
Suitability of Different Live Feed for First Feeding of Freshwater Fish Larvae

Abstract
:1. Introduction
2. Material and Methods
2.1. Institutional Review Board Statement
2.2. Species Involved in the Study
2.3. Live Food Used for Feeding of Fish Larvae
2.3.1. Collection of Lake Zooplankton
2.3.2. Microalgae Culture
2.3.3. Paramecium and Rotifer Culture
2.3.4. Artemia Culture
2.3.5. Morphometric Investigations
2.4. Larvae First Feeding Experiments
2.5. Fish Investigations
2.6. Statistics
3. Results
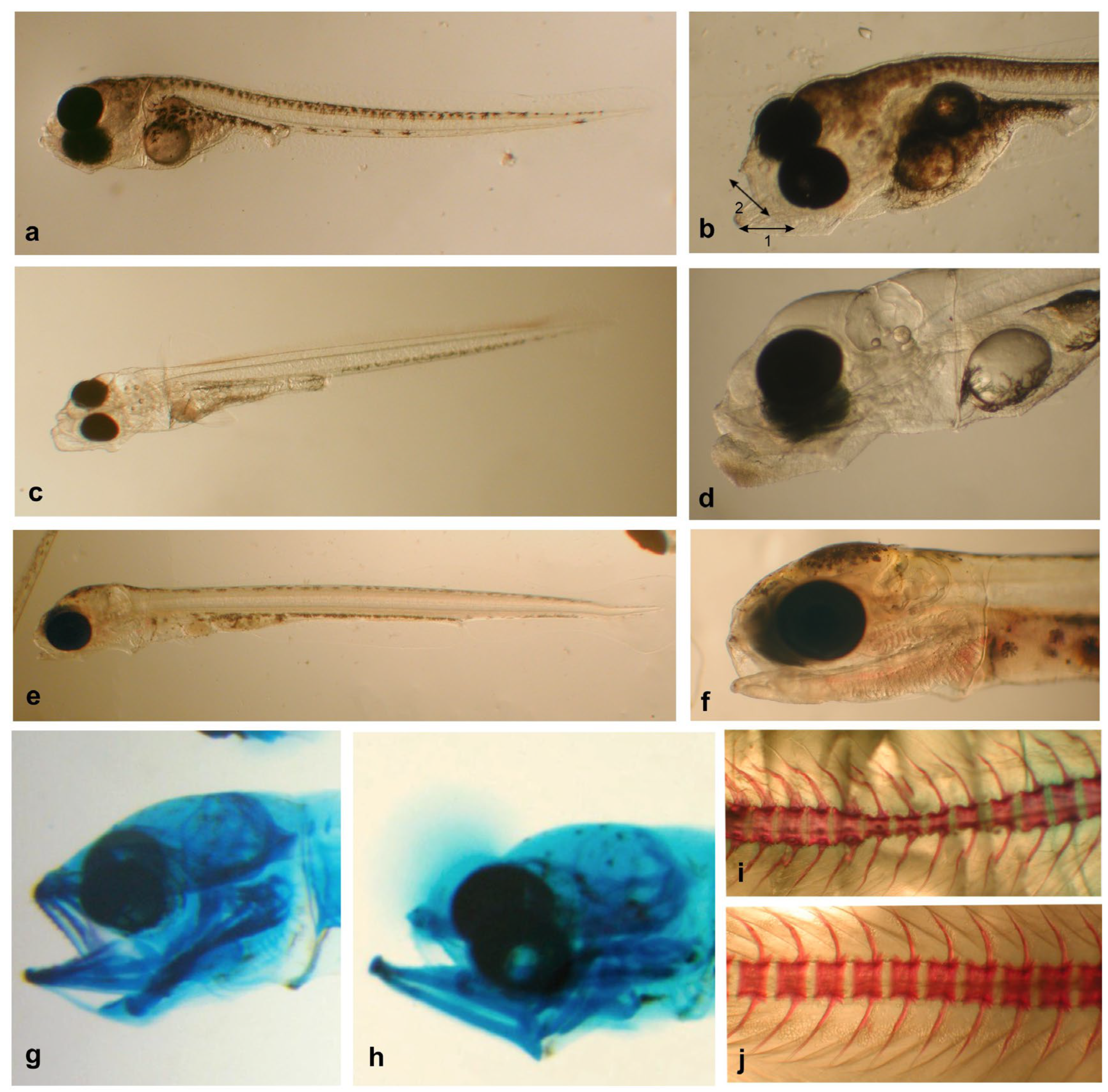
3.1. Morphometric Characterization Larvae
3.2. Morphometric Characterization of the Tested Live Feed
3.3. Feeding Trials
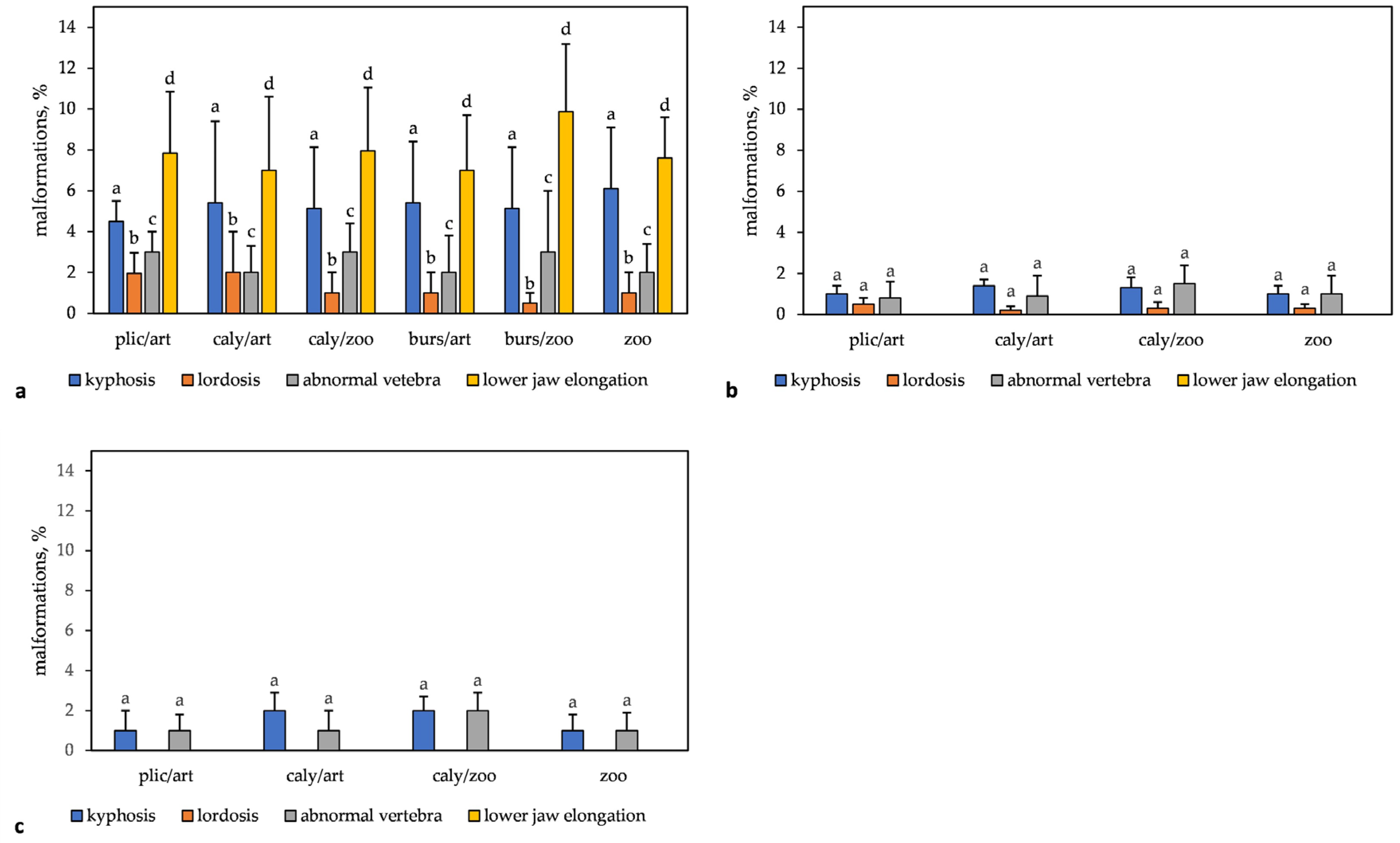
3.3.1. S. lucioperca
3.3.2. L. lota
3.3.3. Coregonus atterensis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tacon, A.G.J. Trends in global aquaculture and aquafeed production: 2000–2017. Rev. Fish. Sci. Aquacul. 2020, 28, 43–56. [Google Scholar] [CrossRef]
- Hamre, K.; Yufera, M.; Rønnestad, I.; Boglione, C.; Conceição, L.; Izquierdo, M. Fish larval nutrition and feed formulation: Knowledge gaps and bottlenecks for advances in larval rearing. Rev. Aquacult. 2013, 5, S26–S58. [Google Scholar] [CrossRef]
- Dhont, J.; Dierckens, K.; Støttrup, J.G.; Stappen, G.; Wille, M.; Sorgeloos, P. Rotifers, Artemia and copepods as live feeds for fish larvae in aquaculture. In Advances in Aquaculture Hatchery Technology; Allan, G., Burnell, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 157–202. [Google Scholar]
- Pan, Y.J.; Dahms, H.U.; Hwang, J.S.; Souissi, S. Recent trends in live feeds for marine larviculture: A mini review. Front. Mar. Sci. 2022, 9, 864165. [Google Scholar] [CrossRef]
- Kotani, T. Enrichment of rotifers and its effect on the growth and survival of fish larvae. In Rotifers; Hagiwara, A., Yoshinaga, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 47–62. [Google Scholar]
- Støttrup, J.G. Production and nutritional value of copepods. In Live Feeds in Marine Aquaculture; Støttrup, J.G., McEvoy, L.A., Eds.; Blackwell Science: Oxford, UK, 2013; pp. 145–205. [Google Scholar]
- Busch, K.E.T.; Peruzzi, S.; Tonning, F.; Falk-Petersen, I.B. Effect of prey type and size on the growth, survival and pigmentation of cod (Gadus morhua, L.) larvae. Aquacult. Nutr. 2011, 17, 595–603. [Google Scholar] [CrossRef]
