

Saprophytic Filamentous Fungi against Helminths Affecting Captive Wild Animals


 , ,
, ,  ,
,  , and
, and
Definition
:1. Introduction
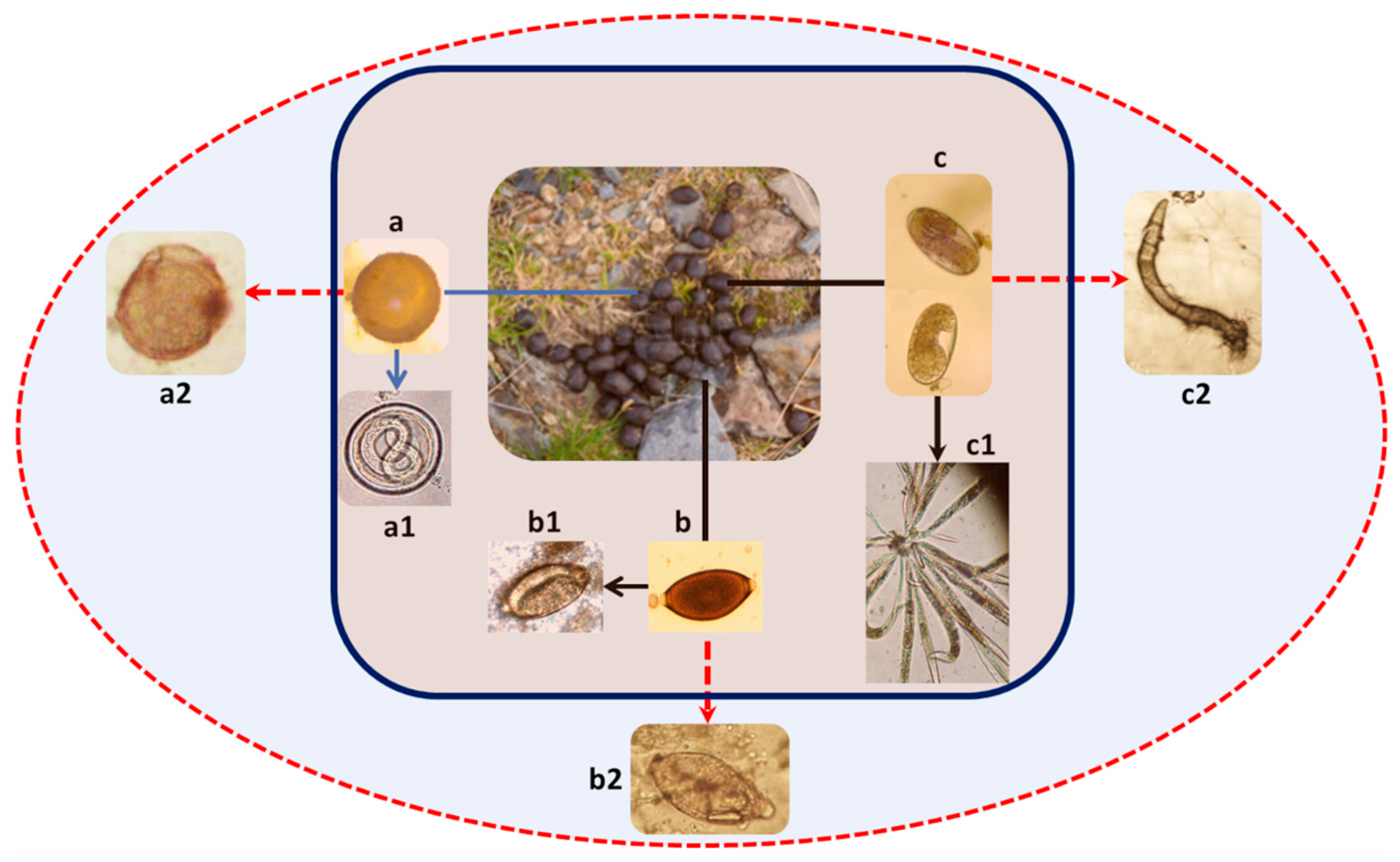
1.1. Main Parasites Affecting Captive-Bred Animals
1.1.1. Ascarids
1.1.2. Strongylids
1.1.3. Trichurids
1.1.4. Bronchopulmonary Nematodes
1.2. Clinical Importance and Detection
1.3. Control of Parasites in Captive Animals
- -
- In a large number of cases, deworming is carried out without a prior coprological examination.
