
Life’s a Gas: A Thermodynamic Theory of Biological Evolution

Abstract
:
1. Introduction
1.1. Biological Evolution
- (1)
- genetic variation (a combination of mutation, segregation and independent recombination) exists within any given population for almost all traits;
- (2)
- The transfer of genes from one generation to the next occurs independently of other genes;
- (3)
- In general, more offspring are produced than survive to reproduce;
- (4)
- Individuals that do survive and procreate represent a subset of the population of that generation which are genetically best adapted to their environment.
1.2. Thermodynamics
1.2.1. Beginnings
- -
- Heat could neither be created nor destroyed and that the total heat of the universe was constant;
- -
- When a temperature gradient exists, work can be done;
- -
- There can be no such thing as perpetual motion.
1.2.2. The Maximum Entropy Production Principle (MEPP)
1.3. Thermodynamics and Evolution
2. A Thermodynamic Theory of Evolution
- (1)
- An explanation of the known facts relating to the origin, architecture, structure and function of life on earth from an abiotic beginning (be that on this planet or on another astronomical body (e.g., Panspermia));
- (2)
- An explanation for the tempo of biological evolution as observed in the fossil record;
- (3)
- An explanation of the diversity and ecology of life on earth;
- (4)
- Such an explanation should not be contrary to observations made at the molecular, organismal, population nor ecosystem levels of organization.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level of organization | Thermodynamic control |
|---|---|
| DNA | Random mutations increase information entropy within genetic material while correction mechanisms increase entropy out with genetic material (i.e. correction consumes free energy) [39,69]. |
| Amino acids | Early abiogenic amino acids shown to form along a thermodynamic gradient [70]. Later biogenic amino acids produce increased entropy of formation. Early coding constrained thermodynamically in terms of concentration and availability of amino acids [71,72,73]. |
| Proteins | Amino acid content often constrained by energetic limitations [72]. Folding and function of proteins thermodynamically determined [74,75]. |
