
A 4-Phenoxyphenol Derivative Exerts Inhibitory Effects on Human Hepatocellular Carcinoma Cells through Regulating Autophagy and Apoptosis Accompanied by Downregulating α-Tubulin Expression


 ,
, 
Abstract
:1. Introduction
2. Results
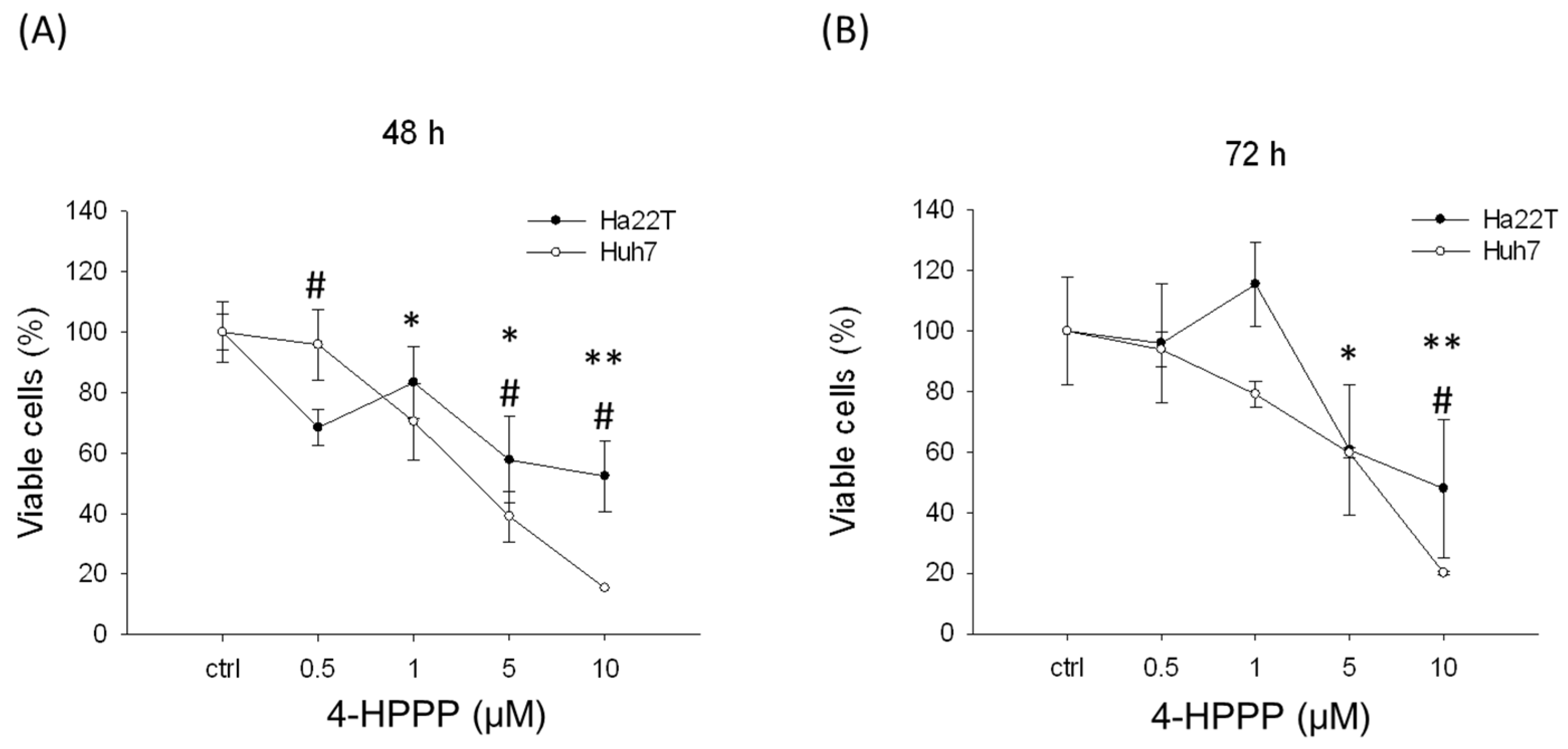
2.1. The Anti-Proliferative Effects of 4-HPPP on HCC Cells
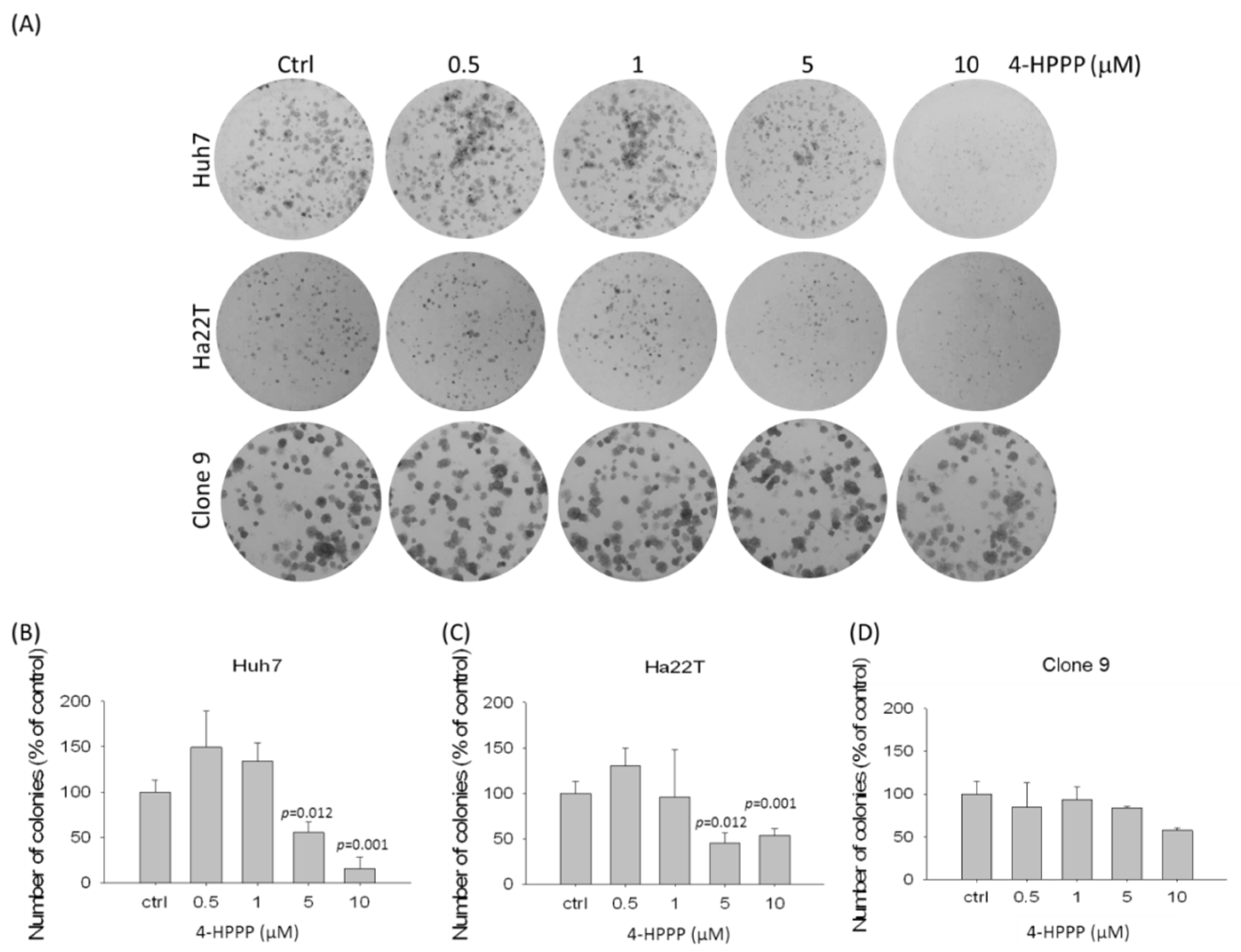
2.2. The Assessment of 4-HPPP-Induced Long-Term Anti-Proliferation of HCC
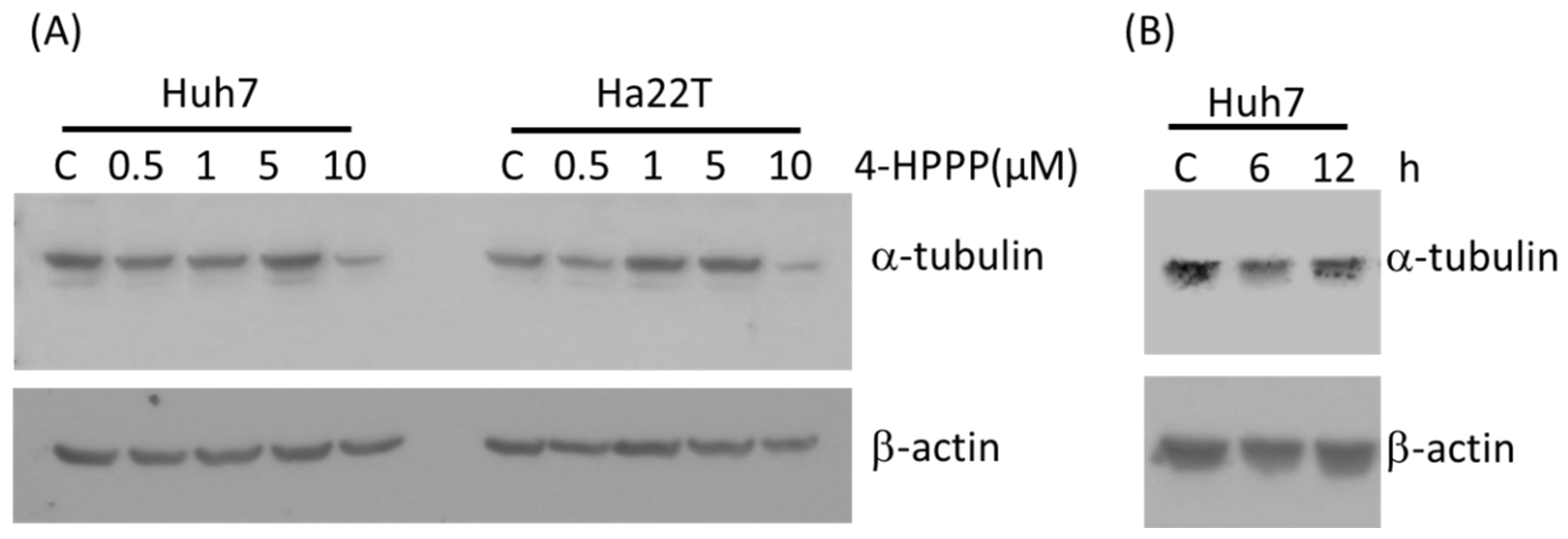
2.3. 4-HPPP Inhibits α-Tubulin Expression
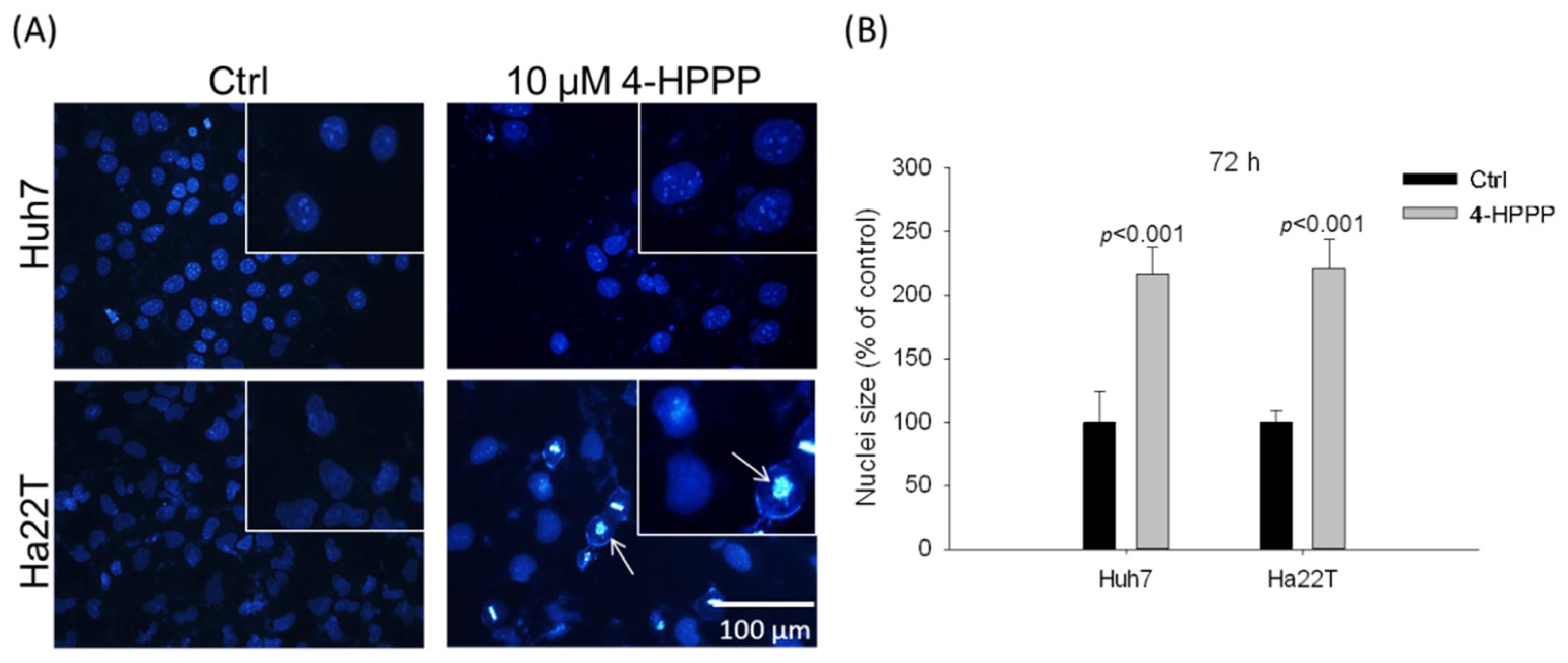
2.4. 4-HPPP Induces Nuclei Enlargement of HCC Cells
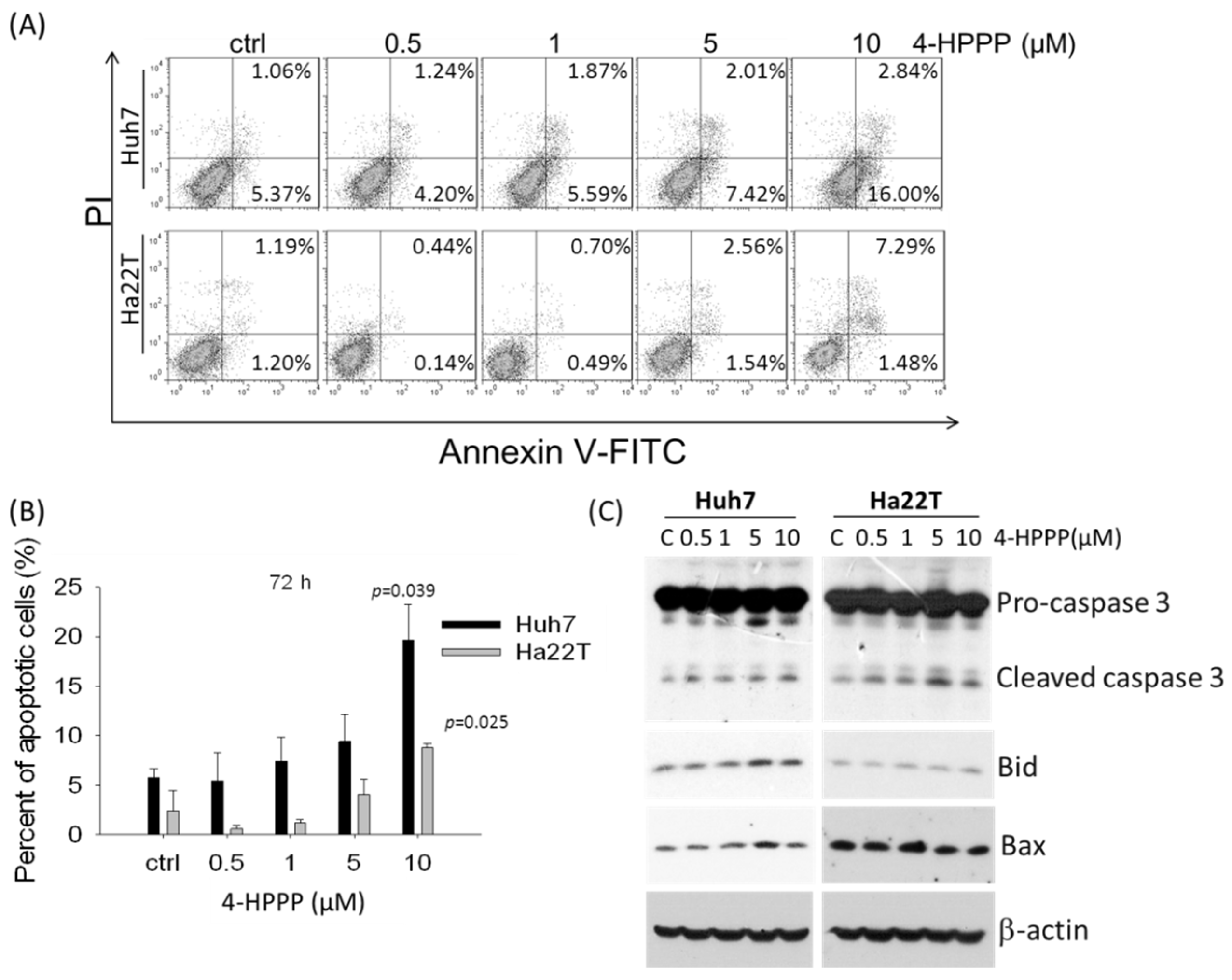
2.5. 4-HPPP Induces Apoptosis of HCC Cells
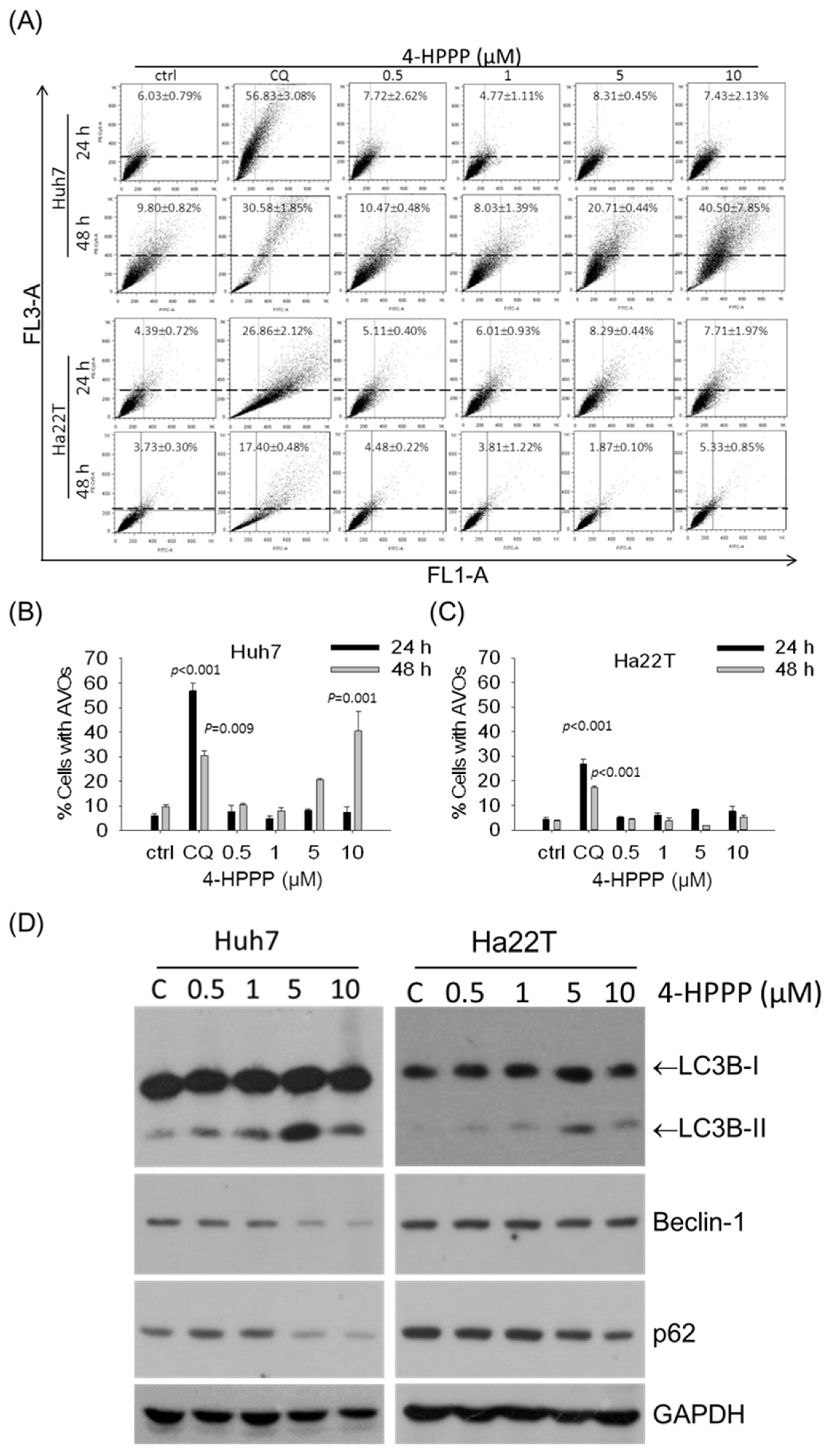
2.6. 4-HPPP-Induced Formation of Autolysosomes and the Expression of Autophagy Marker Protein in HCC Cells
2.7. 4-HPPP Induces the Activation of γH2AX and Down-Regulates the Pro-Survival Pathways
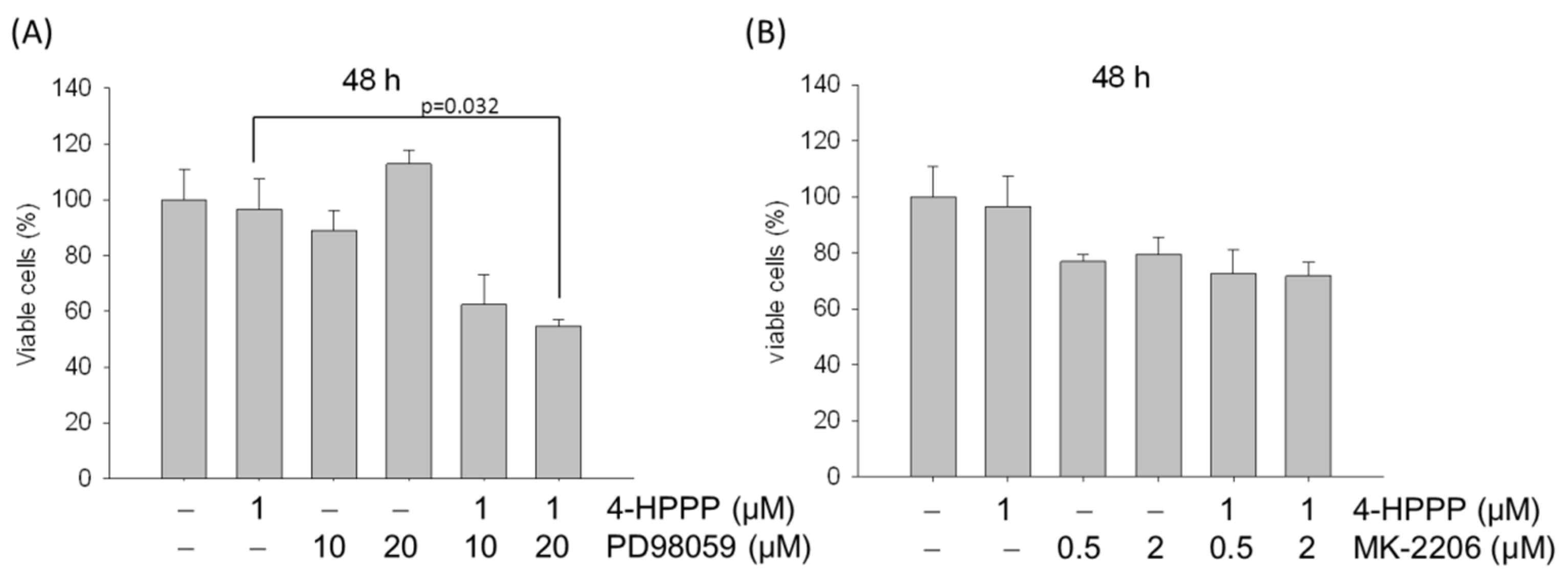
2.8. ERK Blockade Rescues 4-HPPP-Inhibition of Ha22T Cells
3. Discussion
4. Materials and Methods
4.1. Source of 4-[4-(4-Hydroxyphenoxy)phenoxy]phenol
4.2. Reagents
4.3. Cell Culture
4.4. Cell Growth Assessment
4.5. Colony Formation Assay
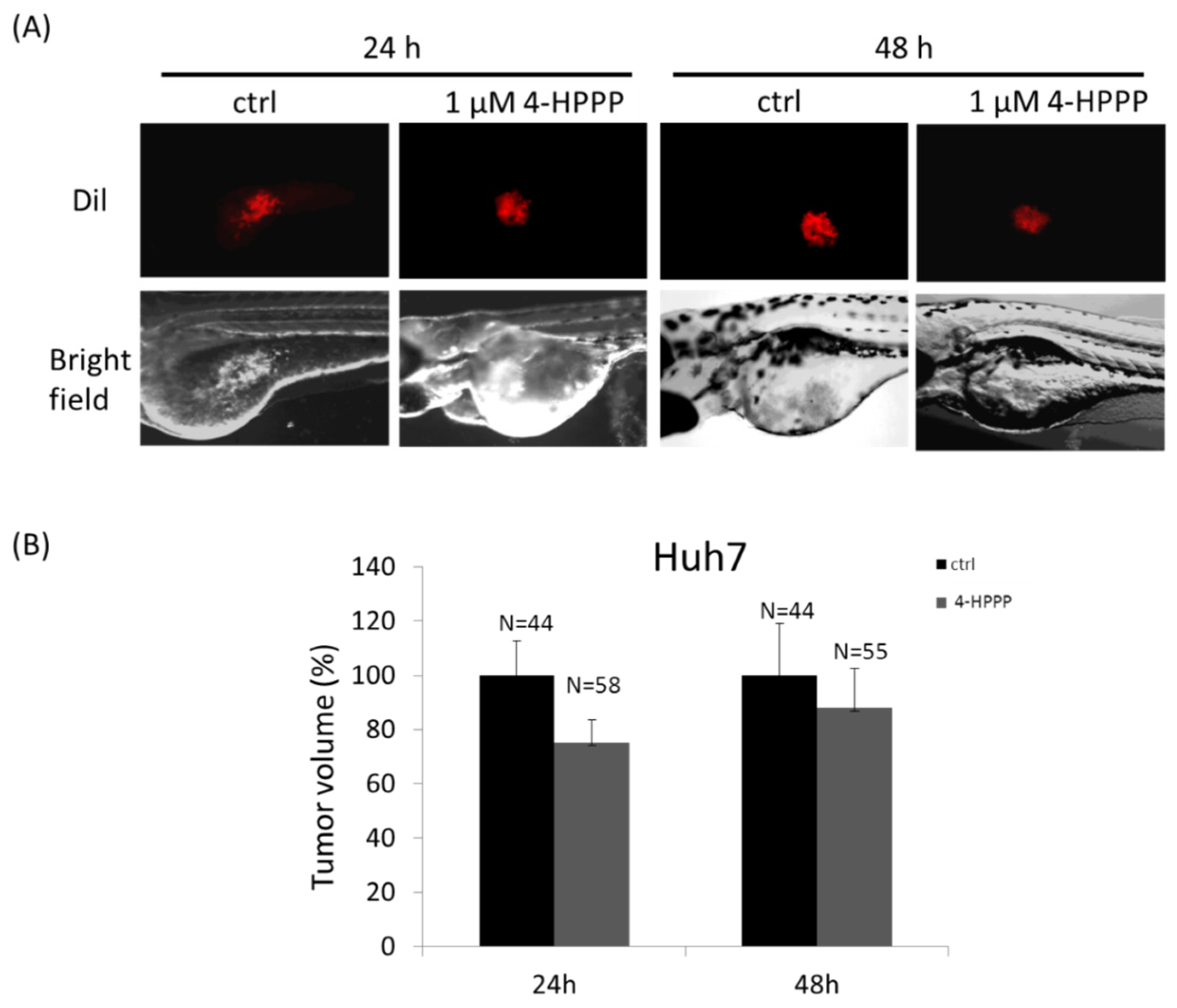
4.6. Zebrafish Xenograft Assay
4.7. DAPI Staining
4.8. The Assessment Apoptosis Assay
4.9. Acridine Orange (AO) Staining
4.10. Western Blotting Assay
4.11. Assessment of ERK and Akt Inhibition
4.12. Cell Cycle Distribution
4.13. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Akt | Protein kinase B |
| AO | Acridine orange |
| AVOs | Acidic vesicular organelles |
| Ctrl | Control |
| COX-2 | Cyclooxygenase-2 |
| CQ | Chloroquine |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| DiI | 1,1′-Dioctadecyl-3,3,3′,3′-Tetramethylindocarbocyanine Perchlorate |
| DMSO | Dimethyl sulfoxide |
| ERK | Extracellular signal-regulated kinase |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| γH2AX | Gamma-H2A histone family, member X |
| HCC | Hepatocellular carcinoma |
| 4-HPPP | 4-[4-(4-Hydroxyphenoxy)phenoxy]phenol |
| IC50 | Half-maximum inhibitory concentration |
| LC3B | Microtubule-associated protein-1 light chain-3B |
