

Bacteriocins and Bacteriophages as Dual Biological Players for Food Safety Applications




Definition
:1. Introduction
2. Bacteriocins
3. Bacteriocins in the Food Industry

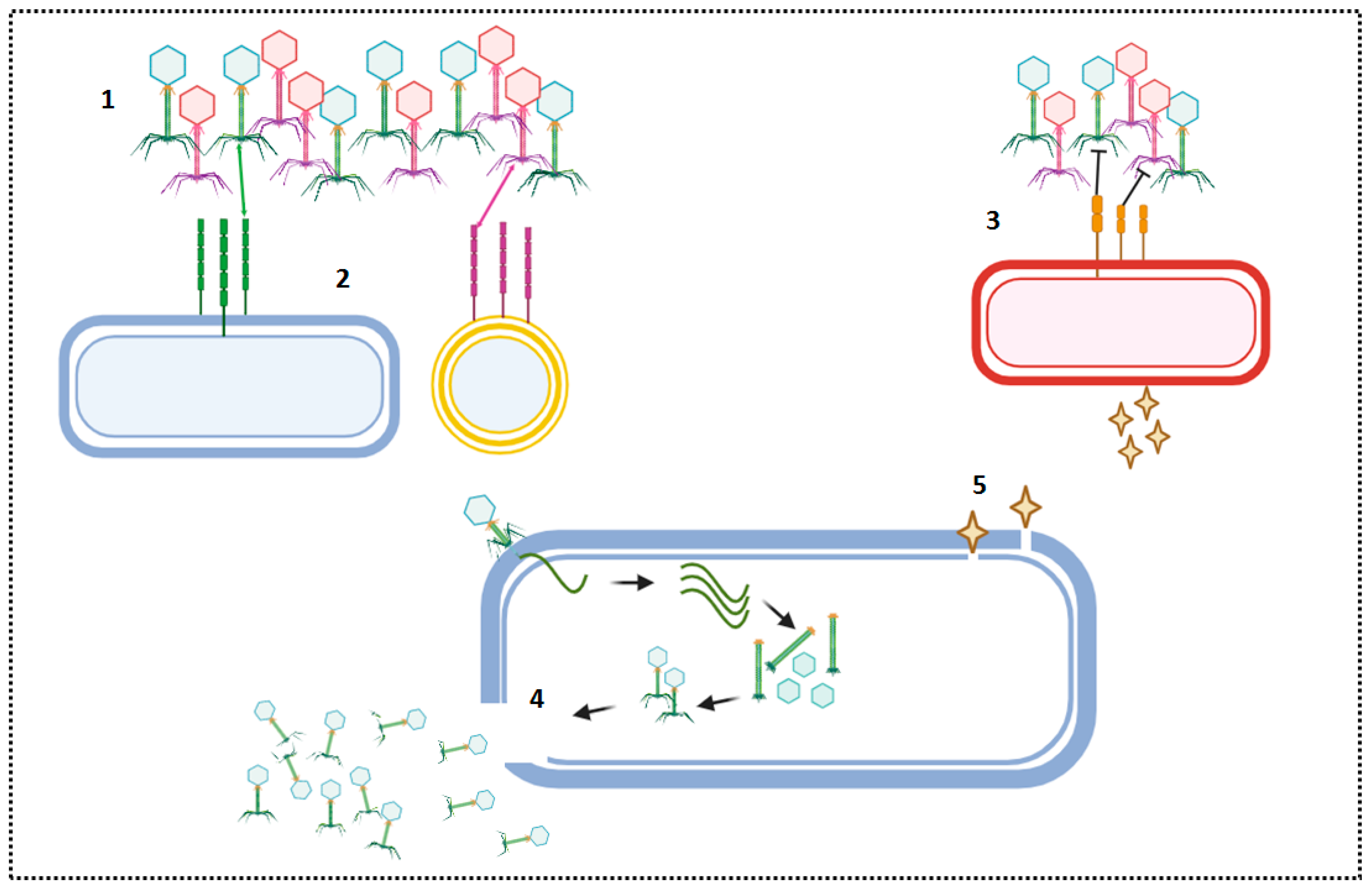{kind=link}
| Producing Strain | Bacteriocin | Application | Target Microorganism | Reference |
|---|---|---|---|---|
| Pediocooccus acidilactici PAC1.0 | Pediocin PA-1/AcH | Raw chicken | Listeria monocytogenes | [54] |
| Latilactobacillus sakei Lb674 | Sakacin P | Chicken | Listeria monocytogenes | [55] |
| Enterococcus faecium CTC492 | Enterocins A and B | Ham | Listeria monocytogenes, Lactobacillus sakei | [56] |
| Enterococcus faecium L50 | Enterocin L50A/L50B | Beers | Lactobacillus brevis, Pediococcus damnosus | [57] |
| Enterococcus faecium FL31 | Bacteriocin BacFL31 | Ground turkey meet | Listeria monocytogenes Salmonella Typhimurium | [58] |
| Lactococcus lactis CSK3533 Lactiplantibacillus plantarum LMG P-26358 | Nisin/Lacticin 3147/Plantaricin | Cheese | Listeria monocytogenes | [59] |
| Lacticaseibacillus rhamnosus RC20975 | Bacteriocin RC20975 | Apple juice | Alicyclobacillus acidoterrestris | [60] |
| Lactiplantibacillus plantarum BM-1 | Plantaricin BM-1 | Fresh pork meat | Listeria monocytogenes | [61] |
| Lactobacillus crustorum MN047 | Bacteriocin BM1300 | Beef meat | Staphylococcus aureus Escherichia coli | [62] |
| Lactiplantibacillus plantarum S7-10 | Plantaricin E and F | Honeybee | Paenibacillus larvae | [63] |
| Lactococcus lactis CAU2013 | Fresh cheese | Listeria monocytogenes | [38] | |
| Lactococcus lactis F01 | bacteriocin peptide C4B | Fish sausage | Clostridium sp., Staphylococcus aureus, Bacillus subtilis, Salmonella Typhimurium, Pseudomonas aeruginosa, Klebsiella pneumoniae and Escherichia coli | [64] |
| Lactiplantibacillus plantarum | Table grapes | Pseudomonas syringae pv. syringae and Botrytis cinerea | [65] | |
| Limosilactobacillus panis C-M2 | Lactocin C-M2 | Fish | Staphylococcus aureus | [66] |
| Enterococcus faecalis L2B21K3 and L3A21K6 | Ripened cheese | Listeria monocytogenes | [67] |
4. Bacteriophages
5. Bacteriophages in the Food Sector
| Bacteriophages | Application | Target Microorganism | Reference |
|---|---|---|---|
| Phage BPECO19 | Chicken meat | Escherichia coli O157:H7 | [109] |
| Phage A511 | Cooked meat | Listeria monocytogenes | [110] |
| Phage Akh-2 | Aquaculture | Aeromonas hydrophila | [111] |
| Phages LSA2308 and LSA2366 (alone or as cocktail) | Milk production chain | Staphylococcus aureus | [112] |
| Phage T156 | Milk | Salmonella Typhimurium | [113] |
| Phages SE-P3, P16, P37 and P47 (alone or as cocktail) | Raw chicken meat | Salmonella enteritidis | [114] |
| Cocktail phages (C14 s, L1, V9 and LL15) | Fruits and vegetables | Escherichia coli O157:H7 | [115] |
| Phage A511 | Cheese | Listeria monocytogenes | [116] |
| PhageGuard ListexTM P100 | Fuet, cocked ham and fresh cheese | Listeria innocua | [117] |
6. The Combination of Bacteriophage and Bacteriocin: A Tool against Foodborne Pathogens
7. Conclusions
Funding
Conflicts of Interest
References
- Brauge, T.; Faille, C.; Leleu, G.; Denis, C.; Hani, A.; Midelet, G. Treatment with disinfectants may induce an increase in viable but non-culturable populations of Listeria monocytogenes in biofilms formed in smoked salmon processing environments. Food Microbiol. 2020, 92, 103548. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial Biofilms in the Food Industry—A Comprehensive Review. Int. J. Environ. Res. Public Health 2021, 18, 2014. [Google Scholar] [CrossRef] [PubMed]
- Wagh, R.V.; Priyadarshi, R.; Rhim, J.-W. Novel Bacteriophage-Based Food Packaging: An Innovative Food Safety Approach. Coatings 2023, 13, 609. [Google Scholar] [CrossRef]
- Rendueles, C.; Duarte, A.-C.; Escobedo, S.; Fernández, L.; Rodríguez, A.; García, P.; Martínez, B. Combined use of bacteriocins and bacteriophages as food biopreservatives. A review. Int. J. Food Microbiol. 2022, 368, 109611. [Google Scholar] [CrossRef] [PubMed]
