. 2019, 34, 145–152. [Google Scholar] [CrossRef]
- Majeed, H.; Gupta, V. Adverse Effects of Radiation Therapy. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Meybodi, N.M.; Mortazavian, A.M.; Monfared, A.B.; Sohrabvandi, S.; Meybodi, F.A. Phytochemicals in Cancer Prevention: A Review of the Evidence. Int. J. Cancer Manag. 2017, 10, e7219. [Google Scholar]
- Singh, D.; Gupta, M.; Sarwat, M.; Siddique, H.R. Apigenin in cancer prevention and therapy: A systematic review and meta-analysis of animal models. Crit. Rev. Oncol. Hematol. 2022, 176, 103751. [Google Scholar] [CrossRef] [PubMed]
- Asemani, Y.; Zamani, N.; Bayat, M.; Amirghofran, Z. Allium vegetables for possible future of cancer treatment. Phytother. Res. 2019, 33, 3019–3039. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its’ Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Park, W.; Amin, A.R.; Chen, Z.G.; Shin, D.M. New Perspectives of Curcumin in Cancer Prevention. Cancer Prev. Res. 2013, 6, 387–400. [Google Scholar] [CrossRef]
- Mortezaee, K.; Salehi, E.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Najafi, M.; Farhood, B.; Rosengren, R.J.; Sahebkar, A. Mechanisms of apoptosis modulation by Curcumin: Implications for cancer therapy. J. Cell. Physiol. 2019, 234, 12537–12550. [Google Scholar] [CrossRef]
- Su, C.C.; Lin, J.G.; Li, T.M.; Chung, J.G.; Yang, J.S.; Ip, S.W.; Lin, W.C.; Chen, G.W. Curcumin-induced apoptosis of human colon cancer colo 205 cells through the production of R.O.S., Ca2+ and the activation of caspase-3. Anticancer Res. 2006, 26, 4379–4389. [Google Scholar] [PubMed]
- Gupta, N.; Verma, K.; Nalla, S.; Kulshreshtha, A.; Lall, R.; Prasad, S. Free Radicals as a Double-Edged Sword: The Cancer Preventive and Therapeutic Roles of Curcumin. Molecules 2020, 25, 5390. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef]
- Singh, D.; Khan, M.A.; Siddique, H.R. Apigenin, A Plant Flavone Playing Noble Roles in Cancer Prevention Via Modulation of Key Cell Signaling Networks. Recent Pat. Anticancer Drug Discov. 2019, 14, 298–311. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anticancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Warkad, M.S.; Kim, C.H.; Kang, B.G.; Park, S.H.; Jung, J.S.; Feng, J.H.; Inci, G.; Kim, S.C.; Suh, H.W.; Lim, S.S.; et al. Metformin-induced R.O.S. upregulation as amplified by Apigenin causes profound anticancer activity while sparing normal cells. Sci. Rep. 2021, 11, 14002. [Google Scholar] [CrossRef]
- Deepika; Maurya, P.K. Health Benefits of Quercetin in Age-Related Diseases. Molecules 2022, 27, 2498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, J.; Yu, C.; Xiang, L.; Li, L.; Shi, D.; Lin, F. Quercetin Enhanced Paclitaxel Therapeutic Effects Towards PC-3 Prostate Cancer Through ER Stress Induction and ROS Production. OncoTargets Ther. 2020, 13, 513–523. [Google Scholar] [CrossRef]
- Biswas, P.; Dey, D.; Biswas, P.K.; Rahaman, T.I.; Saha, S.; Parvez, A.; Khan, D.A.; Lily, N.J.; Saha, K.; Sohel, M.; et al. A Comprehensive Analysis and Anti-Cancer Activities of Quercetin in ROS-Mediated Cancer and Cancer Stem Cells. Int. J. Mol. Sci. 2022, 23, 11746. [Google Scholar] [CrossRef] [PubMed]
- Karakurt, S. Modulatory effects of Rutin on the expression of cytochrome P450s and antioxidant enzymes in human hepatoma cells. Acta Pharm. 2016, 66, 491–502. [Google Scholar] [CrossRef]