| MEK1 | Mitogen-activated protein or extracellular signal-regulated kinase kinase 1 |
| MMP | Matrix metalloproteinases |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| PCNA | Proliferating cell nuclear antigen |
| PI | Propidium iodide |
| PS | Phosphatidylserine |
| SD | Standard deviation |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.T.; Lo, C.M.; Guo, D.Y.; Qi, X.; Li, C.X.; Geng, W.; Liu, X.B.; Ling, C.C.; Ma, Y.Y.; Yeung, W.H.; et al. Identification of transmembrane protein 98 as a novel chemoresistance-conferring gene in hepatocellular carcinoma. Mol. Cancer Ther. 2014, 13, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Janke, C. The tubulin code: Molecular components, readout mechanisms, and functions. J. Cell Biol. 2014, 206, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Iskander, M.N. Microtubulin binding sites as target for developing anticancer agents. Mini Rev. Med. Chem. 2004, 4, 1077–1104. [Google Scholar] [CrossRef] [PubMed]
- Gasparotto, V.; Castagliuolo, I.; Ferlin, M.G. 3-substituted 7-phenyl-pyrroloquinolinones show potent cytotoxic activity in human cancer cell lines. J. Med. Chem. 2007, 50, 5509–5513. [Google Scholar] [CrossRef] [PubMed]
- Viola, G.; Bortolozzi, R.; Hamel, E.; Moro, S.; Brun, P.; Castagliuolo, I.; Ferlin, M.G.; Basso, G. MG-2477, a new tubulin inhibitor, induces autophagy through inhibition of the Akt/mTOR pathway and delayed apoptosis in A549 cells. Biochem. Pharmacol. 2012, 83, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Toma, C.C.; Olah, N.K.; Vlase, L.; Mogosan, C.; Mocan, A. Comparative Studies on Polyphenolic Composition, Antioxidant and Diuretic Effects of Nigella sativa L. (Black Cumin) and Nigella damascena L. (Lady-in-a-Mist) Seeds. Molecules 2015, 20, 9560–9574. [Google Scholar] [CrossRef] [PubMed]
- Dezsi, S.; Badarau, A.S.; Bischin, C.; Vodnar, D.C.; Silaghi-Dumitrescu, R.; Gheldiu, A.M.; Mocan, A.; Vlase, L. Antimicrobial and Antioxidant Activities and Phenolic Profile of Eucalyptus globulus Labill. and Corymbia ficifolia (F. Muell.) K.D. Hill & L.A.S. Johnson Leaves. Molecules 2015, 20, 4720–4734. [Google Scholar] [PubMed]