- Muthuvelu, K.-S.; Ethiraj, B.; Pramnik, S.; Raj, N.-K.; Venkataraman, S.; Rajendran, D.-S.; Bharathi, P.; Palanisamy, E.; Narayanan, A.S.; Vaidyanathan, V.K.; et al. Biopreservative technologies of food: An alternative to chemical preservation and recent developments. Food Sci. Biotechnol. 2023, 32, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Drider, D.; Rebuffat, S. Prokaryotic Antimicrobial Peptides: From Genes to Applications; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Whitehead, H.-R. A substance inhibiting bacterial growth, produced by certain strains of lactic streptococci. Biochem. J. 1933, 27, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Jack, R.-W.; Tagg, J.R.; Ray, B. Bacteriocins of gram-positive bacteria. Microbiol. Mol. Biol. Rev. 1995, 59, 171–200. [Google Scholar] [CrossRef]
- Perez, R.-H.; Zendo, T.; Sonomoto, K. Multiple bacteriocin production in lactic acid bacteria. J. Biosci. Bioeng. 2022, 134, 277–287. [Google Scholar] [CrossRef]
- Gough, R.; O’Connor, P.-M.; Rea, M.-C.; Gómez-Sala, B.; Miao, S.; Hill, C.; Brodkorb, A. Simulated gastrointestinal digestion of nisin and interaction between nisin and bile. Lebensm.-Wiss. Technol. 2017, 86, 530–537. [Google Scholar] [CrossRef]
- Klaenhammer, T.-R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef]
- Arnison, P.-G.; Bibb, M.-J.; Bierbaum, G.; Bowers, A.-A.; Bugni, T.-S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2012, 30, 108–160. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.-D.; Ross, R.-P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.-P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed]
- Acedo, J.A.; Chiorean, S.; Vederas, J.C.; van Belkum, M.J. The expanding structural variety among bacteriocins from Gram-positive bacteria. FEMS Microbiol. Rev. 2018, 42, 805–828. [Google Scholar]
- Soltani, S.; Hammami, R.; Cotter, P.-D.; Rebuffat, S.; Said, L.-B.; Gaudrea, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef]
- Perez, R.-H.; Zendo, T.; Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microb. Cell Factories 2014, 13, S3. [Google Scholar] [CrossRef]
- Gálvez, A.; Abriouel, H.; López, R.-L.; Omar, N.-B. Bacteriocin-based strategies for food biopreservation. Int. J. Food Microbiol. 2007, 120, 51–70. [Google Scholar]
- Stern, N.-J.; Svetoch, E.-A.; Eruslanov, B.-V.; Perelygin, V.-V.; Mitsevich, E.-V.; Mitsevich, I.-P.; Pokhilenko, V.D.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S. Isolation of a Lactobacillus salivarius Strain and Purification of Its Bacteriocin, Which Is Inhibitory to Campylobacter jejuni in the Chicken Gastrointestinal System. Antimicrob. Agents Chemother. 2006, 50, 3111–3116. [Google Scholar] [CrossRef]
- Line, J.E.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S.; Siragusa, G.R.; et al. Isolation and Purification of Enterocin E-760 with Broad Antimicrobial Activity against Gram-Positive and Gram-Negative Bacteria. Antimicro. Agents Chemother. 2008, 52, 1094–1100. [Google Scholar] [CrossRef]
- Messaoudi, S.; Madi, A.; Prévost, H.; Feuilloley, M.; Manai, M.; Dousset, X.; Connil, N. In Vitro evaluation of the probiotic potential of Lactobacillus salivarius SMXD51. Anaerobe 2012, 18, 584–589. [Google Scholar] [CrossRef]
- Song, D.-F.; Zhu, M.-Y.; Gu, Q. Purification and Characterization of Plantaricin ZJ5, a New Bacteriocin Produced by Lactobacillus plantarum ZJ5. PLoS ONE 2020, 9, e105549. [Google Scholar] [CrossRef] [PubMed]
- Madi-Moussa, D.; Deracinois, B.; Teiar, R.; Li, Y.; Mihasan, M.; Flahaut, C.; Rebuffat, S.; Coucheney, F.; Drider, D. Structure of Lacticaseicin 30 and Its Engineered Variants Revealed an Interplay between the N-Terminal and C-Terminal Regions in the Activity against Gram-Negative Bacteria. Pharmaceutics 2022, 14, 1921. [Google Scholar] [CrossRef] [PubMed]