- Harzevili, A.; De Charleroy, D.; Auwerx, J.; Vught, I.; Slycken, J.; Dhert, P.; Sorgeloos, P. Larval rearing of burbot (Lota lota L.) using Brachionus calyciflorus rotifer as starter food. J. App. Ichthyol. 2003, 19, 84–87. [Google Scholar] [CrossRef]
- Harzevili, A.S.; Dooremont, I.; Vught, I.; Auwerx, J.; Quataert, P.; De Charleroy, D. First feeding of burbot, Lota lota (Gadidae, Teleostei) larvae under different temperature and light conditions. Aquac. Res. 2004, 35, 49–55. [Google Scholar] [CrossRef]
- Bischoff-Lang, A.; Kubitz, M.; Wranik, C.; Ballesteros-Redondo, L.; Fink, P.; Palm, H. The effect of Brachionus calyciflorus (Rotifera) on larviculture and fatty acid composition of pikeperch (Sander lucioperca (L.) cultured under pseudo-green water conditions. Sustainability 2022, 14, 6607. [Google Scholar] [CrossRef]
- Imentai, A.; Yanes-Roca, C.; Steinbach, C.; Policar, T. Optimized application of rotifers Brachionus plicatilis for rearing pikeperch Sander lucioperca L. larvae. Aquacult. Internat. 2019, 27, 1137–1149. [Google Scholar] [CrossRef]
- Yanes-Roca, C.; Mráz, J.; Born-Torrijos, A.; Holzer, A.S.; Imentai, A.; Policar, T. Introduction of rotifers (Brachionus plicatilis) during pikeperch first feeding. Aquaculture 2018, 497, 260–268. [Google Scholar] [CrossRef]
- Mahmoudzadeh, H.; Ahmadi, M.; Shamsaei, M. Comparison of rotifer Brachionus plicatilis as a choice of live feed with dry feed in rearing Coregonus lavaretus fry. Aquacult. Nutr. 2009, 15, 129–134. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Kletzl, M. A method for rearing perch, Perca fluviatilis, larvae using Paramecium caudatum, followed by wild zooplankton and formulated dry feed in combination with adequate tank systems. J. Agricult. Sci. 2018, 10, 26–42. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Kletzl, M.; Weismann, T. Rearing of burbot, Lota lota (Pisces, Teleostei), larvae with zooplankton and formulated microdiets. J. Agricult. Sci. 2012, 4, 269–277. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Kletzl, M. On-feeding and juvenile production of coregonid species with formulated dry feeds: Effects on fish viability and digestive enzymes. J. Agricult. Sci. 2015, 7, 48–58. [Google Scholar] [CrossRef]
- Ballesteros-Redondo, L.; Palm, H.W.; Bährs, H.; Wacker, A.; Bischoff, A.A. Pikeperch larviculture (Sander lucioperca [L., 1758]) with Brachionus plicatilis (Mueller, 1786) (Rotifera) and Apocyclops panamensis (Marsh, 1913) (Copepoda). J. World Aquacult. Soc. 2023; early view. [Google Scholar]