- Woo, K.W.; Kwon, O.W.; Kim, S.Y.; Choi, S.Z.; Son, M.W.; Kim, K.H.; Lee, K.R. Phenolic derivatives from the rhizomes of Dioscorea nipponica and their anti-neuroinflammatory and neuroprotective activities. J. Ethnopharmacol. 2014, 155, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Wahle, K.W.J.; Brown, I.; Rotondo, D.; Heys, S.D. Plant Phenolics in the Prevention and Treatment of Cancer. Adv. Exp. Med. Biol. 2010, 698, 36–51. [Google Scholar] [PubMed]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Fan, S.; Zhan, Q.; Kohn, K.W.; O’Connor, P.M.; Pommier, Y. Inactivation of p53 increases the cytotoxicity of camptothecin in human colon HCT116 and breast MCF-7 cancer cells. Clin. Cancer Res. 1997, 3, 1653–1660. [Google Scholar] [PubMed]
- Koizumi, N.; Hatano, E.; Nitta, T.; Tada, M.; Harada, N.; Taura, K.; Ikai, I.; Shimahara, Y. Blocking of PI3K/Akt pathway enhances apoptosis induced by SN-38, an active form of CPT-11, in human hepatoma cells. Int. J. Oncol. 2005, 26, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Bruchim, I.; Ben-Harim, Z.; Piura, E.; Haran, G.; Fishman, A. Analysis of two topotecan treatment schedules in patients with recurrent ovarian cancer. J. Chemother. 2016, 28, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Chou, H.L.; Chen, B.H.; Chang, K.F.; Tseng, C.H.; Fong, Y.; Fu, T.F.; Chang, H.W.; Wu, C.Y.; Tsai, E.M.; et al. BPIQ, a novel synthetic quinoline derivative, inhibits growth and induces mitochondrial apoptosis of lung cancer cells in vitro and in zebrafish xenograft model. BMC Cancer 2015, 15, 962. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Chen, Y.L.; Lu, P.J.; Yang, C.N.; Tzeng, C.C. Synthesis and antiproliferative evaluation of certain indeno[1,2-c]quinoline derivatives. Bioorg. Med. Chem. 2008, 16, 3153–3162. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.C.; Hung, C.T.; Chen, K.J.; Wu, W.C.; Suen, J.L.; Chang, C.H.; Lu, C.Y.; Tseng, C.H.; Chen, Y.L.; Chiu, C.C. Quinoline-Based Compound BPIQ Exerts Anti-Proliferative Effects on Human Retinoblastoma Cells via Modulating Intracellular Reactive Oxygen Species. Arch. Immunol. Ther. Exp. 2016, 64, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Kang, Y.J.; Kim, D.H.; Hossain, M.A.; Jang, J.Y.; Lee, S.H.; Yoon, J.H.; Chun, P.; Moon, H.R.; Kim, H.S.; et al. A novel hydroxamic acid derivative, MHY218, induces apoptosis and cell cycle arrest through downregulation of NF-kappa B in HCT116 human colon cancer cells. Int. J. Oncol. 2014, 44, 256–264. [Google Scholar] [PubMed]