- Prudêncio, C.-V.; dos Santos, M.-T.; Vanetti, M.-C.-D. Strategies for the use of bacteriocins in Gram-negative bacteria: Relevance in food microbiology. J. Food Sci. Technol. 2015, 52, 5408–5417. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ramos, A.; Madi-Moussa, D.; Coucheney, F.; Drider, D. Current Knowledge of the Mode of Action and Immunity Mechanisms of LAB-Bacteriocins. Microorganisms 2021, 9, 2107. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, D.; Nag, M.; Dutta, B.; Sarkar, T.; Pati, S.; Basu, D.; Kari, Z.A.; Wei, L.S.; Smaoui, S.; Goh, K.W.; et al. Bacteriocin: A natural approach for food safety and food security. Front. Bioeng. Biotechnol. 2022, 10, 1005918. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.-C.; Todorov, S.-D.; Chihib, N.-E.; Drider, D.; Nero, L.-A. Lactic Acid Bacteria (LAB) and Their Bacteriocins as Alternative Biotechnological Tools to Control Listeria monocytogenes Biofilms in Food Processing Facilities. Mol. Biotechnol. 2018, 60, 712–726. [Google Scholar] [CrossRef]
- Kirtonia, K.; Salauddin, M.; Bharadwaj, K.-K.; Pati, S.; Dey, A.; Shariati, M.-A.; Tilak, V.K.; Kuznetsova, E.; Sarkar, T. Bacteriocin: A new strategic antibiofilm agent in food industries. Biocat. Agric. Biotechnol. 2021, 36, 102141. [Google Scholar] [CrossRef]
- Castellano, P.; Pérez-Ibarreche, M.; Blanco-Massani, M.; Fontana, C.; Vignolo, G.-M. Strategies for Pathogen Biocontrol Using Lactic Acid Bacteria and Their Metabolites: A Focus on Meat Ecosystems and Industrial Environments. Microorganisms 2017, 5, 38. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.-K.; Rajput, Y.-S.; Sood, S.-K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Nanda, P.-K.; Pateiro, M.; Lorenzo, J.-M.; Dhar, P.; Das, A.-K. Lactic Acid Bacteria and Bacteriocins: Novel Biotechnological Approach for Biopreservation of Meat and Meat Products. Microorganisms 2022, 10, 2058. [Google Scholar] [CrossRef]
- Borges, F.; Briandet, R.; Callon, C.; Champomier-Vergès, M.-C.; Christieans, S.; Chuzeville, S.; Denis, C.; Desmasures, N.; Desmonts, M.-H.; Feurer, C.; et al. Contribution of omics to biopreservation: Toward food microbiome engineering. Front. Microbiol. 2022, 13, 951182. [Google Scholar] [CrossRef] [PubMed]
- Todorov, S.-D.; Popov, I.; Weeks, R.; Chikindas, M.-L. Use of Bacteriocins and Bacteriocinogenic Beneficial Organisms in Food Products: Benefits, Challenges, Concerns. Foods 2022, 11, 3145. [Google Scholar] [CrossRef] [PubMed]
- Speranza, B.; Corbo, M.-R. Chapter 11—The Impact of Biofilms on Food Spoilage. In The Microbiological Quality of Food; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 259–282. [Google Scholar]
- Speranza, B.; Liso, A.; Russo, V.; Corbo, M.-R. Evaluation of the Potential of Biofilm Formation of Bifidobacterium longum subsp. infantis and Lactobacillus reuteri as Competitive Biocontrol Agents Against Pathogenic and Food Spoilage Bacteria. Microorganisms 2020, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Song, X.; Chen, M.; Tian, S.; Lu, Z.; Sun, J.; Li, X.; Lu, Y.; Yuk, H. Combating biofilms of foodborne pathogens with bacteriocins by lactic acid bacteria in the food industry. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1657–1676. [Google Scholar] [CrossRef]
- Richard, C.; Brillet, A.; Pilet, M.-F.; Prévost, H.; Drider, D. Evidence on inhibition of Listeria monocytogenes by divercin V41 action. Lett. Appl. Microbiol. 2003, 36, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Kim, G.-B. Inhibition of Listeria monocytogenes in Fresh Cheese Using a Bacteriocin-Producing Lactococcus lactis CAU2013 Strain. Food Sci. Anim. Resour. 2022, 42, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Jiang, H.; Cheng, H.; Fang, J.; Huang, G. Strategies for screening, purification and characterization of bacteriocins. Int. J. Biol. Macromol. 2018, 117, 781–789. [Google Scholar] [CrossRef]
- Lahiri, D.; Chakraborti, S.; Jasu, A.; Nag, M.; Dutta, B.; Dash, S.; Ray, R.R. Production and purification of bacteriocin from Leuconostoc lactis SM 2 strain. Biocat. Agric. Biotechnol. 2020, 30, 101845. [Google Scholar] [CrossRef]