- Imentai, A.; Rašković, B.; Steinbach, C.; Rahimnejad, S.; Yanes-Roca, C.; Policar, T. Effects of first feeding regime on growth performance, survival rate and development of digestive system in pikeperch (Sander lucioperca) larvae. Aquaculture 2020, 529, 735636. [Google Scholar] [CrossRef]
- Kottelat, M. European freshwater fishes. Env. Biol. Fish. 1998, 51, 116. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Husted, and Detonula confervacea (Cleve). Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Allen, M.M.; Stanier, R.Y. Growth and Division of Some Unicellular Blue-green Algae. J. Gen. Microbiol. 1968, 51, 199–202. [Google Scholar] [CrossRef]
- Hoshina, R. Size of Paramecium bursaria individuals under cold and dark conditions. Biologia 2014, 69, 1018–1022. [Google Scholar] [CrossRef]
- Suantika, G.; Dhert, P.; Nurhudah, M.; Sorgeloos, P. High-density production of the rotifer Brachionus plicatilis in a recirculation system: Consideration of water quality, zootechnical and nutritional aspects. Aquacult. Engin. 2000, 21, 201–213. [Google Scholar] [CrossRef]
- Park, H.; Lee, K.W.; Cho, S.; Kim, H.; Jung, M.M.; Kim, H.S. High density culture of the freshwater rotifer, Brachionus calyciflorus. Hydrobiologia 2001, 446, 369–374. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Meth. 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Wolnicki, J.; Leszek Myszkowski, L.; Kamiński, R. The influence of water temperature on the growth, survival, condition and biological quality of juvenile burbot, Lota lota (L.). Arch. Pol. Fish. 2001, 9, 79–86. [Google Scholar]
- Albert, A.; Vetemaa, M.; Saat, T. Effects of salinity on the development of Peipsi whitefish Coregonus lavaretus maraenoides Poljakow embryos. Annales Zool. Fenn. 2004, 41, 85–88. [Google Scholar]
- Walker, M.B.; Kimmel, C.B. A two-color acid-free cartilage and bone stain for zebrafish larvae. Biotech. Histochem. 2007, 82, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Boglione, C.; Gisbert, E.; Gavaia, P.; Witten, P.; Moren, M.; Fontagné-Dicharry, S.; Koumoundouros, G. Skeletal anomalies in reared European fish larvae and juveniles. Part 2: Main typologies, occurrences and causative factors. Rev. Aquacult. 2013, 5, 121–167. [Google Scholar] [CrossRef]
- Peterson, W.T. Patterns in stage duration and development among marine and freshwater calanoid and cyclopoid copepods: A review of rules, physiological constraints, and evolutionary significance. Hydrobiologia 2001, 453, 91–105. [Google Scholar] [CrossRef]