| Cells | Cellularity allows reduction of entropy within cells, providing the stability needed for cellular physiology [76]. Cells also export high levels of entropy [77]. Metabolic networks evolve towards maximum entropy production [78]. |
| Organisms | Increasing complexity brings increasing entropic dissipation. Advent of multicellularity, cellular specialization, increasing size and homeothermy all increased entropy production [76]. Form represents diffusion into ecospace, while function conforms to laws of thermodynamics. |
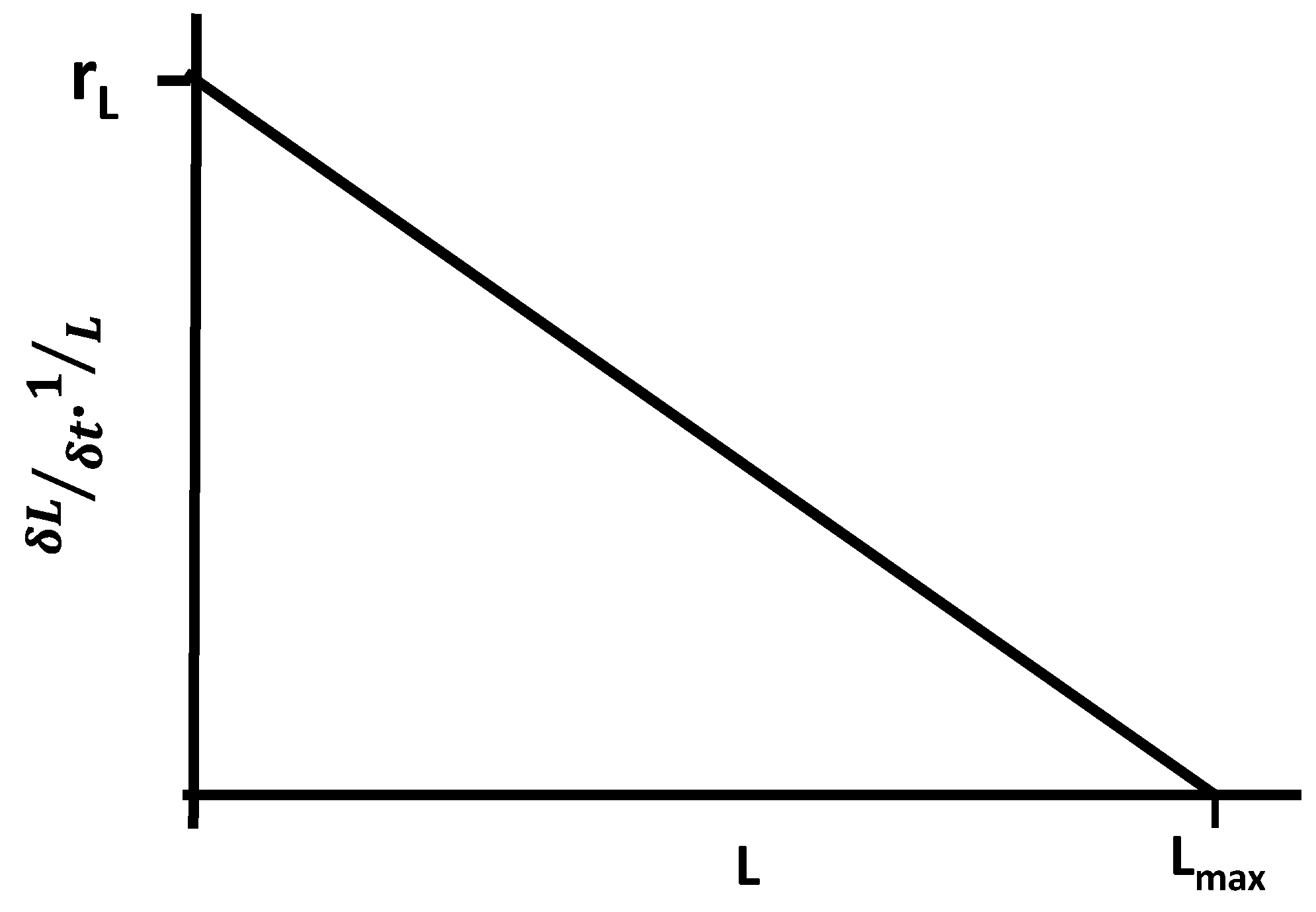
| Populations | Logistic growth of populations follows MEPP, wherein Krepresents Smax. Increased free energy availability allows population increase within ecosystem constraints [79]. |
| Speciation | Exploration of ecospace through random mutations delivers increased diversity, expressed as speciation if reproductive barriers are in place. Speciation is a eukaryotic trait, since bacteria share DNA plasmids between taxa making the species concept redundant for prokaryotes [80]. |
| Ecosystems | Ecological succession follows the MEPP [81,82,83] with ecosystems moving towards Smax [84]. |