- Lall, R.K.; Syed, D.N.; Adhami, V.M.; Khan, M.I.; Mukhtar, H. Dietary Polyphenols in Prevention and Treatment of Prostate Cancer. Int. J. Mol. Sci. 2015, 16, 3350–3376. [Google Scholar] [CrossRef] [PubMed]
- Parsai, S.; Keck, R.; Skrzypczak-Jankun, E.; Jankun, J. Analysis of the anticancer activity of curcuminoids, thiotryptophan and 4-phenoxyphenol derivatives. Oncol. Lett. 2014, 7, 17–22. [Google Scholar] [PubMed]
- Poon, R.T.; Fan, S.T. Hepatectomy for hepatocellular carcinoma: Patient selection and postoperative outcome. Liver Transpl. 2004, 10, S39–S45. [Google Scholar] [CrossRef] [PubMed]
- Chan-On, W.; Huyen, N.T.; Songtawee, N.; Suwanjang, W.; Prachayasittikul, S.; Prachayasittikul, V. Quinoline-based clioquinol and nitroxoline exhibit anticancer activity inducing FoxM1 inhibition in cholangiocarcinoma cells. Drug Des. Dev. Ther. 2015, 9, 2033–2047. [Google Scholar]
- Yu, J.; Taylor, K.E.; Zou, H.X.; Biswas, N.; Bewtra, J.K. Phenol Conversion and Dimeric Intermediates in Horseradish Peroxidase-Catalyzed Phenol Removal from Water. Environ. Sci. Technol. 1994, 28, 2154–2160. [Google Scholar] [CrossRef] [PubMed]
- Nepali, K.; Ojha, R.; Lee, H.Y.; Liou, J.P. Early investigational tubulin inhibitors as novel cancer therapeutics. Expert Opin. Investig. Drug 2016, 25, 917–936. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, E.S.; Heath, E.I.; Posadas, E.M.; Yu, E.Y.; Harrison, M.R.; Bruce, J.Y.; Cho, S.Y.; Wilding, G.E.; Fetterly, G.J.; Hangauer, D.G.; et al. A phase 2 study of KX2-391, an oral inhibitor of Src kinase and tubulin polymerization, in men with bone-metastatic castration-resistant prostate cancer. Cancer Chemother. Pharmacol. 2013, 71, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Meira, D.D.; de Almeida, V.H.; Mororo, J.S.; Nobrega, I.; Bardella, L.; Silva, R.L.A.; Albano, R.M.; Ferreira, C.G. Combination of cetuximab with chemoradiation, trastuzumab or MAPK inhibitors: Mechanisms of sensitisation of cervical cancer cells. Br. J. Cancer 2009, 101, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.-X.; Yin, X.-M. Chapter 20 Analyzing Macroautophagy in Hepatocytes and the Liver. In Methods Enzymology; Burlington Academic Press: Burlington, MA, USA, 2009; Volume 453, pp. 397–416. [Google Scholar]