- Mittmann, B.; Ungerer, P.; Klann, M.; Stollewerk, A.; Wolff, C. Development and staging of the water flea Daphnia magna (Straus, 1820; Cladocera, Daphniidae) based on morphological landmarks. EvoDevo. 2014, 5, 12. [Google Scholar] [CrossRef]
- Udit, U.; Biswal, A.; Mane, A.; Sinha, V.; Hussan, A.; Munilkumar, S. Culture of Brachionus calyciflorus as fish food organisms: An approach to improve larval survival of freshwater fish. J. Exp. Zool. India 2020, 23, 313–321. [Google Scholar]
- Kodama, Y.; Fujishima, M. Endosymbiotic Chlorella variabilis reduces mitochondrial number in the ciliate Paramecium bursaria. Scient. Rep. 2022, 12, 8216. [Google Scholar] [CrossRef]
- Müller, T.; Pál, L.; Bódis, M.; Kucska, B.; Wágner, L.; Bercsényi, M.; Husvéth, F.; Szabó, G.; Molná, T. Effect of dietary fat replacement on body composition of intensively reared hybrids of pikeperch and Volga pikeperch. Isr. J. Aquacult. Bam. 2012, 64, 1–7. [Google Scholar] [CrossRef]
- Paffenhöfer, G.A.; Strickler, J.R.; Lewis, K.D.; Richman, S. Motion behavior of nauplii and early copepodid stages of marine planktonic copepods. J. Plankt. Res. 1996, 18, 1699–1715. [Google Scholar] [CrossRef]
- Titelman, J.; Kiørboe, T. Motility of copepod nauplii and implications for food encounter. Mar. Ecol. Prog. Ser. 2003, 247, 123–135. [Google Scholar] [CrossRef]
- Zingel, P.; Paaver, T.; Karus, K.; Agasild, H.; Nõges, T. Ciliates as the crucial food source of larval fish in a shallow eutrophic lake. Limn. Oceano. 2012, 57, 1049–1056. [Google Scholar] [CrossRef]
- Zingel, P.; Agasild, H.; Karus, K.; Buholce, L.; Nõges, T. Importance of ciliates as food for fish larvae in a shallow sea bay and a large shallow lake. Europ. J. Prot. 2019, 67, 59–70. [Google Scholar] [CrossRef]
- Bruno, E.; Mahjoub, M.; Hansen, B.; Munk, P.; Støttrup, J.G. Feeding behavior and capture success of turbot Psetta maxima larvae during the transition from upright to tilted swimming position. Aquat. Liv. Res. 2017, 30, 35. [Google Scholar] [CrossRef]
- Kestemont, P.; Xueliang, X.; Hamza, N.; Maboudou, J.; Toko, I. Effect of weaning age and diet on pikeperch larviculture. Aquaculture 2007, 264, 197–204. [Google Scholar] [CrossRef]