- -
- The antiparasitic drugs used for livestock are often not specific for the target species, so they are underdosed to avoid intoxication problems.
- -
- Animal handling problems can cause most treatments to be administered orally due to the impossibility of giving them parenterally. This oral administration does not ensure an adequate ingestion of antiparasitic drugs, so there will be animals that ingest a higher dose than recommended and others in which the dosage is practically nil.
- -
- One of the main limiting factors regarding the implementation of preventive measures against parasites is the place where these animals are located, which is usually limited and permanent. The effectiveness of any antiparasitic treatment is reduced as a consequence of numerous reinfections when they are continuously located in the same plot.
- -
2. Application of Helmintophagous Fungi against Parasites Affecting Wild Captive Animals
- -
- Nematode-trapping fungi: These are characterized by producing an extensive system of hyphae or mycelium where, at certain intervals of development, they present specialized structures such as hyphal nets, knobs, branches, or rings to trap and sustain live nematodes (larval stages); therefore, they are ideal for the control of parasites such as strongyles [31]. Examples of this group are Duddingtonia flagrans, Arthrobotrys spp., and Monacrosporium thaumasium [32,33,34,35], which can develop different types of traps in which the mobile parasitic stages become caught, absorbed, and finally destroyed.
- -
- Predatory fungi or egg parasites: The hyphae of these fungi adhere to and penetrate the egg through small pores in the vitelline layer, causing permeability alterations. These fungi continue their development until they colonize the contents of the egg (embryo or larva). Good examples of this type of fungus are Pochonia chlamydosporia and Mucor circinelloides [36,37,38]. The parasiticide activity of these fungi is classified into three types, depending on how the egg morphology and viability are affected. Predatory fungi exhibit parasiticide activity type 1 when the hyphae surround the eggshell without penetrating it and where no embryo alterations are observed, but parasite development is stopped; activity type 2 occurs when the eggshell and embryo show morphological alterations without hyphae penetration; and, finally, type 3 activity takes place when the hyphae penetrate the eggshell and destroy the embryo so that the eggs become completely unviable, being unable to infect animals. If no damage or alterations are observable in the eggs, they remain viable, and this is considered activity type 0 [38].
- -
- Endoparasitic fungi: These fungi can infect nematodes through their spores. The fungi do not develop outside the host’s body and are not able to penetrate unless they are ingested, which makes their culture and maintenance in laboratory conditions very difficult and is the main reason for them not being used in current investigations.
2.1. Biological Control of Helminths Affecting Captive Animals: Analysis in Feces
2.2. Administration of Helmintophagous Fungi to Captive Animals
3. Conclusions and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Wolfe, B.A. Bovidae (except sheep and goats) and Antilocapridae. In Fowler’s Zoo and Wild Animal Medicine, 1st ed.; Miller, R.E., Fowler, M.E., Eds.; Elsevier: St. Louis, MO, USA, 2015; Volume 8, pp. 626–645. ISBN 978-1-4557-7397-8. [Google Scholar] [CrossRef]