- O’Connor, P.-M.; Kuniyoshi, T.-M.; Oliveira, R.-P.; Hill, C.; Ross, R.-P.; Cotter, P.-D. Antimicrobials for food and feed; a bacteriocin perspective. Curr. Opin. Biotechnol. 2020, 61, 160–167. [Google Scholar] [CrossRef]
- Shafique, B.; Ranjha, M.-M.-A.-N.; Murtaza, M.-A.; Walayat, N.; Nawaz, A.; Khalid, W.; Mahmood, S.; Nadeem, M.; Manzoor, M.F.; Ameer, K.; et al. Recent Trends and Applications of Nanoencapsulated Bacteriocins against Microbes in Food Quality and Safety. Microorganisms 2023, 11, 85. [Google Scholar] [CrossRef]
- Realini, C.-E.; Marcos, B. Active and intelligent packaging systems for a modern society. Meat Sci. 2014, 98, 404–419. [Google Scholar] [CrossRef] [PubMed]
- Daba, G.M.; Elkhateeb, W.A. Bacteriocins of lactic acid bacteria as biotechnological tools in food and pharmaceuticals: Current applications and future prospects. Biocatal. Agric. Biotechnol. 2020, 28, 101750. [Google Scholar] [CrossRef]
- Mokoena, M.-P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Hussien, H.; Abd-Rabou, H.-S.; Saad, M.-A. The impact of incorporating Lactobacillus acidophilus bacteriocin with inulin and FOS on yogurt quality. Sci. Rep. 2022, 12, 13401. [Google Scholar] [CrossRef] [PubMed]
- Bukvicki, D.; D’Alessandro, M.; Rossi, S.; Siroli, L.; Gottardi, D.; Braschi, G.; Patrignani, F.; Lanciotti, R. Essential Oils and Their Combination with Lactic Acid Bacteria and Bacteriocins to Improve the Safety and Shelf Life of Foods: A Review. Foods 2023, 12, 3288. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Dong, Q.; Song, X.; Xu, L.; Xia, X.; Aslam, M.-Z.; Ma, Y.; Qin, X.; Wang, X.; Liu, Y.; et al. Effective combination of nisin and sesamol against Listeria monocytogenes. LWT—Food Sci. Technol. 2023, 176, 114546. [Google Scholar] [CrossRef]
- Austrich-Comas, A.; Serra-Castelló, C.; Jofré, A.; Gou, P.; Bover-Cid, S. Control of Listeria monocytogenes in chicken dry-fermented sausages with bioprotective starter culture and high-pressure processing. Front. Microbiol. 2023, 13, 983265. [Google Scholar] [CrossRef]
- da Costa, R.-J.; Voloski, F.-L.-S.; Mondadori, R.-G.; Duval, E.-H.; Fiorentini, Â.-M. Preservation of Meat Products with Bacteriocins Produced by Lactic Acid Bacteria Isolated from Meat. J. Food Qual. 2019, 2019, e4726510. [Google Scholar] [CrossRef]
- Leverentz, B.; Conway, W.S.; Camp, M.J.; Janisiewicz, W.J.; Abuladze, T.; Yang, M.; Saftner, R.; Sulakvelidze, A. Biocontrol of Listeria monocytogenes on Fresh-Cut Produce by Treatment with Lytic Bacteriophages and a Bacteriocin. Appl. Environ. Microbiol. 2003, 69, 4519–4526. [Google Scholar] [CrossRef]
- Kim, S.-G.; Lee, Y.-D.; Park, J.-H.; Moon, G.-S. Synergistic Inhibition by Bacteriocin and Bacteriophage against Staphylococcus aureus. Food Sci. Anim. Resour. 2019, 39, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.-H.; Bhunia, A.-K.; Johnson, M.-G. Complete Inhibition of Low Levels of Listeria monocytogenes on Refrigerated Chicken Meat with Pediocin AcH Bound to Heat-Killed Pediococcus acidilactici Cells. J. Food Prot. 1996, 59, 1187–1192. [Google Scholar] [CrossRef]
- Katla, T.; Møretrø, T.; Sveen, I.; Aasen, I.M.; Axelsson, L.; Rørvik, L.M.; Naterstad, K. Inhibition of Listeria Monocytogenes in Chicken Cold Cuts by Addition of Sakacin P and Sakacin P-producing Lactobacillus Sakei. J. Appl. Microbiol. 2002, 93, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Marcos, B.; Aymerich, T.; Monfort, J.-M.; Garriga, M. High-pressure processing and antimicrobial biodegradable packaging to control Listeria monocytogenes during storage of cooked ham. Food Microbiol. 2008, 25, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Basanta, A.; Sánchez, J.; Gómez-Sala, B.; Herranz, C.; Hernández, P.-E.; Cintas, L.-M. Antimicrobial activity of Enterococcus faecium L50, a strain producing enterocins L50 (L50A and L50B), P and Q, against beer-spoilage lactic acid bacteria in broth, wort (hopped and unhopped), and alcoholic and non-alcoholic lager beers. Int. J. Food Microbiol. 