| Biomes | Biome conditions determined by solar radiation density, interacting with the hydrological cycle, and forming the backdrop within which ecosystems develop [85,86]. |
| Earth system | Climate, global circulation patterns and tectonic activity all shown to follow MEPP [36,40,87]. |
3. A Thermodynamic Model of Biological Diversity through Time

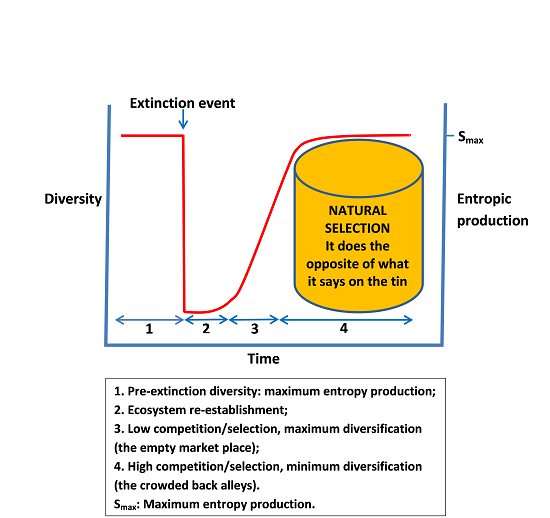
3.1. Increasing Diversity Towards a Dynamic Equilibrium of Maximum Entropy Production

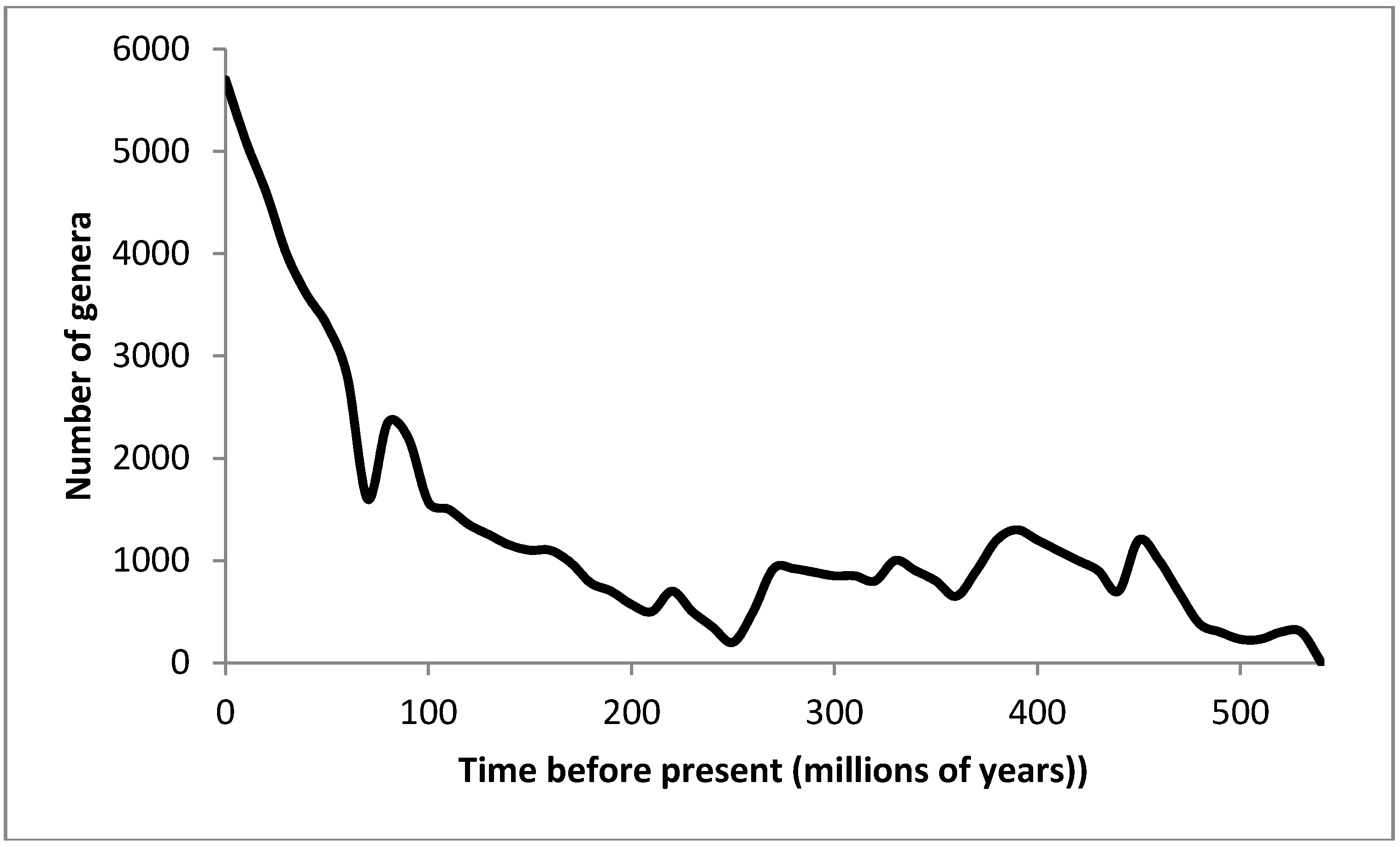
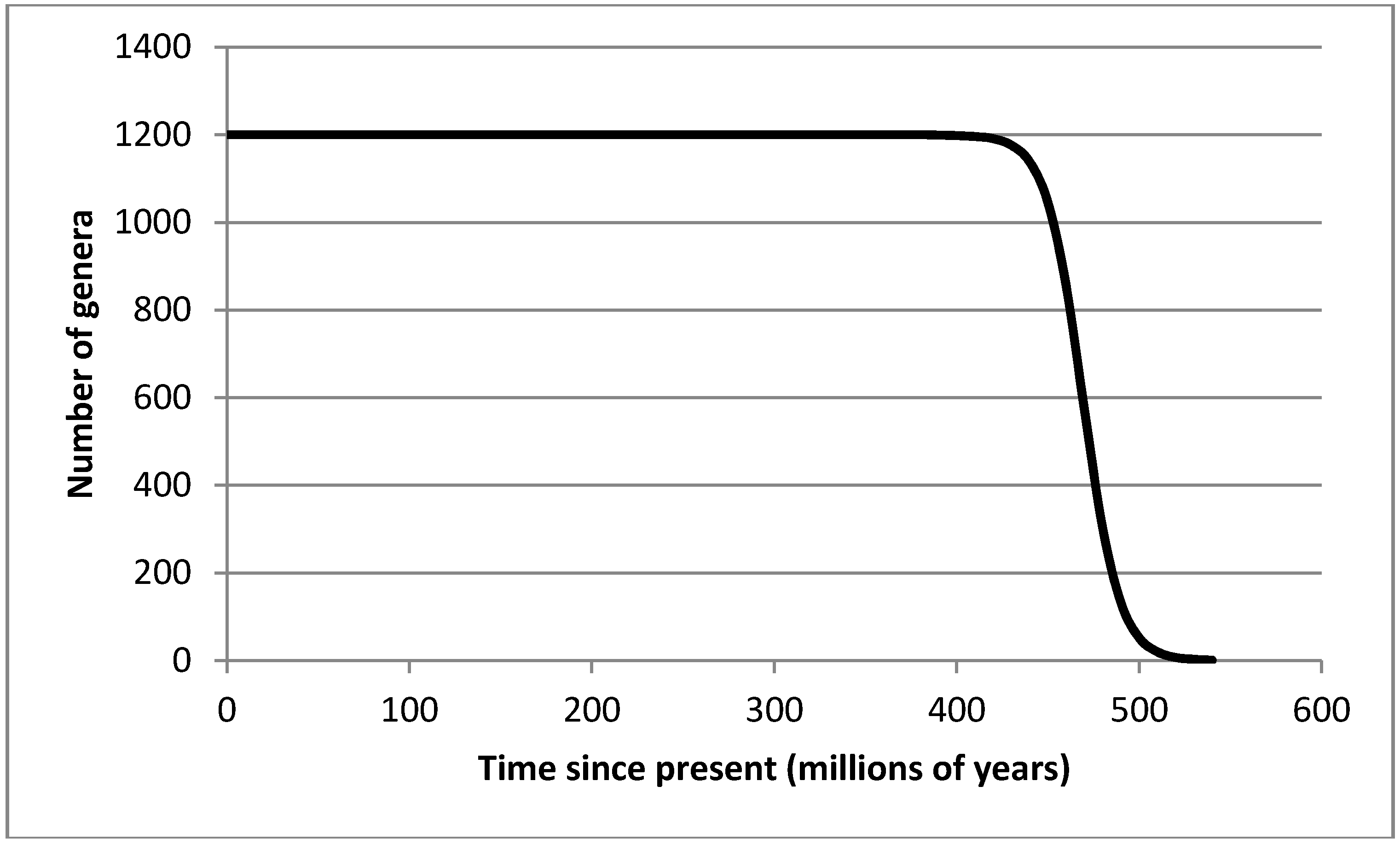
3.2. Recovery from Significant Extinction Events
| Years before present (×106) | Event | Proportion of pre-extinction diversity remaining (De) |
|---|---|---|
| 440 | Late Ordovician | 0.6 |
| 392 | Devonian 1 | 0.72 |
| 370 | Devonian 2 | 0.65 |
| 360 | Devonian 3 | 0.69 |
| 340 | Devonian 4 | 0.69 |
| 257 | End Permian 1 | 0.525 |
| 250 | End Permian 2 | 0.44 |
| 205 | Late Triassic | 0.57 |
| 183 | Torcian turnover | 0.8 |
| 154 | End Jurassic | 0.8 |
| 90 | Cenomanian | 0.86 |
| 65 | End Cretaceous | 0.6 |
| 34 | End Eocene | 0.84 |


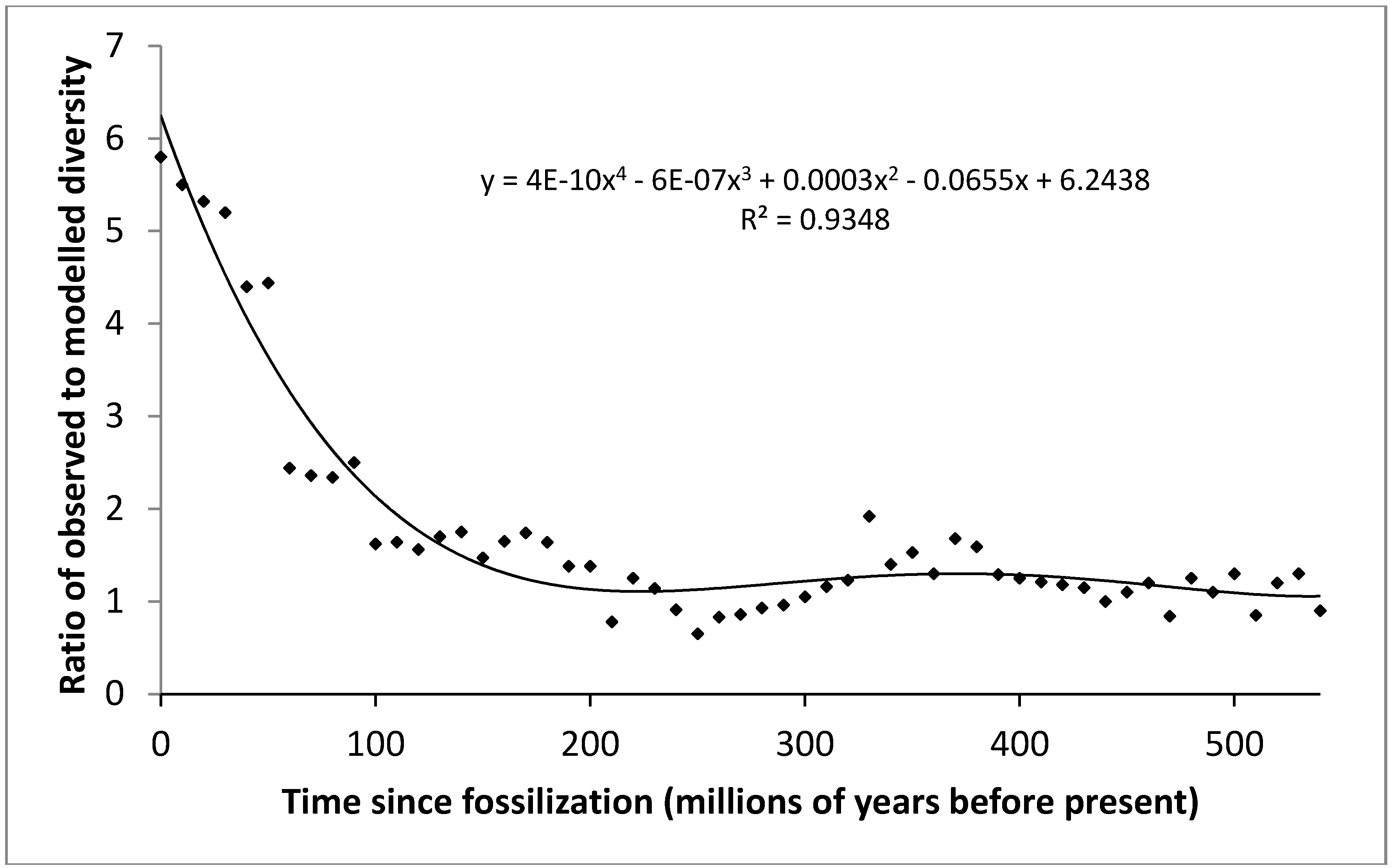
3.2. Likelihood of Discovery of Fossils Decreases with Age


