
Yeast Smell Like What They Eat: Analysis of Volatile Organic Compounds of Malassezia furfur in Growth Media Supplemented with Different Lipids

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
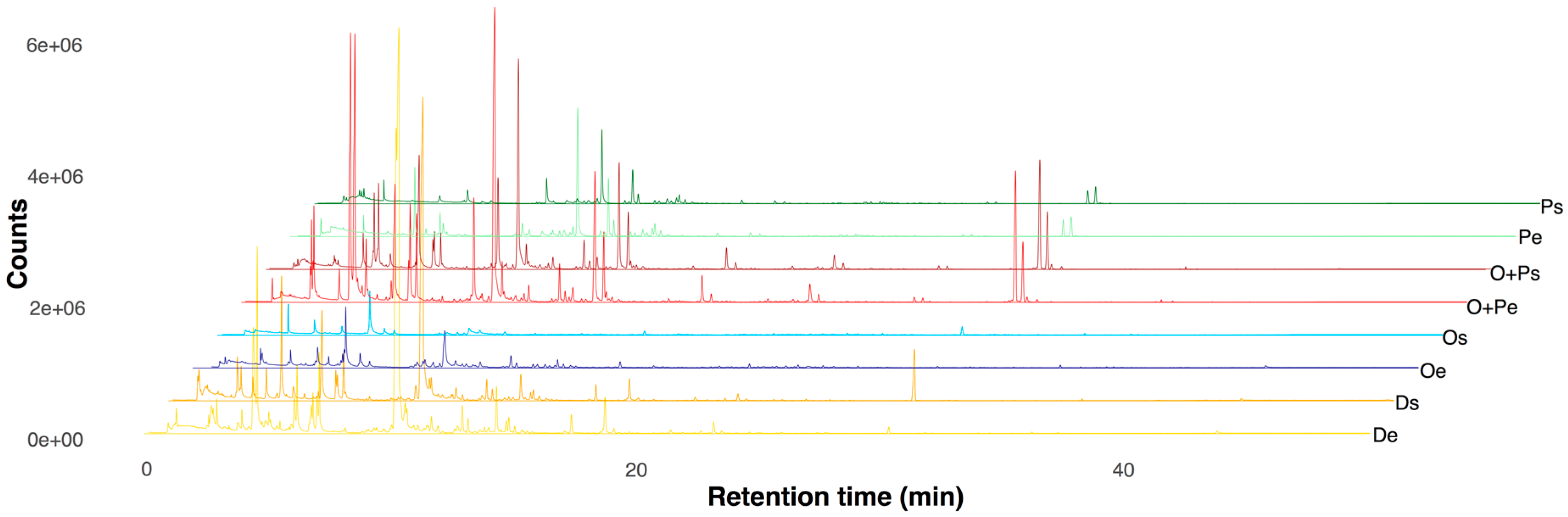
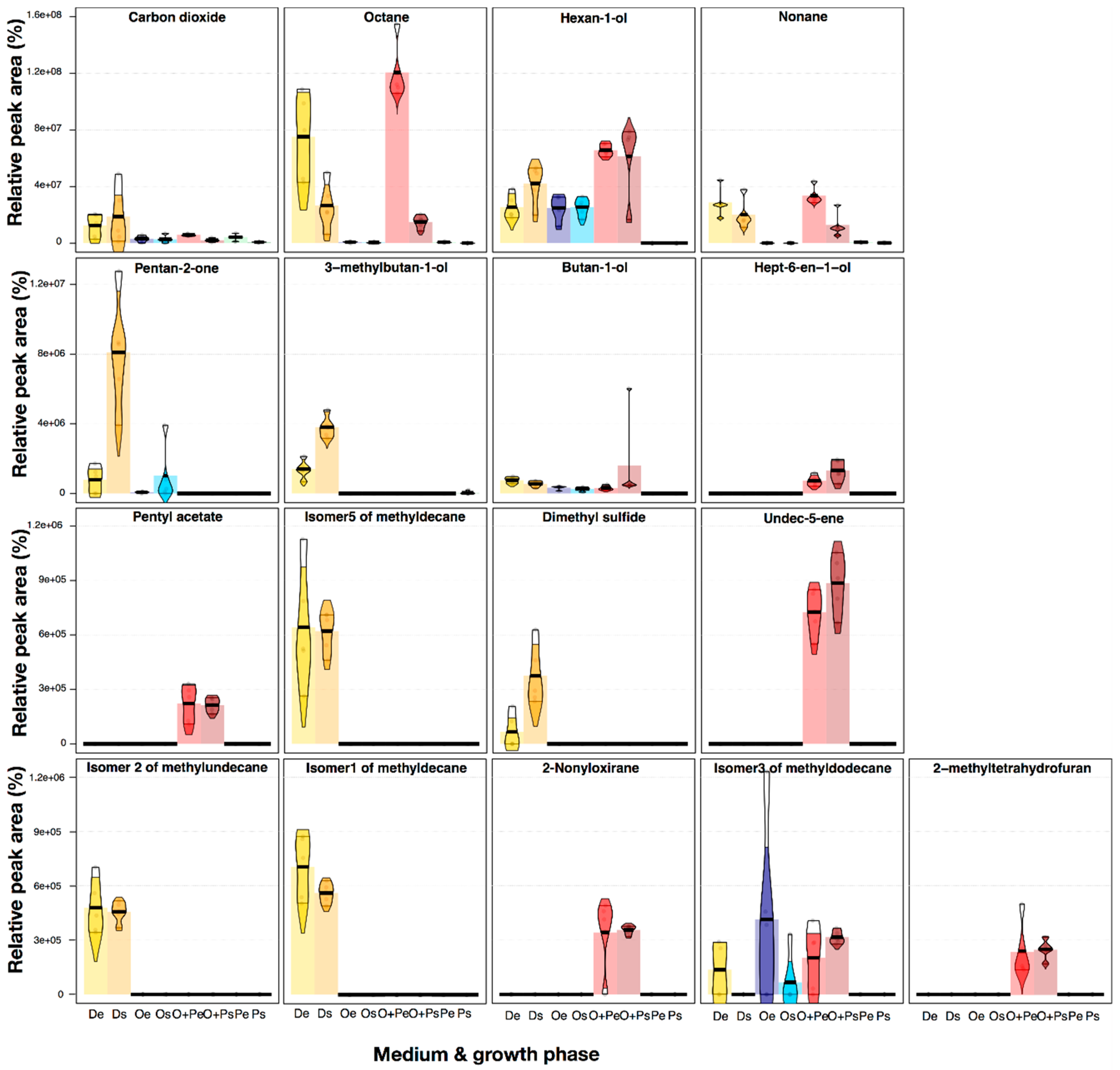
2.1. Profiling of VOCs from M. furfur
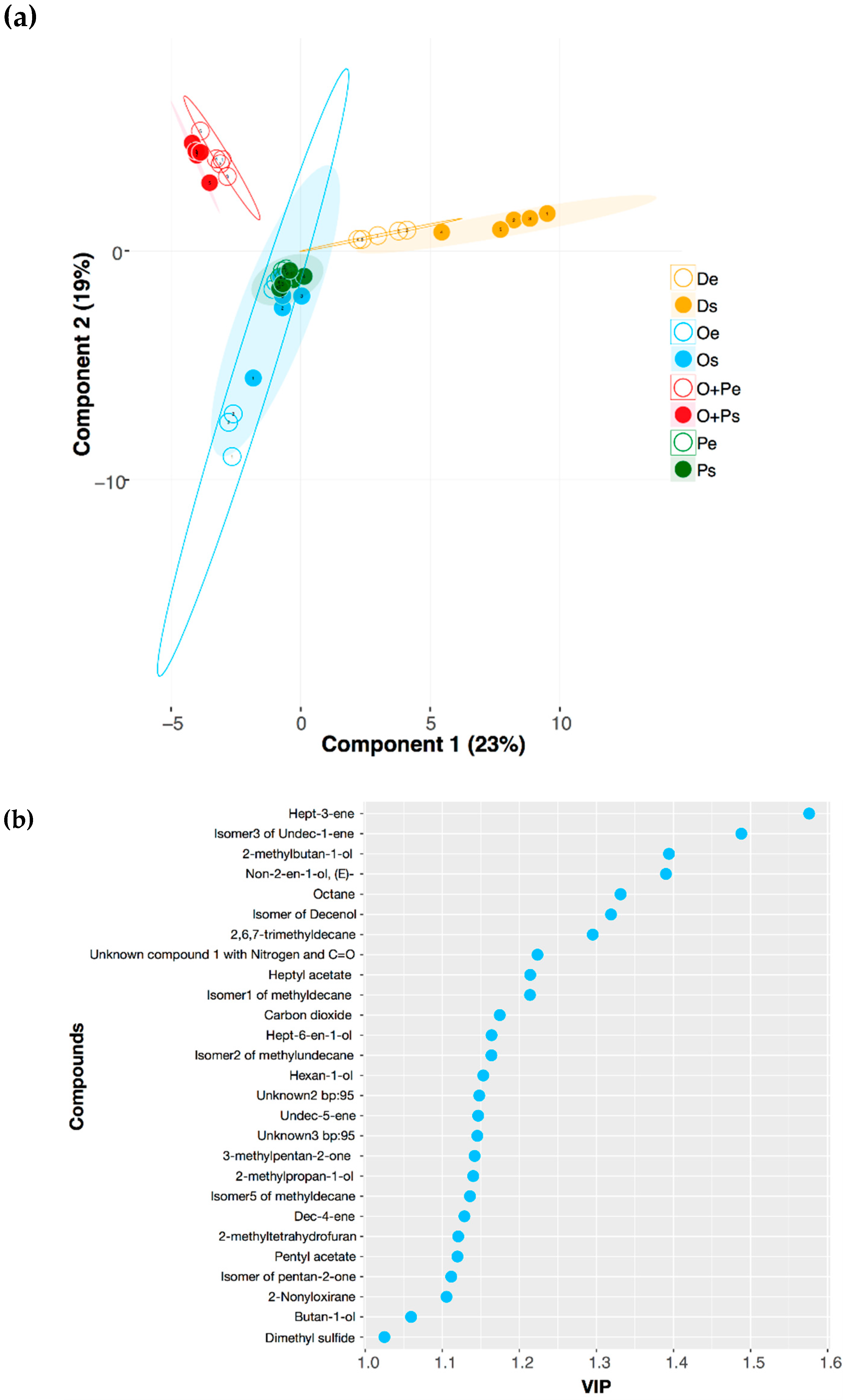
2.2. VOCs Profile Differentiation in Four Growth Media in the Exponential and Stationary Phase
3. Materials and Methods
3.1. Chemicals
3.2. Medium Preparation
3.3. Yeast Growth
3.4. HS-SPME/GC-MS Analysis
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shifrine, M.; Marr, A.G. The Requirement of fatty acids by Pityrosporum ovale. J. Gen. Microbiol. 1963, 32, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Saunders, C.W.; Hu, P.; Grant, R.A.; Boekhout, T.; Kuramae, E.E.; Kronstad, J.W.; DeAngelis, Y.M.; Reeder, N.L.; Johnstone, K.R.; et al. Dandruff-associated Malassezia genomes reveal convergent and divergent virulence traits shared with plant and human fungal pathogens. Proc. Natl. Acad. Sci. USA 2007, 104, 18730–18735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Zhao, H.; Li, C.; Rajapakse, M.P.; Wong, W.C.; Xu, J.; Saunders, C.W.; Reeder, N.L.; Reilman, R.A.; Scheynius, A.; et al. Genus-Wide comparative genomics of Malassezia delineates its phylogeny, physiology, and niche adaptation on human skin. PLoS Genet. 2015, 11, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Lorch, J.M.; Palmer, J.M.; Vanderwolf, K.J.; Schmidt, K.Z.; Verant, M.L.; Weller, T.J.; Blehert, D.S. Malassezia vespertilionis sp. nov.: A new cold-tolerant species of yeast isolated from bats. Persoonia—Mol. Phylogeny Evol. Fungi 2018, 41, 56–70. [Google Scholar] [CrossRef]
- Gao, Z.; Perez-Perez, G.I.; Chen, Y.; Blaser, M.J. Quantitation of major human cutaneous bacterial and fungal populations. J. Clin. Microbiol. 2010, 48, 3575–3581. [Google Scholar] [CrossRef] [PubMed]
- Ashbee, H.R.; Evans, E.G.V. Immunology of diseases associated with Malassezia species. Clin. Microbiol. Rev. 2002, 15, 21–57. [Google Scholar] [CrossRef] [PubMed]
- Mayser, P.A.; Lang, S.K. Pathogenicity of Malassezia yeasts. In The Mycota Human and Animal Relationships; Brakhage, A., Zipfel, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 115–141. ISBN 978-3-540-87406-5. [Google Scholar]