- El Kertaoui, N.; Lund, I.; Assogba, H.; Domínguez, D.; Izquierdo, M.S.; Baekelandt, S.; Cornet, V.; Mandiki, S.N.M.; Montero, D.; Kestemont, P. Key nutritional factors and interactions during larval development of pikeperch (Sander lucioperca). Sci. Rep. 2019, 9, 7074. [Google Scholar] [CrossRef]
- Ajiboye, O.O.; Yakubu, A.F.; Adams, T.E.; Olaji, E.D.; Nwogu, N.A. A review of the use of copepods in marine fish larviculture. Rev. Fish Biol. Fish 2010, 21, 225–246. [Google Scholar] [CrossRef]
- Hansen, B.W. Advances using copepods in aquaculture. J. Plankt. Res. 2017, 39, 972–974. [Google Scholar] [CrossRef]
- Rasdi, N.W.; Qin, J. Copepod supplementation as a live food improved growth and survival of Asian seabass Lates calcarifer larvae. Aquacult. Res. 2018, 49, 3606–3613. [Google Scholar] [CrossRef]
- Chakrabarti, R.; Sharma, J.G. Influence of management protocols on carp growth under nursery conditions: Relative importance of food and water quality. Aquac. Int. 1998, 6, 293–301. [Google Scholar] [CrossRef]
- Piasecki, W.; Goodwin, A.E.; Eiras, J.C.; Nowak, B.F. Importance of copepoda in freshwater aquaculture. Zool. Stud. 2004, 43, 193–205. [Google Scholar]
- Ghan, D.; Sprules, W.G. Diet, prey selection, and growth of larval and juvenile burbot Lota lota (L.). J. Fish Biol. 1993, 42, 47–64. [Google Scholar] [CrossRef]
- Donner, M.T.; Eckmann, R. Diel vertical migration of larval and early-juvenile burbot optimises survival and growth in a deep, pre-alpine lake. Freshw. Biol. 2011, 56, 916–925. [Google Scholar] [CrossRef]
- Żarski, D.; Sasinowski, W.; Kucharczyk, D.; Kwiatkowski, M.; Krejszeff, S.; Targońska, K. Mass initial rearing of burbot Lota lota (L.) larvae under controlled conditions. Pol. J. Nat. Sci. 2009, 24, 76–84. [Google Scholar] [CrossRef]
- Wocher, H.; Harsányi, A.; Schwarz, J.F. Larviculture of burbot (Lota lota L.): Larval rearing using Artemia and weaning onto dry feed. Aquac. Res. 2013, 44, 106–113. [Google Scholar] [CrossRef]
- Lahnsteiner, F. Digestive enzyme system of larvae of different freshwater teleosts and its differentiation during the initial phase of exogenous feeding. Czech J. Animal Sci. 2017, 62, 403–416. [Google Scholar] [CrossRef]
- Esmaeilzadeh-Leithner, S.; Wanzenböck, J. Suitability of two agglomerated commercial microdiets for rearing larvae of different strains of Coregonus lavaretus under cold-water conditions. Aquac. Nutr. 2017, 24, 260–268. [Google Scholar] [CrossRef]