- Palomero, A.M.; Cazapal-Monteiro, C.F.; Valderrábano, E.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Soil fungi enable the control of gastrointestinal nematodes in wild bovidae captive in a zoological park: A 4-year trial. Parasitology 2020, 147, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.-C.; Štrkolcová, G. Prevalence of endoparasites in carnivores in a zoo and a wolves park in Germany. Folia Vet. 2019, 63, 54–59. [Google Scholar] [CrossRef]
- Nath, T.C.; Eom, K.S.; Choe, S.; Hm, S.; Islam, S.; Ndosi, B.A.; Kang, Y.; Bia, M.M.; Kim, S.; Eamudomkarn, C.; et al. Insight into One Health approach: Endoparasite infections in captive wildlife in Bangladesh. Pathogens 2021, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, H.A. Anthelmintics resistance; how to overcome it? Iran. J. Parasitol. 2013, 8, 18–32. [Google Scholar]
- Young, K.E.; Jensen, J.M.; Craig, T.M. Evaluation of an anthelmintic activity in captive wild ruminants by fecal egg reduction tests and a larval development assay. J. Zoo Wildl. Med. 2000, 31, 348–352. [Google Scholar] [CrossRef]
- Garretson, P.D.; Hammond, E.E.; Craig, T.M.; Holman, P.J. Anthelmintic resistant Haemonchus contortus in a giraffe (Giraffa camelopardalis) in Florida. J. Zoo Wildl. Med. 2009, 40, 131–139. [Google Scholar] [CrossRef]
- Sengar, A.; Shrivastav, A.B.; Singh, K.P.; Rokde, A. Noninvasive assessment of gastrointestinal parasites infection in free-ranging wild herbivores and adjoining livestock of Panna Tiger Reserve, Madhya Pradesh, India. Vet. World 2017, 10, 748–751. [Google Scholar] [CrossRef]
- Atanaskova, E.; Kochevski, Z.; Stefanovska, J.; Nikolovski, G. Endoparasites in wild animals at the zoological garden in Skopje, Macedonia. JoTT Note 2011, 3, 1955–1958. [Google Scholar] [CrossRef]
- Mateus, T.L.; Castro, A.; Ribeiro, J.N.; Vieira-Pinto, M. Multiple zoonotic parasites identified in dog feces collected in Ponte de Lima, Portugal—A potential threat to human health. Int. J. Environ. Res. Public Health 2014, 11, 9050–9067. [Google Scholar] [CrossRef]
- Hoopes, J.; Hill, J.E.; Polley, L.; Fernando, C.; Wagner, B.; Schurer, J.; Jenkins, E. Enteric parasites of free-roaming, owned, and rural cats in prairie regions of Canada. Can. Vet. J. 2015, 56, 495–501. [Google Scholar]
- Côtes, E. Clinic and Veterinary Advisor. Dogs and Cats, 3rd ed.; Elsevier: Toronto, ON, Canada, 2014. [Google Scholar]
- Vandemark, L.M.; Jia, T.-W.; Zhou, X.-N. Social science implications for control of helminth infections in southeast Asia. Adv. Parasitol. 2010, 73, 137–170. [Google Scholar] [CrossRef] [PubMed]
- Elsheikha, H.M. Major cestodes infections. In Essentials of Veterinary Parasitology, 1st ed.; Elsheikha, H.M., Khan, N.A., Eds.; Caister Academic Press: Poole, UK, 2011; pp. 71–79. ISBN 978-1-904455-80-6. [Google Scholar]
- Maesano, G.; Capasso, M.; Ianniello, D.; Cringoli, G.; Rinaldi, L. Parasitic infections detected by FLOTAC in zoo mammals from Warsaw, Poland. Acta Parasitol. 2014, 59, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Gałęcki, R.; Sokół, R.; Koziatek, S. Parasites of wild animals as a potential source of hazard to humans. Ann. Parasitol. 2015, 61, 105–108. [Google Scholar] [PubMed]
- Voinot, M.; Bonilla, R.; Sousa, S.; Sanchís, J.; Canhão-Dias, M.; Romero Delgado, J.; Lozano, J.; Sánchez-Andrade, R.; Sol Arias, M.; Madeira de Carvalho, L. Control of Strongyles in First-Season Grazing Ewe Lambs by Integrating Deworming and Thrice-Weekly Administration of Parasiticidal Fungal Spores. Pathogens 2021, 10, 1338. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A.; Cazapal-Monteiro, C.F.; Arroyo, F.L.; Silva, M.I.; Palomero, A.M.; Paz-Silva, A.; Sanchez-Andrade, R.; Arias, M.S. Biological control of soil transmitted helminths (STHs) in a zoological park by using saprophytic fungi. Biol. Control 2018, 122, 24–30. [Google Scholar] [CrossRef]