4. Discussion
4.1. A Comparison of Natural Selection and Thermodynamics as Theories of Biological Evolution.
4.1.1. Progress
4.1.2. Tempo
4.1.3. Form vs. Function
4.1.4. Biosphere Architecture
4.1.5. Competition
4.1.6. Fitness
5. Conclusions
Acknowledgements
Conflicts of Interest
References
- Teilhard de Chardin, P. The Phenomenon of Man; Harper: New York, NY, USA, 1973. [Google Scholar]
- Baverstock, K.; Rönkkö, M. The evolutionary origin of form and function. J. Physiol. Lond. 2014, 592, 2261–2265. [Google Scholar] [CrossRef] [PubMed]
- Lamarck, J.-B. Philosophie Zoologique, ou Exposition des Considérations Relative à l'Histoire Naturelle des Animaux, Volume 1; Librairie P. Savy: Paris, France, 1809. [Google Scholar]
- Wells, W.C. Two Essays: Upon a Single Vision with Two Eyes, the Other on Dew; Constable: London, UK, 1818. [Google Scholar]
- Matthew, P. Naval Timbers and Arboriculture: with Critical Notes on Authors who have Recently Treated the Subject of Planting; Adam Black: Edinburgh, UK, 1931. [Google Scholar]
- Blyth, E. An attempt to classify the “varieties” of animals, with observations on the marked seasonal and other changes which naturally take place in various British species, and which do not constitute varieties. Mag. Nat. Hist. 1835, 8, 40–53. [Google Scholar]
- Wallace, A.R. On the tendency of varieties to indefinitely from the original type. Proc. Linnean Soc. 1858, 3, 53–62. [Google Scholar]
- Gould, S.J.; Lewontin, R.C. The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. Lond. B 1979, 205, 581–598. [Google Scholar] [CrossRef] [PubMed]
- Voltaire. Candide, ou l’Optimisme; Cramer, Marc-Michel Rey, Jean Nourse, Lambert and others: Paris, France, 1759. [Google Scholar]
- Mendel, J.G. Versuche über Pflanzenhybriden Verhandlungen des naturforschenden Vereines in Brünn. Bd. IV für das Jahr, Abhandlungen 1866, 3–47. [Google Scholar]
- Fisher, R.A. The Genetical Theory of Natural Selection; Dover: New York, USA, 1930. [Google Scholar]
- Huxley, J. Evolution: the Modern Synthesis; Allen and Unwin: London, UK, 1942. [Google Scholar]
- Pittendrigh, C. Adaptation, natural selection and behaviour. In Behaviour and Evolution; Roe, A., Simpson, G.G., Eds.; Yale University Press: New Haven, CT, USA, 1958; pp. 390–416. [Google Scholar]
- Jablonka, E.; Lamb, M.J. Soft inheritance: challenging the modern synthesis. Genet. Mol. Biol. 2008, 31, 389–395. [Google Scholar] [CrossRef]
- Pigliucci, M.; Müller, G.B. Evolution: the Extended Synthesis; MIT Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Danchin, E.; Charmantier, A.; Champagne, F.A.; Mesoudi, A.; Pujol, B.; Blanchet, S. Beyond DNA: integrating inclusive inheritance into an extended theory of evolution. Nat. Genet. 2011, 12, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Carnot, S. Réflexions sur la Puissance Motrice du Feu et sur les Machines Propres a Développer Cette Puissance; Chez Bachelier: Paris, France, 1824. [Google Scholar]
- Clausius, R. Über die bewegende Kraft der Wärme, Part I, Part II. Ann. Phys. Berl. 1850, 79, 368–397, 500–524. [Google Scholar] [CrossRef]
- Thomson, W. On the dynamical theory of heat, with numerical results deduced from Mr Joule’s equivalent of a thermal unit, and M. Regnault’s observations on steam. Trans. R. Soc. Edinb. 1851, XX, 261–268. [Google Scholar] [CrossRef]
- Clausius, R. The Mechanical Theory of Heat: With its Applications to the Steam-Engine and to the Physical Properties of Bodies; John van Voorst: London, UK, 1867. [Google Scholar]
- Nernst, W. The New Heat Theorem; Methuen and Co.: London, UK, 1917. [Google Scholar]
- Saha, M.N.; Srivastava, B.N. A Treatise on Heat; Including Kinetic Theory of Gases, Thermodynamics and Recent Advances in Statistical Thermodynamics, the second and revised edition of A Text Book of Heat; The Indian Press: Allahabad and Calcutta, India, 1935. [Google Scholar]
- Gibbs, J.W. Elementary Principles in Statistical Mechanics; Charles Scribner’s Sons: New York, NY, USA, 1902. [Google Scholar]
- Jaynes, E.T. Information theory and statistical mechanics. Phys. Rev. 1957, 106, 620–630. [Google Scholar] [CrossRef]
- Jaynes, E.T. Information theory and statistical mechanics. II. Phys. Rev. 1957, 108, 171–190. [Google Scholar] [CrossRef]
- Hatsopoulos, G.N.; Keenan, J.H. Principles of General Thermodynamics (Vol. 398); Wiley: New York, NY, USA, 1965. [Google Scholar]
- Berthelot, M. Essai de Mecanique Chimique Fondee sur la Thermochimie; Dunod: Paris, France, 1879. [Google Scholar]
- Rayleigh, L. On our perception of the direction of a source of sound. In Proceedings of the Musical Association, 2nd Session; Royal Musical Association: London, UK, 1875; pp. 75–84. [Google Scholar]
- Onsager, L. Reciprocal relations in irreversible processes. II. Phys. Rev. 1931, 38, 2265–2279. [Google Scholar] [CrossRef]
- Ziegler, H. Some extremum principles in irreversible thermodynamics with applications to continuum mechanics. In Progress in Solid Mechanics; Sneddon, I.N., Hill, R., Eds.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1963; pp. 92–193. [Google Scholar]
- Jaynes, E.T. Concentration of distributions at entropy maxima. In E. T. Jaynes: Papers on Probability, Statistics, and Statistical Physics; Rosenkrantz, R.D., Ed.; D. Reidel Publishing Company: Dordrecht, The Netherlands, 1983; pp. 315–336. [Google Scholar]
- Kleidon, A.; Malhi, Y.; Cox, P.M. Maximum entropy production in environmental and ecological systems. Proc. R. Soc. Lond. B 2010, 365, 1297–1302. [Google Scholar] [CrossRef] [PubMed]