- Piekarski, A.; Khaldi, S.; Greene, E.; Lassiter, K.; Mason, J.G.; Anthony, N.; Bottje, W.; Dridi, S. Tissue distribution, gender- and genotype-dependent expression of autophagy-related genes in avian species. PLoS ONE 2014, 9, e112449. [Google Scholar] [CrossRef] [PubMed]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.-A.; Outzen, H.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. p62/SQSTM1 Binds Directly to Atg8/LC3 to Facilitate Degradation of Ubiquitinated Protein Aggregates by Autophagy. J. Biol. Chem. 2007, 282, 24131–24145. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Yoshimori, T. How to Interpret LC3 Immunoblotting. Autophagy 2007, 3, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.M.; Liu, J.S.; Deng, L.J.; Chen, M.F.; Yiu, A.; Cao, H.H.; Tian, H.Y.; Fung, K.P.; Kurihara, H.; Pan, J.X.; et al. Arenobufagin, a natural bufadienolide from toad venom, induces apoptosis and autophagy in human hepatocellular carcinoma cells through inhibition of PI3K/Akt/mTOR pathway. Carcinogenesis 2013, 34, 1331–1342. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Feng, Y.B.; Zhu, M.F.; Tsang, C.M.; Man, K.; Tong, Y.; Tsao, S.W. Berberine Induces Autophagic Cell Death and Mitochondrial Apoptosis in Liver Cancer Cells: The Cellular Mechanism. J. Cell. Biochem. 2010, 111, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Miyazawa, K.; Naito, M.; Toyotake, J.; Tauchi, T.; Itoh, M.; Yuo, A.; Hayashi, Y.; Georgescu, M.M.; Kondo, Y.; et al. Vitamin K2 induces autophagy and apoptosis simultaneously in leukemia cells. Autophagy 2008, 4, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Choi, P.R.; Kang, Y.J.; Sung, B.; Kim, J.H.; Moon, H.R.; Chung, H.Y.; Kim, S.E.; Park, M.I.; Park, S.J.; Kim, N.D. MHY218-induced apoptotic cell death is enhanced by the inhibition of autophagy in AGS human gastric cancer cells. Int. J. Oncol. 2015, 47, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yang, J.; Huang, H. Oxidative stress induces H2AX phosphorylation in human spermatozoa. FEBS Lett. 2006, 580, 6161–6168. [Google Scholar] [CrossRef] [PubMed]