- Boekhout, T.; Mayser, P.; Guého-Kellermann, E.; Velegraki, A. Malassezia and the Skin. Science and Clinical Practice; Boekhout, T., Mayser, P., Guého-Kellermann, E., Velegraki, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 978-3-642-03615-6. [Google Scholar]
- Kesavan, S.; Holland, K.T.; Ingham, E. The effects of lipid extraction on the immunomodulatory activity of Malassezia species in vitro. Med. Mycol. 2000, 38, 239–247. [Google Scholar] [CrossRef] [Green Version]
- DeAngelis, Y.M.; Gemmer, C.M.; Kaczvinsky, J.R.; Kenneally, D.C.; Schwartz, J.R.; Dawson, T.L. Three etiologic facets of dandruff and seborrheic dermatitis: Malassezia fungi, sebaceous lipids, and individual sensitivity. J. Investig. Dermatol. Symp. Proc. 2005, 10, 295–297. [Google Scholar] [CrossRef]
- Bjerre, R.D.; Bandier, J.; Skov, L.; Engstrand, L.; Johansen, J.D. The role of the skin microbiome in atopic dermatitis: A systematic review. Br. J. Dermatol. 2017, 177, 1272–1278. [Google Scholar] [CrossRef]
- Li, H.; Goh, B.N.; Teh, W.K.; Jiang, Z.; Goh, J.P.Z.; Goh, A.; Wu, G.; Hoon, S.S.; Raida, M.; Camattari, A.; et al. Skin commensal Malassezia globosa secreted protease attenuates Staphylococcus aureus biofilm formation. J. Investig. Dermatol. 2018, 138, 1137–1145. [Google Scholar] [CrossRef]
- Cogen, A.L.; Nizet, V.; Gallo, R.L. Skin microbiota: A source of disease or defence? Br. J. Dermatol. 2008, 158, 442–455. [Google Scholar] [CrossRef] [PubMed]
- Bouslimani, A.; Porto, C.; Rath, C.M.; Wang, M.; Guo, Y.; Gonzalez, A.; Berg-Lyon, D.; Ackermann, G.; Moeller Christensen, G.J.; Nakatsuji, T.; et al. Molecular cartography of the human skin surface in 3D. Proc. Natl. Acad. Sci. USA 2015, 112, E2120–E2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, M.; Goldberg, D.; Aiello, A.; Larson, E.; Foxman, B. Skin microbiota: Microbial community structure and its potential association with health and disease. Infect. Genet. Evol. 2011, 11, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dréno, B.; Pécastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium acnes (Propionibacterium acnes) and acne vulgaris: A brief look at the latest updates. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Becker, J.; et al. Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Mayser, P.; Haze, P.; Papavassilis, C.; Pickel, M.; Gruender, K.; Gueho, E. Differentiation of Malassezia species: Selectivity of Cremophor EL, castor oil and ricinoleic acid for M. furfur. Br. J. Dermatol. 1997, 137, 208–213. [Google Scholar] [CrossRef]
- Wilde, P.F.; Stewart, P.S. A study of the fatty acid metabolism of the yeast Pityrosporum ovale. Biochem. J. 1968, 108, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Triana, S.; de Cock, H.; Ohm, R.A.; Danies, G.; Wösten, H.A.B.; Restrepo, S.; González Barrios, A.F.; Celis, A. Lipid metabolic versatility in Malassezia spp. yeasts studied through metabolic modeling. Front. Microbiol. 2017, 8, 1–18. [Google Scholar] [CrossRef]
- Kaneko, T.; Makimura, K.; Onozaki, M.; Ueda, K.; Yamada, Y.; Nishiyama, Y.; Yamaguchi, H. Vital growth factors of Malassezia species on modified CHROMagar Candida. Med. Mycol. 2005, 43, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Celis, A.M. Unraveling Lipid Metabolism in Lipid-Dependent Pathogenic Malassezia Yeasts. Ph.D. Thesis, Utrecht University, Utrecht, The Netherlands, 2017. [Google Scholar]
- Scotter, J.M.; Langford, V.S.; Wilson, P.F.; McEwan, M.J.; Chambers, S.T. Real-time detection of common microbial volatile organic compounds from medically important fungi by Selected Ion Flow Tube-Mass Spectrometry (SIFT-MS). J. Microbiol. Methods 2005, 63, 127–134. [Google Scholar] [CrossRef]
- Bazemore, R.A.; Feng, J.; Cseke, L.; Podila, G.K. Biomedically important pathogenic fungi detection with volatile biomarkers. J. Breath Res. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Heddergott, C.; Latgé, J.P.; Calvo, A.M. The volatome of Aspergillus fumigatus. Eukaryot. Cell 2014, 13, 1014–1025. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.; Thomas, H.R.; Daniels, S.D.; Lynch, R.C.; Fortier, S.M.; Shea, M.M.; Rearden, P.; Comolli, J.C.; Baden, L.R.; Marty, F.M. A breath fungal secondary metabolite signature to diagnose invasive aspergillosis. Clin. Infect. Dis. 2014, 59, 1733–1740. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhou, L.; Chen, H.; Wang, C.-Z.; Xia, Z.; Yuan, C.