- Imani, A.; Maleki, N.; Bohlouli, S.; Kouhsoltani, M.; Sharifi, S.; Maleki Dizaj, S. Molecular mechanisms of anticancer effect of Rutin. Phytother. Res. 2021, 35, 2500–2513. [Google Scholar] [CrossRef]
- Birková, A.; Hubková, B.; Bolerázska, B.; Mareková, M.; Čižmárová, B. Caffeic acid: A brief overview of its presence, metabolism, and bioactivity. Bioact. Compd. Health Dis. 2020, 3, 74–81. [Google Scholar] [CrossRef]
- Anantharaju, P.G.; Reddy, D.B.; Padukudru, M.A.; Chitturi, C.M.K.; Vimalambike, M.G.; Madhunapantula, S.V. Induction of colon and cervical cancer cell death by cinnamic acid derivatives is mediated through the inhibition of Histone Deacetylases (HDAC). PLoS ONE 2017, 12, e0186208. [Google Scholar] [CrossRef]
- Damasceno, S.S.; Dantas, B.B.; Ribeiro-Filho, J.; Antônio, M.; Araújo, D.; da Costa, J.G.M. Chemical Properties of Caffeic and Ferulic Acids in Biological System: Implications in Cancer Therapy: A Review. Curr. Pharm. Des. 2017, 23, 3015–3023. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, D.; Jiang, R.; Li, H.; Wan, J.; Li, H. Ferulic acid exerts antitumor activity and inhibits metastasis in breast cancer cells by regulating epithelial to mesenchymal transition. Oncol. Rep. 2016, 36, 271–278. [Google Scholar] [CrossRef]
- Balaji, C.; Muthukumaran, J.; Nalini, N. Chemopreventive effect of sinapic acid on 1,2-dimethylhydrazine-induced experimental rat colon carcinogenesis. Hum. Exp. Toxicol. 2014, 33, 1253–1268. [Google Scholar] [CrossRef]
- Badr, D.A.; Amer, M.E.; Abd-Elhay, W.M.; Nasr, M.S.; Abuamara, T.M.; Ali, H.; Mohamed, A.F.; Youssef, M.A.; Awwad, N.S.; Ju, Y.H.; et al. Histopathological and genetic changes proved the anticancer potential of free and nano-capsulated sinapic acid. Appl. Biol. Chem. 2019, 62, 59. [Google Scholar] [CrossRef]
- Subramanian, A.P.; John, A.A.; Vellayappan, M.V.; Balaji, A.; Jaganathan, S.K.; Supriyanto, E.; Yusof, M. Gallic acid: Prospects and molecular mechanisms of its anticancer activity. RSC Adv. 2015, 5, 35608–35621. [Google Scholar] [CrossRef]
- Kaur, R.; Arora, S. Interactions of betulinic acid with xenobiotic metabolizing and antioxidative enzymes in DMBA-treated Sprague Dawley female rats. Free Radic. Biol. Med. 2013, 65, 131–142. [Google Scholar] [CrossRef]
- He, W.; Li, X.; Xia, S. Lupeol triterpene exhibits potent antitumor effects in A427 human lung carcinoma cells via mitochondrial mediated apoptosis, ROS generation, loss of mitochondrial membrane potential and downregulation of m-TOR/PI3Ksol; AKT signalling pathway. J. BUON 2018, 23, 635–640. [Google Scholar]
- Liu, Z.; Zhu, P.; Tao, Y.; Shen, C.; Wang, S.; Zhao, L.; Wu, H.; Fan, F.; Lin, C.; Chen, C.; et al. Cancer-promoting effect of Capsaicin on DMBA/TPA-induced skin tumorigenesis by modulating inflammation, Erk and p38 in mice. Food Chem. Toxicol. 2015, 81, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, H.; Yu, W.; Wei, L.; Wen, B. Effect of cold exposure and Capsaicin on the expression of histone acetylation and Toll-like receptors in 1,2-dimethylhydrazine-induced colon carcinogenesis. Environ. Sci. Pollut. Res. 2021, 28, 60981–60992. [Google Scholar] [CrossRef] [PubMed]
- Kobets, T.; Smith, B.P.C.; Williams, G.M. Food-Borne Chemical Carcinogens and the Evidence for Human Cancer Risk. Foods 2022, 11, 2828. [Google Scholar] [CrossRef]
- Pauzi, N.A.M.; Cheema, M.S.; Ismail, A.; Ghazali, A.R.; Abdullah, R. Are Genotoxic and Carcinogenic Compounds Present in Malaysian Traditional Medicines and Herbal Supplements? A Review Based on the Malaysian Herbal Monograph. Malays. J. Med. Health Sci. 2021, 17, 207–222. [Google Scholar]