2008, 125, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Chakchouk-Mtibaa, A.; Smaoui, S.; Ktari, N.; Sellem, I.; Najah, S.; Karray-Rebai, I.; Mellouli, L. Biopreservative Efficacy of Bacteriocin BacFL31 in Raw Ground Turkey Meat in terms of Microbiological, Physicochemical, and Sensory Qualities. Biocontrol Sci. 2017, 22, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; Griffin, C.; O’Connor, P.-M.; Serrano, L.-M.; Meijer, W.C.; Hill, C.; Ross, R.P. A Multibacteriocin Cheese Starter System, Comprising Nisin and Lacticin 3147 in Lactococcus lactis, in Combination with Plantaricin from Lactobacillus plantarum. Appl. Envrion. Microbiol. 2017, 83, e00799-17. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Yue, T.; Yuan, Y.; Dai, L. Activity of paracin C from lactic acid bacteria against Alicyclobacillus in apple juice: Application of a novelty bacteriocin. J. Food Saf. 2017, 37, e12350. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, M.; Gao, X.; Shao, Y.; Liu, H.; Jin, J.; Yang, W.; Zhang, H. Development and antimicrobial application of plantaricin BM-1 incorporating a PVDC film on fresh pork meat during cold storage. J. Appl. Microbiol. 2018, 125, 1108–1116. [Google Scholar] [CrossRef]
- Lu, Y.; Aizhan, R.; Yan, H.; Li, X.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Characterization, modes of action, and application of a novel broad-spectrum bacteriocin BM1300 produced by Lactobacillus crustorum MN047. Braz. J. Microbiol. 2020, 51, 2033–2048. [Google Scholar] [CrossRef]
- Lazzeri, A.-M.; Mangia, N.-P.; Mura, M.-E.; Floris, I.; Satta, A.; Ruiu, L. Potential of novel food-borne Lactobacillus isolates against the honeybee pathogen Paenibacillus larvae. Biocontrol. Sci. Technol. 2020, 30, 897–908. [Google Scholar] [CrossRef]
- Fotso Techeu, U.-D.; Kaktcham, P.-M.; Momo, H.-K.; Foko Kouam, E.-M.; Tchamani-Piame, L.; Ngouenam, R.-J.; Zambou Ngoufack, F. Isolation, Characterization, and Effect on Biofilm Formation of Bacteriocin Produced by Lactococcus lactis F01 Isolated from Cyprinus carpio and Application for Biopreservation of Fish Sausage. BioMed Res. Int. 2022, 2022, e8437926. [Google Scholar] [CrossRef] [PubMed]
- Petkova, M.; Gotcheva, V.; Dimova, M.; Bartkiene, E.; Rocha, J.-M.; Angelov, A. Screening of Lactiplantibacillus plantarum Strains from Sourdoughs for Biosuppression of Pseudomonas syringae pv. syringae and Botrytis cinerea in Table Grapes. Microorganisms 2022, 10, 2094. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Wu, H.; Zhu, Y.; Zhou, J.; Yan, W.; Jianhao, Z.; Liu, X. Preservative effects of a novel bacteriocin from Lactobacillus panis C-M2 combined with dielectric barrier discharged cold plasma (DBD-CP) on acquatic foods. Food Sci. Technol. Int. 2023, 29, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.P.M.; Teixeira, J.A.; Silva, C.C.G. Application of enterocin-whey films to reduce Listeria monocytogenes contamination on ripened cheese. Food Microbiol. 2023, 109, 104134. [Google Scholar] [CrossRef]
- d’Herelle, F. An invisible microbe that is antagonistic to the dysentery bacillus. CR Acad. Sci. 1917, 165, 373–375. [Google Scholar]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.-G. Bacteriophage Therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef]
- Atterbury, R.-J. Bacteriophage biocontrol in animals and meat products. Microb. Biotechnol. 2009, 2, 601–612. [Google Scholar] [CrossRef]
- Fernández, L.; Gutiérrez, D.; Rodríguez, A.; García, P. Application of Bacteriophages in the Agro-Food Sector: A Long Way Toward Approval. Front. Cell Infect. Microbiol. 2018, 8, 296. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Bolton, D.; McAuliffe, O.; Coffey, A. Bacteriophages in Food Applications: From Foe to Friend. Annu. Rev. Food Sci. Technol. 2019, 10, 151–172. [Google Scholar] [CrossRef]