- Barbosa, A.D.S.; Pinheiro, J.L.; Dos Santos, C.R.; de Lima, C.S.C.C.; Dib, L.V.; Echarte, G.V.; Augusto, A.M.; Bastos, A.C.M.P.; Antunes Uchôa, C.M.; Bastos, O.M.P.; et al. Gastrointestinal Parasites in Captive Animals at the Rio de Janeiro Zoo. Acta Parasitol. 2020, 6, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Saidi, A.; Mimouni, R.; Hamadi, F.; Oubrou, W. Coprological survey of protostrongylid infections in antelopes from Souss-Massa National Park (Morocco). Helminthologia 2020, 57, 306–313. [Google Scholar] [CrossRef]
- Panayotova-Pencheva, M.S. Species composition and morphology of protostrongylids (Nematoda: Protostrongylidae) in ruminants from Bulgaria. Parasitol. Res. 2011, 109, 1015–1020. [Google Scholar] [CrossRef]
- Panuska, C. Lungworms of ruminants. Vet. Clin. Food Anim. 2006, 22, 583–593. [Google Scholar] [CrossRef]
- Overgaauw, P.A.M.; van Knapen, F. Veterinary and public health aspects of Toxocara spp. Vet. Parasitol. 2013, 193, 398–403. [Google Scholar] [CrossRef]
- Goossens, E.; Dorny, P.; Boomker, J.; Vercammen, F.; Vercruysse, J. A 12-month survey of the gastro-intestinal helminths of antelopes, gazelles and giraffids kept at two zoos in Belgium. Vet. Parasitol. 2005, 127, 303–312. [Google Scholar] [CrossRef]
- Martínez, I.M.; Lumaret, J.-P.; Ortiz, R.Z.; Kadiri, N. The effects of sublethal and lethal doses of ivermectin on the reproductive physiology and larval development of the dung beetle Euoniticellus intermedius (Coleoptera: Scarabaeidae). Can. Entomol. 2017, 149, 461–472. [Google Scholar] [CrossRef]
- Verdú, J.R.; Cortez, V.; Ortiz, A.J.; Lumaret, J.-P.; Lobo, J.M.; Sánchez-Piñeiro, F. Biomagnification and body distribution of ivermectin in dung beetles. Sci. Rep. 2020, 10, 9073. [Google Scholar] [CrossRef] [PubMed]
- Fagiolini, M.; Lia, R.P.; Laricchiuta, P.; Cavicchio, P.; Mannella, R.; Cafarchia, C.; Otranto, D.; Finotello, R.; Perrucci, S. Gastrointestinal parasites in mammals of two Italian zoological gardens. J. Zoo Wildl. Med. 2010, 41, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Gavin, P.J.; Kazacos, K.R.; Shulman, S.T. Baylisascariasis. Clin. Microbiol. Rev. 2005, 18, 703–718. [Google Scholar] [CrossRef] [PubMed]
- Gawor, J.; Borecka, A. Quantifying the risk of zoonotic geohelminth infections for rural household inhabitants in Central Poland. Ann. Agric. Environ. Med. 2017, 24, 44–48. [Google Scholar] [CrossRef]
- Grønvold, J.; Henriksen, S.A.; Larsen, M.; Nansen, P.; Wolstrup, J. Biological control aspects of biological control—With special reference to arthropods, protozoans and helminths of domesticated animals. Vet. Parasitol. 1996, 64, 47–64. [Google Scholar] [CrossRef]
- Sagüés, M.F.; Purslow, P.; Fernández, S.; Fusé, L.; Iglesias, L.; Saumell, C. Nematophagous fungi used for the biological control of gastrointestinal nematodes in livestock and administration routes. Rev. Iberoam. Micol. 2011, 28, 143–147. [Google Scholar] [CrossRef]
- Maciel, A.S.; Freitas, L.G.; Figueiredo, L.D.; Campos, A.K.; Mello, I.N. Antagonistic activity of the fungus Pochonia clamydosporia on mature and immature Toxocara canis eggs. Parasitology 2012, 139, 1074–1085. [Google Scholar] [CrossRef]