- Götz, M.E.; Sachse, B.; Schäfer, B.; Eisenreich, A. Myristicin and Elemicin: Potentially Toxic Alkenylbenzenes in Food. Foods 2022, 11, 1988. [Google Scholar] [CrossRef]
- Khalil, M.; Calasso, M.; Bonfrate, L.; Di Ciaula, A.; De Angelis, M.; Wang, D.Q.H.; Portincasa, P. The pros and cons of biological effects of herbs and herb-derived compounds on liver tumorigenesis. Hepatoma Res. 2022, 8, 23. [Google Scholar] [CrossRef]
- Hirose, M.; Yamaguchi, T.; Kimoto, N.; Ogawa, K.; Futakuchi, M.; Sano, M.; Shirai, T. Strong promoting activity of phenylethyl isothiocyanate and benzyl isothiocyanate on urinary bladder carcinogenesis in F344 male rats. Int. J. Cancer. 1998, 77, 773–777. [Google Scholar] [CrossRef]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [PubMed]
- Mukerjee, A.; Ranjan, A.P.; Vishwanatha, J.K. Targeted Nanocurcumin Therapy Using Annexin A2 Antibody Improves Tumor Accumulation and Therapeutic Efficacy Against Highly Metastatic Breast Cancer. J. Biomed. Nanotechnol. 2016, 12, 1374–1392. [Google Scholar] [CrossRef] [PubMed]
- Annaji, M.; Poudel, I.; Boddu, S.H.S.; Arnold, R.D.; Tiwari, A.K.; Babu, R.J. Resveratrol-loaded nanomedicines for cancer applications. Cancer Rep. 2021, 4, e1353. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Ganguly, S.; Ganguly, S.; Debnath, M.C.; Chakraborty, S.; Mukherjee, B.; Chattopadhyay, D. Apigenin-Loaded PLGA-DMSA Nanoparticles: A Novel Strategy to Treat Melanoma Lung Metastasis. Mol. Pharm. 2021, 18, 1920–1938. [Google Scholar] [CrossRef]
- Baksi, R.; Singh, D.P.; Borse, S.P.; Rana, R.; Sharma, V.; Nivsarkar, M. In vitro and in vivo anticancer efficacy potential of Quercetin loaded polymeric nanoparticles. Biomed. Pharmacother. 2018, 106, 1513–1526. [Google Scholar] [CrossRef]
- Hoai, T.T.; Yen, P.T.; Dao, T.T.B.; Long, L.H.; Anh, D.X.; Minh, L.H.; Anh, B.Q.; Thuong, N.T. Evaluation of the Cytotoxic Effect of Rutin Prenanoemulsion in Lung and Colon Cancer Cell Lines. J. Nanomater. 2020, 2020, 8867669. [Google Scholar] [CrossRef]
- Pandey, P.; Rahman, M.; Bhatt, P.C.; Beg, S.; Paul, B.; Hafeez, A.; Al-Abbasi, F.A.; Nadeem, M.S.; Baothman, O.; Anwar, F.; et al. Implication of nano-antioxidant therapy for treatment of hepatocellular carcinoma using PLGA nanoparticles of Rutin. Nanomedicine 2018, 13, 849–870. [Google Scholar] [CrossRef]
- Firouzai-Amandi, A.; Tarahomi, M.; Rahmani Youshanlouie, H.; Mosaddeghi Heris, R.; Jafari-Gharabaghlou, D.; Zarghami, N.; Dadashpour, M. Development, Characterization, and In Vitro Evaluation of Cytotoxic Activity of Rutin Loaded PCL-PEG Nanoparticles Against Skov3 Ovarian Cancer Cell. Asian Pac. J. Cancer Prev. 2022, 23, 1951–1957. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The nanomedicine revolution: Part 2: Current and future clinical applications. P T 2012, 37, 582–591. [Google Scholar]
- Mohapatra, P.; Singh, P.; Singh, D.; Sahoo, S.; Sahoo, S.K. Phytochemical based nanomedicine: A panacea for cancer treatment, present status and future prospective. OpenNano 2022, 7, 100055. [Google Scholar] [CrossRef]
- Kumar, P.; Singh, A.K.; Raj, V.; Rai, A.; Keshari, A.K.; Kumar, D.; Maity, B.; Prakash, A.; Maiti, S.; Saha, S. Poly(lactic-co-glycolic acid)-loaded nanoparticles of betulinic acid for improved treatment of hepatic cancer: Characterization, in vitro and in vivo evaluations. Int. J. Nanomed. 2018, 13, 975–990. [Google Scholar] [CrossRef]