- Kleidon, A.; Lorenz, R.D. Entropy production by earth system processes. In Non-Equilibrium Thermodynamics and the Production of Entropy: Life, Earth, and Beyond; Kleidon, A., Lorenz, R.D., Eds.; Springer: Berlin, Germany, 2004; pp. 1–20. [Google Scholar]
- Martyushev, L.M.; Seleznev, V.D. Maximum entropy production principle in physics, chemistry and biology. Phys. Rep. 2006, 426, 1–45. [Google Scholar] [CrossRef]
- Harte, J. Maximum Entropy and Ecology: On the Inference of Patterns in Nature; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Paltridge, G.W. Global dynamics and climate change: a system of minimum entropy exchange. Q. J. R. Meteorol. Soc. 1975, 101, 475–484. [Google Scholar] [CrossRef]
- Stoy, P.C.; Lin, H.; Novick, K.A.; Siqueira, M.; Juang, J.Y. The role of vegetation on the ecosystem radiative entropy budget and trends along ecological succession. Entropy 2014, 16, 3710–3731. [Google Scholar] [CrossRef]
- Berger, A.L.; Pietra, V.J.D.; Pietra, S.A.D. A maximum entropy approach to natural language processing. Comput. Linguist. 1996, 22, 39–71. [Google Scholar]
- Salamon, P.; Konopka, A.K. A maximum entropy principle for the distribution of local complexity in naturally occurring nucleotide sequences. Comput. Chem. 1992, 16, 117–124. [Google Scholar]
- Dong, W.M.; Bao, A.B.; Shah, H.C. Use of maximum entropy principle in earthquake recurrence relationships. Bull. Seismol. Soc. Am. 1984, 74, 725–737. [Google Scholar]
- Morel, R.E.; Fleck, G. A fourth law of thermodynamics. Chemistry 2006, 15, 305–310. [Google Scholar]
- Serizawa, H.; Amemiya, T.; Itoh, K. Tree network formation in Poisson equation models and the implications for the maximum entropy production principle. Nat. Sci. 2014, 6, 514. [Google Scholar] [CrossRef]
- De Vladar, H.P.; Barton, N.H. The contribution of statistical physics to evolutionary biology. Trends Ecol. Evol. 2011, 26, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Depew, D.J.; Weber, B.H. Darwinism Evolving: Systems Dynamics and the Genealogy of Natural Selection; Bradford Books/MIT Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Demetrius, L. Thermodynamics and evolution. J. Theor. Biol. 2000, 206, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Collier, J. The dynamics of biological order. In Entropy, information and Evolution; Weber, B.H., Depew, D.J., Smith, J.D., Eds.; MIT Press: Cambridge, MA, USA, 1988; pp. 227–242. [Google Scholar]
- Pross, A. Toward a general theory of evolution: Extending Darwinian theory to inanimate matter. J. Syst. Chem. 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Molchanov, A.M. Thermodynamics and evolution. In Oscillatory Processes in Biological and Chemical Systems; Frank, G.M., Ed.; Nauka: Moscow, USSR, 1967; pp. 292–305. [Google Scholar]
- Prigogine, I.; Nicolis, G.; Babloyantz, A. Thermodynamics of evolution. Phys. Today 1972, 25, 23–28. [Google Scholar] [CrossRef]
- De Beer, G. Some unpublished letters of Charles Darwin. Notes Rec. R. Soc. 1959, 14, 12–66. [Google Scholar] [CrossRef]
- Thompson, D.W. On Growth and Form; Cambridge University Press: Cambridge, UK, 1916. [Google Scholar]
- Boltzmann, L. The second law of thermodynamics. In Theoretical Physics and Philosophical Problems: Selected Writings; McGinness, B., Ed.; D. Reidel: New York, NY, USA, 1974; pp. 13–32. [Google Scholar]
- Lotka, A.J. Contribution to the energetics of evolution. Proc. Natl. Acad. Sci. USA 1922, 8, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Lotka, A.J. Natural selection as a physical principle. Proc. Natl. Acad. Sci. USA 1922, 8, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Odum, H.T.; Pinkerton, R.C. Time’s speed regulator: The optimum efficiency for maximum output in physical and biological systems. Am. Sci. 1955, 43, 331–343. [Google Scholar]
- Odum, H.T. Self-organization and maximum empower. In Maximum Power: The Ideas and Applications of H.T. Odum; Hall, C.A.S., Ed.; Colorado University Press: Colorado, CO, USA, 1995; pp. 311–330. [Google Scholar]
- Schrödinger, E. What is Life? The Physical Aspect of the Living Cell; Cambridge University Press: Cambridge, UK, 1944. [Google Scholar]
- Prigogine, I. Order through fluctuation: self-organization and social system. In Evolution and Consciousness; Jantsch, E., Waddington, C.H., Eds.; Addison-Wesley: London, UK, 1976; pp. 93–133. [Google Scholar]
- Fenchel, T. Intrinsic rate of natural increase: the relationship with body size. Oecologia 1974, 14, 317–326. [Google Scholar] [CrossRef]
- Ulanowicz, R.E. Ecology, the Ascendent Perspective: Robert E. Ulanowicz; Columbia University Press: New York, NY, USA, 1997. [Google Scholar]
- Wiley, E.O.; Brooks, D.R. Victims of history—a nonequilibrium approach to evolution. Syst. Biol. 1982, 31, 1–24. [Google Scholar] [CrossRef]
- Annila, A.; Salthe, S. 2010. Physical foundations of evolutionary theory. J. Non-Equilib. Thermodyn. 2010, 35, 301–321. [Google Scholar] [CrossRef]
- Swenson, R. Emergent attractors and the law of maximum entropy production: foundations to a theory of general evolution. Syst. Res. 1989, 6, 187–197. [Google Scholar] [CrossRef]
- Moore, E.H. On a form of general analysis with application to linear differential and integral equations. In Proceedings of Atti del IV Congresso Internazionale dei Matematici, Rome, Italy, 6–11 April 1908; Volume 2, pp. 98–114.