- Dickey, J.S.; Baird, B.J.; Redon, C.E.; Sokolov, M.V.; Sedelnikova, O.A.; Bonner, W.M. Intercellular communication of cellular stress monitored by γ-H2AX induction. Carcinogenesis 2009, 30, 1686–1695. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Juergens, S.; Zhang, X.; Rezaei, D.; Yu, C.; Strauch, E.D.; Wang, J.-Y.; Cheng, E.; Meyer, F.; Wang, D.H.; et al. Deoxycholic acid causes DNA damage while inducing apoptotic resistance through NF-κB activation in benign Barrett’s epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G278–G286. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Gopalakrishnan, V.; Hegde, M.; Kumar, S.; Karki, S.S.; Raghavan, S.C.; Choudhary, B. A Novel Resveratrol Based Tubulin Inhibitor Induces Mitotic Arrest and Activates Apoptosis in Cancer Cells. Sci. Rep. 2016, 6, 34653. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.; Jiang, J.-D.; Middleton, K.M.; Wang, Y.; Weisz, I.; Ling, Y.-H.; Bekesi, J.G. Novel Suicide Ligands of Tubulin Arrest Cancer Cells in S-Phase. Neoplasia 1999, 1, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Cheriyamundath, S.; Mahaddalkar, T.; Kantevari, S.; Lopus, M. Induction of acetylation and bundling of cellular microtubules by 9-(4-vinylphenyl) noscapine elicits S-phase arrest in MDA-MB-231 cells. Biomed. Pharmacother. 2017, 86, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, X.Y.; Shen, H.J.; Wang, D.X.; Wang, Y.H. Phosphorylated ERK is a potential predictor of sensitivity to sorafenib when treating hepatocellular carcinoma: Evidence from an in vitro study. BMC Med. 2009, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Wang, X.M.; Pan, C.; Ma, Q.M. Down-regulation of extracellular signal-regulated kinase 1/2 activity in P-glycoprotein-mediated multidrug resistant hepatocellular carcinoma cells. World J. Gastroenterol. 