-S. Solid-phase microextraction technology for in vitro and in vivo metabolite analysis. Trends Analyt. Chem. 2016, 80, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.; Lee, N.; Seo, J.A.; Kim, Y.S. Comparative analysis of nonvolatile and volatile metabolites in Lichtheimia ramosa cultivated in different growth media. Biosci. Biotechnol. Biochem. 2017, 81, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Rees, C.A.; Burklund, A.; Stefanuto, P.H.; Schwartzman, J.D.; Hill, J.E. Comprehensive volatile metabolic fingerprinting of bacterial and fungal pathogen groups. J. Breath Res. 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Hung, R.; Lee, S.; Bennett, J.W. Fungal volatile organic compounds and their role in ecosystems. Appl. Microbiol. Biotechnol. 2015, 99, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Tejero Rioseras, A.; Garcia Gomez, D.; Ebert, B.E.; Blank, L.M.; Ibáñez, A.J.; Sinues, P.M.L. Comprehensive real-time analysis of the yeast Volatilome. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Romero-Guido, C.; Belo, I.; Ta, T.M.N.; Cao-Hoang, L.; Alchihab, M.; Gomes, N.; Thonart, P.; Teixeira, J.A.; Destain, J.; Waché, Y. Biochemistry of lactone formation in yeast and fungi and its utilisation for the production of flavour and fragrance compounds. Appl. Microbiol. Biotechnol. 2011, 89, 535–547. [Google Scholar] [CrossRef]
- Braga, A.; Belo, I. Biotechnological production of γ-decalactone, a peach like aroma, by Yarrowia lipolytica. World J. Microbiol. Biotechnol. 2016, 32, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, R.E. The consequences of volatile organic compound mediated bacterial and fungal interactions. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2002, 81, 357–364. [Google Scholar] [CrossRef]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Hung, R.; Lee, S.; Padhi, S. Fungal and bacterial Volatile Organic Compounds: An overview and their role as ecological signaling agents. In Fungal Associations; Springer: Berlin/Heidelberg, Germany, 2012; Volume 9, pp. 373–393. ISBN 9783642308260. [Google Scholar]
- Lemfack, M.C.; Nickel, J.; Dunkel, M.; Preissner, R.; Piechulla, B. mVOC: A database of microbial volatiles. Nucleic Acids Res. 2014, 42, D744–D748. [Google Scholar] [CrossRef]