- Cho, H.Y.; Mavi, A.; Chueng, S.T.D.; Pongkulapa, T.; Pasquale, N.; Rabie, H.; Han, J.; Kim, J.H.; Kim, T.H.; Choi, J.W.; et al. Tumor Homing Reactive Oxygen Species Nanoparticle for Enhanced Cancer Therapy. ACS Appl. Mater. Interfaces 2019, 11, 23909–23918. [Google Scholar] [CrossRef] [PubMed]
- Bayat Mokhtari, R.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhao, C.; Wang, L. Molecular-targeted agents combination therapy for cancer: Developments and potentials. Int. J. Cancer 2014, 134, 1257–1269. [Google Scholar] [CrossRef]
- Aumeeruddy, M.Z.; Mahomoodally, M.F. Combating breast cancer using combination therapy with 3 phytochemicals: Piperine, sulforaphane, and thymoquinone. Cancer 2019, 125, 1600–1611. [Google Scholar] [CrossRef]
- Ghosh, S. Cisplatin: The first metal based anticancer drug. Bioorg Chem. 2019, 88, 102925. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Wang, X.L.; He, D.H.; Cheng, Y.X. Protection against chemotherapy- and radiotherapy-induced side effects: A review based on the mechanisms and therapeutic opportunities of phytochemicals. Phytomedicine 2021, 80, 153402. [Google Scholar] [CrossRef]
- Singh, P.; Arora, D.; Shukla, Y. Enhanced chemoprevention by the combined treatment of Pterostilbene and lupeol in B[a]P-induced mouse skin tumorigenesis. Food Chem. Toxicol. 2017, 99, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Malekinejad, F.; Kheradmand, F.; Khadem-Ansari, M.H.; Malekinejad, H. Lupeol synergizes with Doxorubicin to induce anti-proliferative and apoptotic effects on breast cancer cells. Daru J. Pharm. Sci. 2022, 30, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xu, Y.; Ji, W.; Song, L.; Dai, C.; Zhan, L. Effects of exposure to benzo[a]pyrene on metastasis of breast cancer are mediated through ROS-ERK-MMP9 axis signaling. Toxicol. Lett. 2015, 234, 201–210. [Google Scholar] [CrossRef]
- Hassan, S.M.; Khalaf, M.M.; Sadek, S.A.; Abo-Youssef, A.M. Protective effects of Apigenin and Myricetinin against cisplatin-induced nephrotoxicity in mice. Pharm. Biol. 2017, 55, 766–774. [Google Scholar] [CrossRef] [PubMed]



| Phytochemical | Doses | Effect | Models | Ref. |
|---|---|---|---|---|
| Curcumin | 2% w/v for 30 days | Scavenges ROS and induce scavenging enzymes | Male mice | [61] |
| 1 kg/day for 2 days | Inhibited NF-kB | HCT116 xenograft in nude mice | [62] | |
| 5–75 μM, for 6–72 h | Inhibited COX-2 | HT-29 cells | [62] | |
| 60 μM | Inhibit p53 phosphorylation | Colon Cancer cells | [62] | |
| 0, 5, 10, 20 and 50 μM for various time periods. | Increase ROS, Ca2+, BAX, Cytochrome C, p53, and p21, Caspase 3, and reduce MMP | Colo-205 colon cancer cells | ||
| Resveratrol | 50 μM for 24 h | Suppress NAF-1 and upregulate Nrf-2, ROS | Human pancreatic cancer cell lines Panc-1, Mia paca-2, CF pac-1, and BxPC-3 | [65] |
| Apigenin | 12.5–50 μM | Overproduction of ROS, genotoxicity, and cell cycle arrest | Papillary thyroid carcinoma cells | [67] |
| (20 μM with 10 μM Metformin) or (5 mg/kg b.w with 75 mg/kg b.w. Metformin) | ROS-dependent DNA damage and antioxidant | Human pancreatic cells and mice | [68] | |
| Quercetin | 15 μM with 12.5 μM Paclitaxel | ER stress and ROS-induced DNA damage | Prostate cancer cell line | [70] |
| 0–400 μM | Increased ROS | Breast cancer, MCF-7 | [71] | |
| Rutin | 0–100 μM | Increase antioxidant status | Liver cancer, HEPG2 | [72,73] |
| 60–100 μM | ROS-generation | Cervical cancer, HPV-C33A | ||