- Billard, R. The Carp: Biology and Culture; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Mitra, G.; Mukhopadhyay, P.K.M.; Ayyappan, S. Biochemical composition of zooplankton community grown in freshwater earthen ponds: Nutritional implication in nursery rearing of fish larvae and early juveniles. Aquaculture 2007, 272, 346–360. [Google Scholar] [CrossRef]
- Zaki, M.I.; Saad, H. Comparative study on growth and survival of larval and juvenile Dicentrarchus labrax rearing on rotifer and Artemia enriched with four different microalgae species. Afr. J. Biotech. 2010, 9, 3676–3688. [Google Scholar]
- Fernández-Diaz, C.; Pascual, E.; Yúfera, M. Feeding behaviour and prey size selection of gilthead seabream, Sparus aurata, larvae fed on inert and live food. Marine Biol. 1994, 118, 323–328. [Google Scholar] [CrossRef]
- Støttrup, J.G.; Attramadal, Y. The influence of different rotifer and Artemia enrichment diets on growth, survival and pigmentation in turbot (Scophthalmus maximus L.) larvae. J. World Aquacult. Soc. 2007, 23, 307–316. [Google Scholar] [CrossRef]
- Hamre, K.; Ted, A.; Mollan, T.A.; Sæle, Ø.; Erstad, B. Rotifers enriched with iodine and selenium increase survival in Atlantic cod (Gadus morhua) larvae. Aquaculture 2008, 284, 190–195. [Google Scholar] [CrossRef]
- Karlsen, Ø.; van der Meeren, T.; Rønnestad, I.; Mangor-Jensen, A.; Galloway, T.F.; Kjørsvik, E.; Hamre, K. Copepods enhance nutritional status, growth and development in Atlantic cod (Gadus morhua L.) larvae—Can we identify the underlying factors? PeerJ. 2015, 3, e902. [Google Scholar] [CrossRef]
- Mona, M.; Rizk, E.T.; El-feky, M.; Elawany, M. Effect of nutritional quality of rotifers and copepods on sea bream (Sparus aurata) fry fish productivity. Egypt. J. Exp. Biol. 2019, 15, 135–142. [Google Scholar] [CrossRef]
- Puello-Cruz, A.; Flores-Rojas, A.; Almazán-Rueda, P.; García-Ortega, A. Use of copepods during early feeding stages of spotted rose snapper larvae in a scale-up system. North Am. J. Aquacult. 2015, 77, 230–238. [Google Scholar] [CrossRef]
- Turgay, E.; Steinum, T.M.; Eryalçın, K.M.; Yardımcı, R.E.; Karataş, S. The influence of diet on the microbiota of live-feed rotifers (Brachionus plicatilis) used in commercial fish larviculture. FEMS Microbiol Lett. 2020, 367, fnaa020. [Google Scholar] [CrossRef]
- Vázquez-Salgado, L.; Marsella, A.; Biasini, L.; Buratin, A.; Pretoo, T.; Melchiotti, E.; Bandín, I.; Toffan, A. Role of rotifers in betanodavirus transmission to European sea bass larvae. Front. Vet. Sci. 2022, 9, 932327. [Google Scholar] [CrossRef]