- Arias, M.S.; Cazapal-Monteiro, C.F.; Suárez, J.; Miguélez, S.; Francisco, I.; Arroyo, F.L.; Suárez, J.L.; Paz-Silva, A.; Sánchez-Andrade, R.; Mendoza de Gives, P. Mixed production of filamentous fungal spores for preventing soil-transmitted helminth zoonoses: A preliminary analysis. Biomed. Res. Int. 2013, 2013, 567876. [Google Scholar] [CrossRef]
- Lozano, J.; Almeida, C.; Oliveira, M.; Paz-Silva, A.; Madeira de Carvalho, L. Biocontrol of avian gastrointestinal parasites using predatory fungi: Current status, challenges and opportunities. Parasitologia 2022, 2, 37–44. [Google Scholar] [CrossRef]
- Pérez-Anzúrez, G.; Olmedo-Juárez, A.; von-Son de Fernex, E.; Alonso-Díaz, M.A.; Delgado-Nuñez, E.J.; López-Arellano, M.E.; González-Cortázar, M.; Zamilpa, A.; Ocampo-Gutierrez, A.Y.; Paz-Silva, A.; et al. Arthrobotrys musiformis (Orbiliales) kills Haemonchus contortus infective larvae (Trichostrongylidae) through its predatory activity and its fungal culture infiltrates. Pathogens 2023, 11, 1068. [Google Scholar] [CrossRef] [PubMed]
- Lýsek, H.; Fassatiová, O.; Cuervo, N.P.; Lorenzo, N.H. Ovicidal fungi in soils of Cuba. Folia Parasitol. 1982, 29, 265–270. [Google Scholar] [PubMed]
- Carvalho, R.O.; Araújo, J.V.; Braga, F.R.; Araujo, J.M.; Alves, C.D. Ovicidal activity of Pochonia chlamydosporia and Paecilomyces lilacinus on Txocara canis eggs. Vet. Parasitol. 2010, 169, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Cazapal-Monteiro, C.F.; Hernández, J.A.; Arroyo, F.L.; Miguélez, S.; Romasanta, Á.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Analysis of the effect of soil saprophytic fungi on the eggs of Baylisascaris procyonis. Parasitol. Res. 2015, 114, 2443–2450. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.Á.; Cazapal-Monteiro, C.F.; Sanchís, J.; Sánchez-Andrade, R.; Paz-Silva, A.; Arias, M.S. Potential usefulness of filamentous fungi to prevent zoonotic soil-transmitted helminths. Vector Borne Zoonotic Dis. 2018, 18, 690–696. [Google Scholar] [CrossRef]
- Arroyo, F.L.; Arias, M.S.; Cazapal-Monteiro, C.F.; Hernández, J.A.; Suárez, J.; Miguélez, S.; Romasanta, A.; Sánchez-Andrade, R.; Paz-Silva, A. The capability of the fungus Mucor circinelloides to maintain parasiticidal activity after the industrial feed pelleting enhances the possibilities of biological control to livestock parasites. Biol. Control 2016, 92, 38–44. [Google Scholar] [CrossRef]
- Arias, M.S.; Arroyo, F.L.; Cazapal-Monteiro, C.F.; Hernández, J.A.; Suárez, J.; Francisco, I.; López-Arellano, M.E.; Sánchez-Andrade, R.; Mendoza de Gives, P.; Paz-Silva, A. Formulating Duddingtonia flagrans in nutritional pellets for the sustainable control of equine strongyles. J. Sci. Technol. Environ. 2015, 5, 3000249. [Google Scholar]
- Paraud, C.; Chartier, C. Biological control of infective larvae of a gastro-intestinal nematode (Teladorsagia circumcincta) and a small lungworm (Muellerius capillaris) by Duddingtonia flagrans in goat faeces. Parasitol. Res. 2003, 89, 102–106. [Google Scholar] [CrossRef]
- Paraud, C.; Cabaret, J.; Pors, I.; Chartier, C. Impact of the nematophagous fungus Duddingtonia flagrans on Muellerius capillaris larvae in goat faeces. Vet. Parasitol. 2005, 131, 71–78. [Google Scholar] [CrossRef]
- Canhão-Dias, M.; Paz-Silva, A.; Madeira de Carvalho, L.M. The efficacy of predatory fungi on the control of gastrointestinal parasites in domestic and wild animals-A systematic review. Vet. Parasitol. 2020, 283, 109173. [Google Scholar] [CrossRef] [PubMed]