- Gordillo-Altamirano, F.-L.; Barr, J.-J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, R.-W.; Smith, M.-C.-M.; Burns, R.-N.; Ford, M.-E.; Hatfull, G.-F. Evolutionary relationships among diverse bacteriophages and prophages: All the world’s a phage. Proc. Natl. Acad. Sci. USA 1999, 96, 2192–2197. [Google Scholar] [CrossRef]
- Ackermann, H.-W. Bacteriophage taxonomy. Microbiology 2011, 32, 90–94. [Google Scholar] [CrossRef]
- Turner, D.; Kropinski, A.-M.; Adriaenssens, E.-M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; Ross, R.-P.; Hill, C. Bacteriocins and bacteriophage; a narrow-minded approach to food and gut microbiology. FEMS Microbiol. Rev. 2017, 41 (Suppl. 1), S129–S153. [Google Scholar] [CrossRef]
- AL-Ishaq, R.-K.; Skariah, S.; Büsselberg, D. Bacteriophage Treatment: Critical Evaluation of Its Application on World Health Organization Priority Pathogens. Viruses 2021, 13, 51. [Google Scholar] [CrossRef]
- Wernicki, A.; Nowaczek, A.; Urban-Chmiel, R. Bacteriophage therapy to combat bacterial infections in poultry. Virol. J. 2017, 14, 179. [Google Scholar] [CrossRef]
- Fister, S.; Robben, C.; Witte, A.-K.; Schoder, D.; Wagner, M.; Rossmanith, P. Influence of Environmental Factors on Phage–Bacteria Interaction and on the Efficacy and Infectivity of Phage P100. Front. Microbiol. 2016, 7, 1152. [Google Scholar] [CrossRef]
- Nabergoj, D.; Modic, P.; Podgornik, A. Effect of bacterial growth rate on bacteriophage population growth rate. MicrobiologyOpen 2018, 7, e00558. [Google Scholar] [CrossRef]
- Fernández, L.; Escobedo, S.; Gutiérrez, D.; Portilla, S.; Martínez, B.; García, P.; Rodríguez, A. Bacteriophages in the Dairy Environment: From Enemies to Allies. Antibiotics 2017, 6, 27. [Google Scholar] [CrossRef]
- Kazi, M.; Annapure, U.-S. Bacteriophage biocontrol of foodborne pathogens. J. Food Sci. Technol. 2016, 53, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Moye, Z.-D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Hyla, K.; Dusza, I.; Skaradzińska, A. Recent Advances in the Application of Bacteriophages against Common Foodborne Pathogens. Antibiotics 2022, 11, 536. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Yoo, D.-S.; Chang, Y.; Kim, S.-Y.; Han, J. Polycaprolactone film functionalized with bacteriophage T4 promotes antibacterial activity of food packaging toward Escherichia coli. Food Chem. 2021, 346, 128883. [Google Scholar] [CrossRef] [PubMed]
- Kering, K.-K.; Kibii, B.-J.; Wei, H. Biocontrol of phytobacteria with bacteriophage cocktails. Pest. Manag. Sci. 2019, 75, 1775–1781. [Google Scholar] [CrossRef]
- Vu, N.-T.; Oh, C.-S. Bacteriophage Usage for Bacterial Disease Management and Diagnosis in Plants. Plant Pathol. J. 2020, 36, 204–217. [Google Scholar] [CrossRef]
- Jassim, S.-A.-A.; Limoges, R.-G.; El-Cheikh, H. Bacteriophage biocontrol in wastewater treatment. World J. Microbiol. Biotechnol. 2016, 32, 70. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Bolton, D.; McAuliffe, O.; Coffey, A. The use of bacteriophages to control and detect pathogens in the dairy industry. Int. J. Dairy Technol. 2020, 73, 1–11. [Google Scholar] [CrossRef]
- Falardeau, J.; Trmčić, A.; Wang, S. The occurrence, growth, and biocontrol of Listeria monocytogenes in fresh and surface-ripened soft and semisoft cheeses. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4019–4048. [Google Scholar] [CrossRef]
- Nasr-Eldin, M.A.; Gamal, E.; Hazz, A.M.; Abo-Elmaaty, S.A. Isolation, characterization, and application of lytic bacteriophages for controlling Enterobacter cloacae complex (ECC) in pasteurized milk and yogurt. Folia Microbiol. 2023, 68, 911–924. [Google Scholar] [CrossRef]
- Axelsson, L.; Bjerke, G.A.; McLeod, A.; Berget, I.; Holck, A.L. Growth Behavior of Listeria monocytogenes in a Traditional Norwegian Fermented Fish Product (Rakfisk), and Its Inhibition through Bacteriophage Addition. Foods 2020, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Komora, N.; Maciel, C.; Amaral, R.A.; Fernandes, R.; Castro, S.M.; Saraiva, J.A.; Teixeira, P. Innovative hurdle system towards Listeria monocytogenes inactivation in a fermented meat sausage model—High pressure processing assisted by bacteriophage P100 and bacteriocinogenic Pediococcus acidilactici. Food Res. Int. 2021, 148, 110628. [Google Scholar] [CrossRef] [PubMed]