2009, 15, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Lin, Y.; TW, O.L.; Chou, C.K.; Lee, T.S.; Liu, T.J.; P’Eng, F.K.; Chen, T.Y.; Hu, C.P. Induction of plasma protein secretion in a newly established human hepatoma cell line. Mol. Cell. Biol. 1983, 3, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, H.; Taketa, K.; Yamane, T.; Miyazaki, M.; Miyano, K.; Sato, J. Phenotypical stability of a human hepatoma cell line, HuH-7, in long-term culture with chemically defined medium. Gan 1984, 75, 151–158. [Google Scholar] [PubMed]
- Lee, J.C.; Tseng, C.K.; Young, K.C.; Sun, H.Y.; Wang, S.W.; Chen, W.C.; Lin, C.K.; Wu, Y.H. Andrographolide exerts anti-hepatitis C virus activity by up-regulating haeme oxygenase-1 via the p38 MAPK/Nrf2 pathway in human hepatoma cells. Br. J. Pharmacol. 2014, 171, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Wen-Tsan, C.; Yao, F.; Shih-Chang, C.; Chon-Kit, C.; Han-Lin, C.; Chun-Feng, Y.; Chih-Hua, T.; Yeh-Long, C.; Chien-Chih, C. 9-bis[2-(pyrrolidin-1-yl)ethoxy]-6-{4-[2-(pyrrolidin-1-yl)ethoxy]phenyl}-11H-indeno[1,2-c]quinolin-11-one (BPIQ), a quinoline derivative inhibits human hepatocellular carcinoma cells by inducing ER stress and apoptosis. Anticancer Agents Med. Chem. 2016, 16, 1–9. [Google Scholar]
- Tzeng, C.H.; Chen, Y.R.; Tzeng, C.C.; Liu, W.T.; Chou, C.K.; Chiu, C.C.; Chen, Y.L. Discovery of indeno[1,2-b]quinoxaline derivatives as potential anticancer agents. Eur. J. Med. Chem. 2016, 108, 258–273. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Cheng, H.L.; Hsieh, B.S.; Chiu, C.C.; Lee, K.T.; Chang, K.L. Progesterone Increases Apoptosis and Inversely Decreases Autophagy in Human Hepatoma HA22T/VGH Cells Treated with Epirubicin. Sci. World J. 2014, 2014, 567148. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Pan, S.L.; Guh, J.H.; Chen, H.L.; Huang, D.M.; Chang, Y.L.; Kuo, S.C.; Lee, F.Y.; Teng, C.M. YC-1 [3-(5′-hydroxymethyl-2'-furyl)-1-benzyl indazole] exhibits a novel antiproliferative effect and arrests the cell cycle in G(0)-G(1) in human hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2005, 312, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Haung, J.W.; Chang, F.R.; Huang, K.J.; Huang, H.M.; Huang, H.W.; Chou, C.K.; Wu, Y.C.; Chang, H.W. Golden Berry-Derived 4 beta-hydroxywithanolide E for Selectively Killing Oral Cancer Cells by Generating ROS, DNA Damage, and Apoptotic Pathways. PLoS ONE 2013, 8, e64739. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compound 4-HPPPP is available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Phase | 24 h | 48 h | 4-HPPP (μM) | |||