- Lemfack, M.C.; Gohlke, B.O.; Toguem, S.M.T.; Preissner, S.; Piechulla, B.; Preissner, R. MVOC 2.0: A database of microbial volatiles. Nucleic Acids Res. 2018, 46, D1261–D1265. [Google Scholar] [CrossRef]
- Labows, J.N.; McGinley, K.J.; Leyden, J.J.; Webster, G.F. Characteristic γ-lactone odor production of the genus Pityrosporum. Appl. Environ. Microbiol. 1979, 38, 412–415. [Google Scholar]
- Ro, B.I.; Dawson, T.L. The role of sebaceous gland activity and scalp microfloral metabolism in the etiology of seborrheic dermatitis and dandruff. J. Investig. Dermatol. Symp. Proc. 2005, 10, 194–197. [Google Scholar] [CrossRef]
- Bruce, A.; Wheatley, R.E.; Humphris, S.N.; Hackett, C.A.; Florence, M.E.J. Production of volatile organic compounds by Trichoderma in media containing different amino acids and their effect on selected wood decay fungi. Holzforschung 2000, 54, 481–486. [Google Scholar] [CrossRef]
- Sunesson, A.L.; Vaes, W.H.J.; Nilsson, C.A.; Blomquist, G.; Andersson, B.; Carlson, R. Identification of volatile metabolites from five fungal species cultivated on two media. Appl. Environ. Microbiol. 1995, 61, 2911–2918. [Google Scholar]
- Langford, M.L.; Atkin, A.L.; Nickerson, K.W. Cellular interactions of farnesol, a quorum-sensing molecule produced by Candida albicans. Future Microbiol. 2009, 4, 1353–1362. [Google Scholar] [CrossRef]
- Kramer, R.; Abraham, W.R. Volatile sesquiterpenes from fungi: What are they good for? Phytochem. Rev. 2012, 11, 15–37. [Google Scholar] [CrossRef]
- Thalavitiya Acharige, M.J.; Koshy, S.S.; Koo, S. The use of microbial metabolites for the diagnosis of infectious diseases. In Advanced Techniques in Diagnostic Microbiology; Springer International Publishing: Cham, Switzerland, 2018; pp. 261–272. ISBN 9783319339009. [Google Scholar]
- Lin, H.C.; Chooi, Y.H.; Dhingra, S.; Xu, W.; Calvo, A.M.; Tang, Y. The fumagillin biosynthetic gene cluster in Aspergillus fumigatus encodes a cryptic terpene cyclase involved in the formation of β-trans-bergamotene. J. Am. Chem. Soc. 2013, 135, 4616–4619. [Google Scholar] [CrossRef] [PubMed]
- Ashbee, H.R. Update on the genus Malassezia. Med. Mycol. 2007, 45, 287–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weimer, B.; Seefeldt, K.; Dias, B. Sulfur metabolism in bacteria associated with cheese. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 1999, 76, 247–261. [Google Scholar] [CrossRef]