- Parsons, C.; Brown, P.; Kathariou, S. Use of Bacteriophage Amended with CRISPR-Cas Systems to Combat Antimicrobial Resistance in the Bacterial Foodborne Pathogen Listeria monocytogenes. Antibiotics 2021, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, R.R.; Trinh, J.T.; Bomholtz, L.; Brok-Lauridsen, S.K.; Sulakvelidze, A.; Nielsen, D.S. Bacteriophage Cocktail Significantly Reduces Listeria monocytogenes without Deleterious Impact on the Commensal Gut Microbiota under Simulated Gastrointestinal Conditions. Viruses 2022, 14, 190. [Google Scholar] [CrossRef] [PubMed]
- Strydom, A.; Witthuhn, C.R. Listeria monocytogenes: A Target for Bacteriophage Biocontrol. Compr. Rev. Food Sci. Food Saf. 2015, 14, 694–704. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Capita, R.; García-Fernández, C.; Alonso-Calleja, C. Effects of Bacteriophage P100 at Different Concentrations on the Structural Parameters of Listeria monocytogenes Biofilms. J. Food Prot. 2018, 81, 2040–2044. [Google Scholar] [CrossRef]
- Yan, J.; Yang, R.; Yu, S.; Zhao, W. The strategy of biopreservation of meat product against MRSA using lytic domain of lysin from Staphylococcus aureus bacteriophage. Food Biosci. 2021, 41, 100967. [Google Scholar] [CrossRef]
- Kwak, H.; Kim, J.; Ryu, S.; Bai, J. Characterization of KMSP1, a newly isolated virulent bacteriophage infecting Staphylococcus aureus, and its application to dairy products. Int. J. Food Microbiol. 2023, 390, 110119. [Google Scholar] [CrossRef]
- Tabla, R.; Martínez, B.; Rebollo, J.E.; González, J.; Ramírez, M.R.; Roa, I.; Rodríguez, A.; García, P. Bacteriophage performance against Staphylococcus aureus in milk is improved by high hydrostatic pressure treatments. Int. J. Food Microbiol. 2012, 156, 209–213. [Google Scholar] [CrossRef]
- Maje, M.D.; Kaptchouang Tchatchouang, C.D.; Manganyi, M.C.; Fri, J.; Ateba, C.N. Characterisation of Vibrio Species from Surface and Drinking Water Sources and Assessment of Biocontrol Potentials of Their Bacteriophages. Int. J. Microbiol. 2020, 2020, e8863370. [Google Scholar] [CrossRef]
- Wang, C.; Hang, H.; Zhou, S.; Niu, Y.D.; Du, H.; Stanford, K.; McAllister, T.A. Bacteriophage biocontrol of Shiga toxigenic Escherichia coli (STEC) O145 biofilms on stainless steel reduces the contamination of beef. Food Microbiol. 2020, 92, 103572. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, H.; Ryu, S. Bacteriophage and endolysin engineering for biocontrol of food pathogens/pathogens in the food: Recent advances and future trends. Crit. Rev. Food Sci. Nutr. 2022, 63, 8919–8938. [Google Scholar] [CrossRef] [PubMed]
- Ha, E.; Son, B.; Ryu, S. Clostridium perfringens Virulent Bacteriophage CPS2 and Its Thermostable Endolysin LysCPS2. Viruses 2018, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Połaska, M.; Sokołowska, B. Bacteriophages—A new hope or a huge problem in the food industry. AIMS Microbiol. 2019, 5, 324–346. [Google Scholar] [CrossRef]
- Samtlebe, M.; Ergin, F.; Wagner, N.; Neve, H.; Küçükçetin, A.; Franz, C.M.A.P.; Heller, K.J.; Hinrichs, J.; Atamer, Z. Carrier systems for bacteriophages to supplement food systems: Encapsulation and controlled release to modulate the human gut microbiota. LWT—Food Sci. Technol. 2016, 68, 334–340. [Google Scholar] [CrossRef]
- Endersen, L.; Coffey, A. The use of bacteriophages for food safety. Curr. Opin. Food Sci. 2020, 36, 1–8. [Google Scholar] [CrossRef]
- Seo, J.; Seo, D.J.; Oh, H.; Jeon, S.B.; Oh, M.H.; Choi, C. Inhibiting the Growth of Escherichia coli O157:H7 in Beef, Pork, and Chicken Meat using a Bacteriophage. Korean J. Food Sci. Anim. Resour. 2016, 36, 186–193. [Google Scholar] [CrossRef]