|---|---|---|---|---|---|---|
| 0 | 10 | 0 | 10 | |||
| Huh7 | sub-G1 | 1.3 ± 0.2 | 4.4 ± 0.6 | 4.0 ± 0.6 | 13.1 ±6.9 | |
| G1 | 58.1 ± 1.8 | 74.9 ± 1.5 * | 50.9 ± 0.1 | 16.4 ± 0.2 * | ||
| S | 14.0 ± 0.4 | 15.0 ± 0.2 | 12.4 ± 0.3 | 40.1 ± 3.6 * | ||
| G2/M | 24.5 ± 1.6 | 6.9 ± 0.9 * | 19.5 ± 0.6 | 17.7 ± 1.6 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-T.; Liu, W.; Chiu, Y.-H.; Chen, B.-H.; Chuang, S.-C.; Chen, Y.-C.; Hsu, Y.-T.; Lu, M.-J.; Chiou, S.-J.; Chou, C.-K.; et al. A 4-Phenoxyphenol Derivative Exerts Inhibitory Effects on Human Hepatocellular Carcinoma Cells through Regulating Autophagy and Apoptosis Accompanied by Downregulating α-Tubulin Expression. Molecules 2017, 22, 854. https://doi.org/10.3390/molecules22050854
Chang W-T, Liu W, Chiu Y-H, Chen B-H, Chuang S-C, Chen Y-C, Hsu Y-T, Lu M-J, Chiou S-J, Chou C-K, et al. A 4-Phenoxyphenol Derivative Exerts Inhibitory Effects on Human Hepatocellular Carcinoma Cells through Regulating Autophagy and Apoptosis Accompanied by Downregulating α-Tubulin Expression. Molecules. 2017; 22(5):854. https://doi.org/10.3390/molecules22050854
Chicago/Turabian StyleChang, Wen-Tsan, Wangta Liu, Yi-Han Chiu, Bing-Hung Chen, Shih-Chang Chuang, Yen-Chun Chen, Yun-Tzh Hsu, Mei-Jei Lu, Shean-Jaw Chiou, Chon-Kit Chou, and et al. 2017. "A 4-Phenoxyphenol Derivative Exerts Inhibitory Effects on Human Hepatocellular Carcinoma Cells through Regulating Autophagy and Apoptosis Accompanied by Downregulating α-Tubulin Expression" Molecules 22, no. 5: 854. https://doi.org/10.3390/molecules22050854
APA StyleChang, W. -T., Liu, W., Chiu, Y. -H., Chen, B. -H., Chuang, S. -C., Chen, Y. -C., Hsu, Y. -T., Lu, M. -J., Chiou, S. -J., Chou, C. -K., & Chiu, C. -C. (2017). A 4-Phenoxyphenol Derivative Exerts Inhibitory Effects on Human Hepatocellular Carcinoma Cells through Regulating Autophagy and Apoptosis Accompanied by Downregulating α-Tubulin Expression. Molecules, 22(5), 854. https://doi.org/10.3390/molecules22050854