- Schöller, C.E.G.; Gürtler, H.; Pedersen, R.; Molin, S.; Wilkins, K. Volatile metabolites from Actinomycetes. J. Agric. Food Chem. 2002, 50, 2615–2621. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R. Flavors and aromas. In Fungal Biotechnology In Agricultural, Food and Environmental Applications; Arora, D., Ed.; Marcel Dekker Inc.: New York, NY, USA, 2004; pp. 281–289. ISBN 3527403736. [Google Scholar]
- Buzzini, P.; Romano, S.; Turchetti, B.; Vaughan, A.; Pagnoni, U.M.; Davoli, P. Production of volatile organic sulfur compounds (VOSCs) by basidiomycetous yeasts. FEMS Yeast Res. 2005, 5, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Filipiak, W.; Sponring, A.; Filipiak, A.; Baur, M.; Ager, C.; Wiesenhofer, H.; Margesin, R.; Nagl, M.; Troppmair, J.; Amann, A. Volatile Organic Compounds (VOCs) released by pathogenic microorganisms in vitro: Potential breath biomarkers for early-stage diagnosis of disease. In Volatile Biomarkers; Elsevier: Amsterdam, The Netherlands, 2013; pp. 463–512. ISBN 9780444626134. [Google Scholar]
- Sprecher, E. Influence of strain specificity and culture conditions on terpene production by fungi. Planta Med. 1982, 44, 41–43. [Google Scholar] [CrossRef]
- Bjurman, J.; Kristensson, J. Volatile production by Aspergillus versicolor as a possible cause of odor in houses affected by fungi. Mycopathologia 1992, 118, 173–178. [Google Scholar] [CrossRef]
- Gethins, L.; Guneser, O.; Demirkol, A.; Rea, M.C.; Stanton, C.; Ross, R.P.; Karagul Yuceer, Y.; Morrissey, J.P. Influence of Carbon and Nitrogen source on production of volatile fragrance and flavour metabolites by the yeast Kluyveromyces marxianus. Yeast 2015, 32, 67–76. [Google Scholar] [CrossRef]
- Korpi, A.; Järnberg, J.; Pasanen, A.L. Microbial volatile organic compounds. Crit. Rev. Toxicol. 2009, 39, 139–193. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; De Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Thorn, R.M.S.; Reynolds, D.M.; Greenman, J. Multivariate analysis of bacterial volatile compound profiles for discrimination between selected species and strains in vitro. J. Microbiol. Methods 2011, 84, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Bos, L.D.J.; Sterk, P.J.; Schultz, M.J. Volatile Metabolites of Pathogens: A Systematic Review. PLoS Pathog. 