- Ahmadi, H.; Barbut, S.; Lim, L.T.; Balamurugan, S. Examination of the Use of Bacteriophage as an Additive and Determining Its Best Application Method to Control Listeria monocytogenes in a Cooked-Meat Model System. Front. Microbiol. 2020, 11, 779. [Google Scholar] [CrossRef]
- Akmal, M.; Rahimi-Midani, A.; Hafeez-ur-Rehman, M.; Hussain, A.; Choi, T.J. Isolation, Characterization, and Application of a Bacteriophage Infecting the Fish Pathogen Aeromonas hydrophila. Pathogens 2020, 9, 215. [Google Scholar] [CrossRef]
- Ma, F.; Ning, Y.; Wan, Q.; Zou, L.; Liu, Y.; Chen, S.; Li, J.; Zeng, Z.; Yang, Y.; Chen, H.; et al. Bacteriophages LSA2308 and LSA2366 infecting drug-resistant Staphylococcus aureus: Isolation, characterization and potential application for milk safety. LWT—Food Sci. Technol. 2021, 152, 112298. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Ding, Y.; Huang, C.; Zhang, Y.; Wang, J.; Wang, X. Characterization of a novel Siphoviridae Salmonella bacteriophage T156 and its microencapsulation application in food matrix. Food Res. Int. 2021, 140, 110004. [Google Scholar] [CrossRef] [PubMed]
- Aydin Demirarslan, Ö.; Alasalvar, H.; Yildirim, Z. Biocontrol of Salmonella Enteritidis on chicken meat and skin using lytic SE-P3, P16, P37, and P47 bacteriophages. LWT—Food Sci. Technol. 2021, 137, 110469. [Google Scholar] [CrossRef]
- Vengarai Jagannathan, B.; Kitchens, S.; Priyesh Vijayakumar, P.; Price, S.; Morgan, M. Efficacy of Bacteriophage Cocktail to Control, E. coli O157:H7 Contamination on Baby Spinach Leaves in the Presence or Absence of Organic Load. Microorganisms 2021, 9, 544. [Google Scholar] [CrossRef] [PubMed]
- García-Anaya, M.C.; Sepulveda, D.R.; Rios-Velasco, C.; Acosta-Muñiz, C.H. Incorporation of A511 bacteriophage in a whey protein isolate-based edible coating for the control of Listeria monocytogenes in Cheese. Food Packag. Shelf Life 2023, 37, 101095. [Google Scholar] [CrossRef]
- Colás-Medà, P.; Viñas, I.; Alegre, I. Evaluation of Commercial Anti-Listerial Products for Improvement of Food Safety in Ready-to-Eat Meat and Dairy Products. Antibiotics 2023, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Komora, N.; Maciel, C.; Pinto, C.A.; Ferreira, V.; Brandão, T.R.S.; Saraiva, J.M.A.; Castro, S.M.; Teixeira, P. Non-thermal approach to Listeria monocytogenes inactivation in milk: The combined effect of high pressure, pediocin PA-1 and bacteriophage P100. Food Microbiol. 2020, 86, 103315. [Google Scholar] [CrossRef]
- Baños, A.; García-López, J.D.; Núñez, C.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Biocontrol of Listeria monocytogenes in fish by enterocin AS-48 and Listeria lytic bacteriophage P100. LWT—Food Sci. Technol. 2016, 66, 672–677. [Google Scholar] [CrossRef]
- Heo, S.; Kim, M.G.; Kwon, M.; Lee, H.S.; Kim, G.B. Inhibition of Clostridium perfringens using Bacteriophages and Bacteriocin Producing Strains. Korean J. Food Sci. Anim. Resour. 2018, 38, 88–98. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barache, N.; Belguesmia, Y.; Martinez, B.; Seal, B.S.; Drider, D. Bacteriocins and Bacteriophages as Dual Biological Players for Food Safety Applications. Encyclopedia 2024, 4, 79-90. https://doi.org/10.3390/encyclopedia4010007
Barache N, Belguesmia Y, Martinez B, Seal BS, Drider D. Bacteriocins and Bacteriophages as Dual Biological Players for Food Safety Applications. Encyclopedia. 2024; 4(1):79-90. https://doi.org/10.3390/encyclopedia4010007
Chicago/Turabian StyleBarache, Nacim, Yanath Belguesmia, Beatriz Martinez, Bruce S. Seal, and Djamel Drider. 2024. "Bacteriocins and Bacteriophages as Dual Biological Players for Food Safety Applications" Encyclopedia 4, no. 1: 79-90. https://doi.org/10.3390/encyclopedia4010007
APA StyleBarache, N., Belguesmia, Y., Martinez, B., Seal, B. S., & Drider, D. (2024). Bacteriocins and Bacteriophages as Dual Biological Players for Food Safety Applications. Encyclopedia, 4(1), 79-90. https://doi.org/10.3390/encyclopedia4010007