- Johnson, S.; Treasurer, J.; Bravo, S.; Nagasawa, K.; Kabata, Z. A review of the impact of parasitic copepods on marine aquaculture. Zool. Stud. 2004, 43, 229–243. [Google Scholar]
- Lahnsteiner, F.; Kletzl, M.; Weismann, T. The risk of parasite transfer to juvenile fishes by live copepod food with the example Triaenophorus crassus and Triaenophorus nodulosus. Aquaculture 2009, 295, 120–125. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Measurements [mm] | L. lota | S. lucioperca | C. atterensis |
|---|---|---|---|
| Total length | 3.6 ± 0.1 a | 5.0 ± 0.2 b | 13.1 ± 0.7 c |
| Body width | 0.3 ± 0.0 a | 0.6 ± 0.1 b | 0.9 ± 0.1 c |
| Mouth width | 0.23 ± 0.05 a | 0.42 ± 0.03 b | 0.90 ± 0.10 b |
| Mouth length | 0.15 ± 0.04 a | 0.32 ± 0.06 b | 0.59 ± 0.16 b |
| Live Feed | Length of Organism [mm] | Width of Organism [mm] |
|---|---|---|
| Nauplii and copepodites (lake zooplankton) | 0.21 ± 0.03 a | 0.10 ± 0.01 a |
| Artemia nauplii | 0.46 ± 0.04 c | 0.18 ± 0.05 a |
| P. bursaria | 0.15 ± 0.04 d | 0.08 ± 0.02 a |
| B. plicatilis | 0.20 ± 0.03 a | 0.11 ± 0.01 a |
| B. calyciflorus | 0.18 ± 0.03 f | 0.10 ± 0.01 a |
| Feeding Regime | Survival Rate [%] | Total Length [mm] | Body Width [mm] |
|---|---|---|---|
| Larvae starting exogenous feeding | - | 5.0 ± 0.2 a | 0.6 ± 0.1 a |
| (A) B. plicatilis for 10 d | 79.0 ± 7.0 a | 7.7 ± 0.4 b | 0.7 ± 0.1 b |
| (B) B. calyciflorus for 10 d | 84.0 ± 5.0 a | 7.7 ± 0.4 b | 0.7 ± 0.1 b |
| (C) P. bursaria for 10 d | 80.0 ± 9.0 a | 7.3 ± 0.9 b | 0.8 ± 0.1 b |
| (D) Artemia nauplii for 10 d | 17.0 ± 16.0 b | 10.7 ± 2.7 c | 1.0 ± 0.3 c |
| (E) Lake zooplankton for 10 d | 33.0 ± 14.0 c | 10.0 ± 3.0 c | 1.1 ± 0.2 c |
| Feeding Regime | Survival Rate | Body Mass | Total Length | Body Width | ||
|---|---|---|---|---|---|---|
| Day 1–10 | Day 11–20 | [%] | [mg] | [mm] | [mm] | |
| Larvae starting exogenous feeding | - | 2.0 ± 0.3 a | 5.0 ± 0.2 a | 0.6 ± 0.1 a | ||
| (F) | B. plicatilis | Artemia nauplii | 44.0 ± 3.0 a | 23.1 ± 13.1 b | 12.0 ± 2.1 b | 1.6 ± 0.4 b |
| (G) | B. calyciflorus | Artemia nauplii | 38.0 ± 5.0 a | 7.6 ± 1.3 c | 9.7 ± 1.9 c | 0.9 ± 0.2 c |
| (H) | B. calyciflorus | Lake zooplankton | 67.5 ± 11.0 b | 7.4 ± 1.5 c | 8.6 ± 0.7 c | 1.1 ± 0.2 c |
| (I) | P. bursaria | Artemia nauplii | 47.0 ± 9.0 a | 9.0 ± 2.8 c | 8.5 ± 1.8 c | 1.0 ± 0.2 c |
| (J) | P. bursaria | Lake zooplankton | 70.3 ± 4.5 b | 9.0 ± 2.8 c | 7.9 ± 1.4 c | 0.9 ± 0.2 c |