- Wilson, E.O. Consilience: The Unity of Knowledge; Knopf: New York, NY, USA, 1998. [Google Scholar]
- McKay, C.P. What is life—and how do we search for it in other worlds? PloS Biol. 2004, 2, 1260–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovelock, J.E. A physical basis for life detection experiments. Nature 1965, 207, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Pascal, R.; Pross, A.; Sutherland, J.D. Towards an evolutionary theory of the origin of life based on kinetics and thermodynamics. Open Biol. 2013, 3, 130156. [Google Scholar] [CrossRef] [PubMed]
- Tessera, M.; Hoelzer, G.A. On the thermodynamics of multilevel evolution. BioSystems 2013, 113, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Higgs, P.G.; Pudritz, R.E. A thermodynamic basis for prebiotic amino acid synthesis and the nature of the first genetic code. Astrobiology 2009, 9, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Akashi, H.; Gojobori, T. Metabolic efficiency and amino acid composition in the proteomes of Escherichia coli and Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2002, 99, 3695–3700. [Google Scholar] [CrossRef] [PubMed]
- Seligmann, H. Cost-minimization of amino acid usage. J. Mol. Evol. 2003, 56, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, E.N. The triplet code from first principles. J. Biomol. Struct. Dyn. 2004, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Anfinsen, C.B. Principles that govern the folding of protein chains. Science 1973, 181, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Lazaridis, T.; Karplus, M. Thermodynamics of protein folding: a microscopic view. Biophys. Chem. 2002, 100, 367–395. [Google Scholar] [CrossRef]
- Davies, P.C.; Rieper, E.; Tuszynski, J.A. Self-organization and entropy reduction in a living cell. Biosystems 2013, 111, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marína, D.; Martín, M.; Sabater, B. Entropy decrease associated to solute compartmentalization in the cell. Biosystems 2009, 98, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Unrean, P.; Srienc, F. Metabolic networks evolve towards states of maximum entropy production. Metab. Eng. 2011, 13, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Aoki, I. Entropy production in living systems: from organisms to ecosystems. Thermochim. Acta 1995, 25, 359–370. [Google Scholar] [CrossRef]
- Skene, K.R. Shadows on a Cave Wall: A New Theory of Evolution; Ard Macha Press: Angus, UK, 2009. [Google Scholar]
- Aoki, I. Entropy balance in Lake Biwa. Ecol. Model. 1987, 37, 235–248. [Google Scholar] [CrossRef]
- Ludovisi, A. Biotic and abiotic entropy production in lake ecosystems. Ecol. Model. 2004, 179, 145–147. [Google Scholar] [CrossRef]
- Meysman, F.J.R.; Bruers, S. A thermodynamic perspective on food webs: Quantifying entropic production within detrital-based ecosystems. J. Theor. Biol. 2007, 249, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Skene, K.R. The energetics of ecological succession: A logistic model of entropic output. Ecol. Model. 2013, 250, 287–293. [Google Scholar] [CrossRef]
- Kleidon, A. Nonequilibrium thermodynamics and maximum entropy production in the Earth system. Naturwissenschaften 2009, 96, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, M.C.; Zehe, E.; Schymanski, S.J. Importance of temporal variability for hydrological predictions based on the maximum entropy production principle. Geophys. Res. Lett. 2014, 41, 67–73. [Google Scholar] [CrossRef]
- Lucarini, V.; Pascale, S. Entropy production and course graining of the climate fields in a general circulation model. Clim. Dyn. 2014, 43, 981–1000. [Google Scholar] [CrossRef]
- Wynne-Edwards, V.C. Animal Dispersion in Relation to Social Behavior; Oliver & Boyd: London, UK, 1962. [Google Scholar]
- Toussaint, O.; Schneider, E.D. The thermodynamics and evolution of complexity in biological systems. Comp. Biochem. Physiol. 1998, 120, 3–9. [Google Scholar] [CrossRef]
- Johnson, L. The thermodynamic origin of ecosystems. Can. J. Fish. Aquat. Sci. 1981, 38, 571–590. [Google Scholar] [CrossRef]
- Bambach, R.K.; Knoll, A.H.; Wang, S.C. Origination, extinction, and mass depletions of marine diversity. Paleobiology 2004, 30, 522–542. [Google Scholar] [CrossRef]
- Alroy, J. Dynamics of origination and extinction in the marine fossil record. Proc. Natl. Acad. Sci. USA 2008, 105, 11536–11542. [Google Scholar] [CrossRef] [PubMed]
- Sepkoski, J.J., Jr. A compendium of fossil marine families. Contrib. Biol. Geol. 1982, 51, 1–125. [Google Scholar]
- Smith, A.B.; McGowan, A.J. The ties linking rock and fossil records and why they are important for palaeobiodiversity studies. Geol. Soc. Lond. Spec. Publ. 2011, 358, 1–7. [Google Scholar] [CrossRef]
- Allison, P.A.; Bottjer, D.J. Taphonomy: bias and process through time. In Taphonomy: Bias and Process through Time; Allison, P.A., Bottjer, D.J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 1–17. [Google Scholar]
- Thomas, R.D.K. Taphonomy: Ecology’s loss is sedimentology’s gain. Palaios 1986, 1, 206. [Google Scholar]
- Bryan, G.H. Thermodynamics of diffusion. Nature 1907, 76, 637–638. [Google Scholar] [CrossRef]