- Terry, J.A. The Use of Duddingtonia flagrans for Gastrointestinal Parasitic Nematode Control in Feces of Exotic Artiodactylids at Disney’s Animal Kingdom. Master’s Thesis, Louisiana State University (LSU), Baton Rouge, LA, USA, 2013. [Google Scholar]
- Arias, M.; Cazapal-Monteiro, C.; Valderrábano, E.; Miguélez, S.; Rois, J.L.; López-Arellano, M.E.; Madeira de Carvalho, L.; Mendoza de Givez, P.; Sánchez-Andrade, R.; Paz-Silva, A. A preliminary study of biological control of strongyles affecting equids in a zoological park. J. Equine Vet. Sci. 2013, 33, 115–1120. [Google Scholar] [CrossRef]
- Palomero, A.M.; Hernández, J.A.; Cazapal-Monteiro, C.F.; Arroyo, F.B.; Silva, M.I.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Implementation of biological control to the integrated control of strongyle infection among wild captive equids in a zoological park. Biomed. Res. Int. 2018, 2018, 4267683. [Google Scholar] [CrossRef] [PubMed]
- Young, K.R. The Effect of the Nematode Trapping Fungus Duddingtonia flagrans against Gastronintestinal Nematodes of Exotic Ruminant Hoofstock at Disney’s ® Animal Kingdom Lodge. Master’s Thesis, Louisiana State University (LSU), Baton Rouge, LA, USA, 2018. [Google Scholar]


{kind=link}
| Helminth | Fungal Species | % Viability Reduction | Reference |
|---|---|---|---|
| Baylisascaris procyonis | Mucor circinelloides | 53–69% | [38] |
| Paecilomyces | 45–62% | ||
| Verticillium | 53–69% | ||
| Toxascaris leonina | Mucor circinelloides | 58% | [39] |
| Equine strongyle larvae | Duddingtonia flagrans | 84–89% | [41] |
| Parascaris equorum | Mucor circinelloides | 61–67% | [40] |
| Trichuris sp. | Mucor circinelloides | 11–50% | [18] |
| Trichoderma atrobrunnenum | 13–50% |
| Fungal Species | Formulation | Animal Species | Reference |
|---|---|---|---|
| Duddingtonia flagrans | Pellets top-dressed with a solution of chlamydospores | Giraffe Antelope Gerenuk | [45] |
| European donkey African ass Zebra | [46] | ||
| Powdered chlamydospores | Reticulated giraffe Scimitar-horned oryx Roan antelope | [48] | |
| M. circinelloides + D. flagrans | Nutritional pellets manufactured with fungal spores | Blackbuck Gazelle Mouflon Bison Marshbuck Kob | [47] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salmo, R.; Viña, C.; Lozano, J.; Palomero, A.M.; Hernández, J.Á.; Bonilla, R.; Sánchez-Andrade, R.; Paz-Silva, A.; Madeira de Carvalho, L.M.; Arias, M.S.; et al. Saprophytic Filamentous Fungi against Helminths Affecting Captive Wild Animals. Encyclopedia 2024, 4, 91-100. https://doi.org/10.3390/encyclopedia4010008
Salmo R, Viña C, Lozano J, Palomero AM, Hernández JÁ, Bonilla R, Sánchez-Andrade R, Paz-Silva A, Madeira de Carvalho LM, Arias MS, et al. Saprophytic Filamentous Fungi against Helminths Affecting Captive Wild Animals. Encyclopedia. 2024; 4(1):91-100. https://doi.org/10.3390/encyclopedia4010008
Chicago/Turabian StyleSalmo, Rami, Cándido Viña, João Lozano, Antonio M. Palomero, José Ángel Hernández, Rodrigo Bonilla, Rita Sánchez-Andrade, Adolfo Paz-Silva, Luis M. Madeira de Carvalho, María Sol Arias, and et al. 2024. "Saprophytic Filamentous Fungi against Helminths Affecting Captive Wild Animals" Encyclopedia 4, no. 1: 91-100. https://doi.org/10.3390/encyclopedia4010008
APA StyleSalmo, R., Viña, C., Lozano, J., Palomero, A. M., Hernández, J. Á., Bonilla, R., Sánchez-Andrade, R., Paz-Silva, A., Madeira de Carvalho, L. M., Arias, M. S., & Cazapal-Monteiro, C. (2024). Saprophytic Filamentous Fungi against Helminths Affecting Captive Wild Animals. Encyclopedia, 4(1), 91-100. https://doi.org/10.3390/encyclopedia4010008