| Caffeic acid | 0–500 μM | HDAC inhibtion and ROS generation | Cervical cancer (HeLa and SiHa) and colon cancer (HCT-116 and HCT-15) | [75] |
| Ferulic acid | 0–100 μM | Inhibit DNA and lipid damage | Cytoprotective | [77] |
| Sinapic acid | 40 mg/kg b.w. | Modulate LPO markers and increase antioxidant enzyme | Mice | [78] |
| 125.23 μM | Increase in ROS level | HeP-2 cells | [79] | |
| Gallic acid | 0–200 μM; 50 g/ml | Increase in ROS level | Lung cancer, Calu-6 and A549 | [80] |
| Betulinic acid | 10 mg/kg b.w. | Upregulate phase II antioxidant enzyme | Mice | [81] |
| Lupeol | 12.5–50 μM | Increased ROS generation | Lung cancer, A427 | [82] |
| Capsaicin | 10 mg/kg b.w. | Promoted cancer | Female mice | [83] |
| 10 mg/kg b.w. | Promoted cancer | Male Wistar rate | ||
| Cycasin | 50–75 mg/kg b.w. for 5 days | Promoted cancer | Monkey | [85] |
| β-myrcene | 1000 mg/kg b.w. for 5 days/week | Promoted cancer | Mice | |
| Coumarin | 200 mg/kg b.w. | Promoted cancer | Mice | [86] |
| Safrole and Methyleugenol | 5000 mg/kg b.w.; 0.05 μM/b.w. | Promoted cancer | Mice | [87,88] |
| Aristocholic acid | 5 mg/kg b.w. for 3 weeks | Promoted cancer | Mice | [88] |
| Gingko biloba extract | 0–1000 mg/kg b.w., 5 days per week for 14 weeks. | Promoted Cancer | Mice | [88] |
| Isothiocyanates | 0.1% of diet | Promoted cancer | Mice | [89] |
| Annexin A2-conjugated curcumin loaded PLGA nanoparticles. | 0–80 µM | Inhibit angiogenesis and cancer cell survival | Breast cancer cell lines | [91] |
| Resveratrol-loaded nanoparticles | 100–300 µM | Inhibit metastasis and regulate redox homeostasis | Mice | [92] |
| DMSA conjugated Apigenin nanoparticles | 0–16 µg/mL; 5 mg/kg b.w. | Increased bioavailability and anticancer effect | Lung cancer, B16F10 and A549; Mice | [93] |
| Quercetin loaded chitosan nanoparticles | 12.5–200 μM; 25 mg/kg b.w. | Reduce tumor volume and increase the antioxidant level | Lung cancer, A549; breast cancer, MDA MB 468; Mice | [94] |
| Nanoemulsion of Rutin | 30–300 μM; 20–300 μM; 50–300 μM | Increased bioavailability and anticancer effect | Lung cancer, A549; Colon cancer, Caco-2 human fibroblast cells, respectively | [95] |
| Rutin loaded-PCL-PEG and PLGA nanoparticles | 5–50 mg/kg b.w. | Suppress oxidative stress | Rat | [96] |
| Rutin loaded PCL-PEG nanoparticles | 0–60 μM | Suppress oxidative stress | Human ovarian cancer, Skov3 | [97] |
| Betulinic acid loaded PLGA nanoparticles | 10–80 µg/mL; 100 mg/kg b.w. | Balance redox homeostasis | Hep-G2 cells; Wistar rats |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatma, H.; Jameel, M.; Siddique, H.R. An Update on Phytochemicals in Redox Homeostasis: “Virtuous or Evil” in Cancer Chemoprevention? Chemistry 2023, 5, 201-222. https://doi.org/10.3390/chemistry5010017
Fatma H, Jameel M, Siddique HR. An Update on Phytochemicals in Redox Homeostasis: “Virtuous or Evil” in Cancer Chemoprevention? Chemistry. 2023; 5(1):201-222. https://doi.org/10.3390/chemistry5010017
Chicago/Turabian StyleFatma, Homa, Mohd Jameel, and Hifzur R. Siddique. 2023. "An Update on Phytochemicals in Redox Homeostasis: “Virtuous or Evil” in Cancer Chemoprevention?" Chemistry 5, no. 1: 201-222. https://doi.org/10.3390/chemistry5010017
APA StyleFatma, H., Jameel, M., & Siddique, H. R. (2023). An Update on Phytochemicals in Redox Homeostasis: “Virtuous or Evil” in Cancer Chemoprevention? Chemistry, 5(1), 201-222. https://doi.org/10.3390/chemistry5010017