- Truesdell, C. Thermodynamics of diffusion. In Rational Thermodynamics; Truesdell, C., Ed.; Springer: New York, NY, USA, 1984; pp. 219–236. [Google Scholar]
- Kimura, M. Diffusion models in population genetics. J. Appl. Probab. 1964, 1, 177–232. [Google Scholar] [CrossRef]
- Martyushev, L.M.; Seleznev, V.D.; Maximum entropy production principle (MEPP). Comment about restrictions and typical misconceptions of critics of MEPP. 2013; arXiv:1311.2068. [Google Scholar]
- Aoki, I. Entropy and exergy in the development of living systems: A case study of lake-ecosystems. J. Phys. Soc. Jpn. 1998, 67, 2132–2139. [Google Scholar] [CrossRef]
- Farnsworth, K.D.; Niklas, K.J. Theories of optimization, form and function in branching architecture in plants. Funct. Ecol. 1995, 9, 355–363. [Google Scholar] [CrossRef]
- Darwin, C.R. The Origin of Species by Means of Natural Selection or the Preservation of Favoured Races in the Struggle for Life; Senate: London, UK, 1994. [Google Scholar]
- Averiss, R.J.; Skene, K.R. Changes in nutrient heterogeneity in a sand dune succession at Tentsmuir Point, Eastern Scotland. Bot. J. Scotl. 2001, 53, 45–56. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Neutel, A.M.; Heesterbeek, J.A.P.; van de Koppel, J.; Hoenderboom, G.; Vos, A.; Kaldeway, C.; Berendse, F.; Ruiter, P.C. Reconciling complexity with stability in naturally assembling food webs. Nature 2007, 449, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Van de Koppel, J.; Rietkerk, M.; Weissing, F.J. Catastrophic vegetation shifts and soil degradation in terrestrial grazing systems. Trends Ecol. Evol. 1997, 12, 352–356. [Google Scholar] [CrossRef]
- Lindeman, R.L. The trophic-dynamic aspect of ecology. Ecology 1942, 23, 399–418. [Google Scholar] [CrossRef]
- Eldredge, N.; Gould, S.J. Punctuated equilibria: An alternative to phyletic gradualism. In Models in Paleobiology; Schopf, T.J.M., Ed.; Freeman Cooper: San Francisco, CA, USA, 1972; pp. 82–115. [Google Scholar]
- Gould, S.J. Wonderful Life: The Burgess Shale and the Nature of History; Norton: New York, NY, USA, 1989. [Google Scholar]
- Dawkins, C.R. The Extended Phenotype: The Long Reach of the Gene; Oxford University Press: Oxford, UK, 1982. [Google Scholar]
- Higgins, P.A.; Mastrandrea, M.D.; Schneider, S.H. Dynamics of climate and ecosystem coupling: Abrupt changes and multiple equilibria. Philos. Trans. R. Soc. Lond. B 2002, 357, 647–656. [Google Scholar]
- Von Bertalanffy, L. General system theory. Gen. Syst. 1956, 1, 11–17. [Google Scholar]
- Wolkenhauer, O. Systems biology: the reincarnation of systems theory applied in biology? Brief. Bioinform. 2001, 2, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K. Life: an Introduction to Complex Systems Biology; Springer: London, UK, 2006. [Google Scholar]
- Venditti, C.; Meade, A.; Pagel, M. Phylogenies reveal new interpretation of speciation and the Red Queen. Nature 2010, 463, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J. On the non-prevalence of competitive replacement in the evolution of tetrapods. In Evolutionary Palaeobiology; Jablonski, D., Erwin, D.H., Lipps, J., Eds.; Chicago University Press: Chicago, IL, USA, 1996; pp. 185–210. [Google Scholar]
- Brusatte, S.L.; Benton, M.J.; Ruta, M.; Lloyd, G.T. Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs. Science 2008, 321, 1485–1488. [Google Scholar] [CrossRef] [PubMed]
- Alizon, S.; Kucera, M.; Jansen, V.A.A. Competition between cryptic species explains variations in rates of lineage evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 12382–12386. [Google Scholar] [CrossRef] [PubMed]
- Mahler, D.L.; Revell, L.J.; Glor, R.E.; Losos, J.B. Ecological opportunity and the rate of morphological evolution in the diversification of Greater Antillean anoles. Evolution 2010, 64, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Phillimore, A.B.; Price, T.D. Density-dependent cladogenesis in birds. PLoS Biol. 2008, 6, e71. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef] [PubMed]
- Archibald, S.B.; Johnson, K.R.; Mathewes, R.W.; Greenwood, D.R. Intercontinental dispersal of giant thermophilic ants across the Arctic during early Eocene hyperthermals. Proc. R. Soc. Lond. B 2011, 278, 3679–3686. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Muñoz, R.; Bretman, A.; Slate, J.; Walling, C.A.; Tregenza, T. Natural and sexual selection in a wild insect population. Science 2010, 328, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- Evans, J. Quantitative genetic evidence that males trade attractiveness for ejaculate quality in guppies. Proc. R. Soc. Lond. B 2010, 277, 3195–3201. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skene, K.R. Life’s a Gas: A Thermodynamic Theory of Biological Evolution. Entropy 2015, 17, 5522-5548. https://doi.org/10.3390/e17085522
Skene KR. Life’s a Gas: A Thermodynamic Theory of Biological Evolution. Entropy. 2015; 17(8):5522-5548. https://doi.org/10.3390/e17085522
Chicago/Turabian StyleSkene, Keith R. 2015. "Life’s a Gas: A Thermodynamic Theory of Biological Evolution" Entropy 17, no. 8: 5522-5548. https://doi.org/10.3390/e17085522
APA StyleSkene, K. R. (2015). Life’s a Gas: A Thermodynamic Theory of Biological Evolution. Entropy, 17(8), 5522-5548. https://doi.org/10.3390/e17085522