| (K) | Lake zooplankton | Lake zooplankton | 36.0 ± 6.0 a | 18.0 ± 5.4 b | 11.6 ± 1.0 b | 1.5 ± 0.4 b |
| Feeding Regime | Survival Rate | Body Mass | Total Length | Body Width | ||
|---|---|---|---|---|---|---|
| Day 1–10 | Day 11–20 | [%] | [mg] | [mm] | [mm] | |
| Larvae starting exogenous feeding | - | - | 3.6 ± 0.1 a | 0.3 ± 0.0 a | ||
| (F) | B. plicatilis | Artemia nauplii | 44.0 ± 3.0 a | 3.0 ± 0.8 a | 4.9 ± 0.3 b | 0.9 ± 0.1 b |
| (G) | B. calyciflorus | Artemia nauplii | 38.0 ± 5.0 a | 2.2 ± 0.8 b | 4.8 ± 0.5 b | 0.8 ± 0.1 b |
| (J) | P. bursaria | Lake zooplankton | 74.0 ± 6.0 b | 3.5 ± 0.9 c | 5.5 ± 0.0 b | 1.3 ± 0.5 c |
| (H) | B. calyciflorus | Lake zooplankton | 77.0 ± 11.0 b | 3.7 ± 0.6 c | 5.6 ± 0.4 b | 1.6 ± 0.4 c |
| (L) | Lake zooplankton | Lake zooplankton | 83.0 ± 9.0 b | 4.2 ± 1.1 c | 5.8 ± 1.2 b | 1.7 ± 0.5 c |
| Feeding Regime | Survival Rate | Body Mass | Total Length | Body Width | ||
|---|---|---|---|---|---|---|
| Day 1–10 | Day 11–20 | [%] | [mg] | [mm] | [mm] | |
| Larvae starting exogenous feeding | - | 6.6 ± 1.2 a | 13.1 ± 0.7 a | 0.9 ± 0.1 a | ||
| (F) | B. plicatilis | Artemia nauplii | 59.2 ± 11.9 a | 12.5 ± 4.8 b | 15.1 ± 0.6 b | 2.0 ± 0.5 b |
| (H) | B. calyciflorus | Lake zooplankton | 73.4 ± 8.3 b | 12.8 ± 2.6 b | 14.9 ± 0.9 b | 2.2 ± 0.2 b |
| (L) | Lake zooplankton | Lake zooplankton | 86.4 ± 7.9 c | 24.1 ± 6.9 c | 15.2 ± 0.7 b | 2.8 ± 0.4 c |
| (M) | Artemia nauplii | Artemia nauplii | 51.0 ± 7.3 a | 27.8 ± 8.1 c | 15.8 ± 1.9 b | 2.8 ± 0.5 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lahnsteiner, F.; Lahnsteiner, E.; Duenser, A. Suitability of Different Live Feed for First Feeding of Freshwater Fish Larvae. Aquac. J. 2023, 3, 107-120. https://doi.org/10.3390/aquacj3020010
Lahnsteiner F, Lahnsteiner E, Duenser A. Suitability of Different Live Feed for First Feeding of Freshwater Fish Larvae. Aquaculture Journal. 2023; 3(2):107-120. https://doi.org/10.3390/aquacj3020010
Chicago/Turabian StyleLahnsteiner, Franz, Elias Lahnsteiner, and Anna Duenser. 2023. "Suitability of Different Live Feed for First Feeding of Freshwater Fish Larvae" Aquaculture Journal 3, no. 2: 107-120. https://doi.org/10.3390/aquacj3020010
APA StyleLahnsteiner, F., Lahnsteiner, E., & Duenser, A. (2023). Suitability of Different Live Feed for First Feeding of Freshwater Fish Larvae. Aquaculture Journal, 3(2), 107-120. https://doi.org/10.3390/aquacj3020010