2013, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boots, A.W.; Smolinska, A.; Van Berkel, J.J.B.N.; Fijten, R.R.R.; Stobberingh, E.E.; Boumans, M.L.L.; Moonen, E.J.; Wouters, E.F.M.; Dallinga, J.W.; Van Schooten, F.J. Identification of microorganisms based on headspace analysis of volatile organic compounds by gas chromatography-mass spectrometry. J. Breath Res. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Rees, C.A.; Smolinska, A.; Hill, J.E. The volatile metabolome of Klebsiella pneumoniae in human blood. J. Breath Res. 2016, 10, 27101. [Google Scholar] [CrossRef] [PubMed]
- Tabares, M.; Ortiz, M.; Gonzalez, M.; Carazzone, C.; Vives Florez, M.J.; Molina, J. Behavioral responses of Rhodnius prolixus to volatile organic compounds released in vitro by bacteria isolated from human facial skin. PLoS Negl. Trop. Dis. 2018, 12, 1–16. [Google Scholar] [CrossRef]
- Kladsomboon, S.; Thippakorn, C.; Seesaard, T. Development of organic-inorganic hybrid optical gas sensors for the non-invasive monitoring of pathogenic bacteria. Sensors 2018, 18, 3189. [Google Scholar] [CrossRef]
- Sun, D.; She, J.; Gower, J.; Stokes, C.; Windham, G.; Baird, R.; Mlsna, T. Effects of growth parameters on the analysis of Aspergillus flavus volatile metabolites. Separations 2016, 3, 13. [Google Scholar] [CrossRef]
- Casu, F.; Pinu, F.R.; Fedrizzi, B.; Greenwood, D.R.; Villas-Boas, S.G. The effect of linoleic acid on the Sauvignon blanc fermentation by different wine yeast strains. FEMS Yeast Res. 2016, 16, fow050. [Google Scholar] [CrossRef]
- Gromski, P.S.; Muhamadali, H.; Ellis, D.I.; Xu, Y.; Correa, E.; Turner, M.L.; Goodacre, R. A tutorial review: Metabolomics and partial least squares-discriminant analysis—A marriage of convenience or a shotgun wedding. Anal. Chim. Acta 2015, 879, 10–23. [Google Scholar] [CrossRef]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma. II. Production of volatile antibiotics. Trans. Br. Mycol. Soc. 1971, 57, 41-IN4. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Triba, M.N.; Le Moyec, L.; Amathieu, R.; Goossens, C.; Bouchemal, N.; Nahon, P.; Rutledge, D.N.; Savarin, P. PLS/OPLS models in metabolomics: The impact of permutation of dataset rows on the K-fold cross-validation quality parameters. Mol. Biosyst. 2015, 11, 13–19. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of M. furfur CBS 1878 are currently available at the “Grupo de Investigación Celular de Micoorganismos Patógenos (CeMoP)” at Universidad de los Andes. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | De | Ds | Oe | Os | Pe | Ps | OPe | OPs | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | Crit. | RI | RI Exp | Ret. Time | X | SD | N | X | SD | N | X | SD | N | X | SD | N | X | SD | N | X | SD | N | X | SD | N | X | SD | N |
| Carbon dioxide * | a | 1.3 | 7.7 | 4.2 | 5 | 14.6 | 15.8 | 5 | 9.8 | 6.6 | 5 | 8.4 | 6.2 | 5 | 61.5 | 21.4 | 5 | 45.3 | 22.0 | 5 | 2.5 | 0.4 | 5 | 2.4 | 1.7 | 5 | ||
| Pentane * | a | 500 | 1.6 | 1.1 | 1 | 2.0 | 0.3 | 2 | 0.1 | 0.1 | 2 | |||||||||||||||||
| Dimethyl sulphide * | a | 526 | 1.7 | 0.1 | 0.1 | 2 | 0.3 | 0.1 | 5 | |||||||||||||||||||
| 2-methylpent-1-ene | a | 588 | 2.0 | 0.2 | 0.2 | 3 | 0.3 | 1 | ||||||||||||||||||||
| 2-methylpropan-1-ol * | a,b | 629 | 624 | 2.2 | 0.1 | 1 | 0.2 | 0.1 | 5 | |||||||||||||||||||
| Butan-1-ol * | a,b | 663 | 658 | 2.6 | 0.5 | 0.2 | 5 | 0.4 | 0.1 | 5 | 1.0 | 0.3 | 5 | 0.8 | 0.3 | 5 | 0.1 | 0.1 | 5 | 2.1 | 3.5 | 5 | ||||||
| 2-methyltetrahydrofuran | a,b | 678 | 665 | 2.6 | 0.1 | 0.1 | 5 | 0.3 | 0.0 | 5 | ||||||||||||||||||
| Isomer of pentan-2-one | a | 678 | 2.8 | 0.6 | 0.3 | 4 | ||||||||||||||||||||||
| Pentan-2-one * | a,b | 674 | 684 | 2.8 | 0.8 | 0.2 | 3 | 6.2 | 2.0 | 5 | 0.2 | 0.1 | 4 | 3.6 | 5.6 | 5 | ||||||||||||
| Hept-2-ene | a,b | est: 725 | 702 | 3.0 | 0.6 | 1.0 | 4 | 2.4 | 1 | |||||||||||||||||||
| Hept-3-ene | a,b | 730 | 713 | 3.2 | 0.1 | 0.1 | 5 | |||||||||||||||||||||
| 2,5-dimethylhexane | a | est: 688 | 723 | 3.4 | 2.4 | 0.4 | 3 | |||||||||||||||||||||
| 3-methylbutan-1-ol * | a,b | 746 | 730 | 3.5 | 0.9 | 0.2 | 5 | 3.0 | 0.5 | 5 | 11.9 | 1 | ||||||||||||||||
| 2-methylbutan-1-ol * | a,b | 745 | 734 | 3.5 | 0.1 | 0.0 | 5 | 0.2 | 0.1 | 2 | ||||||||||||||||||
| 3-methylpentan-2-one * | a,b | 752 | 748 | 3.8 | 0.1 | 0.0 | 4 | |||||||||||||||||||||
| Octane * | a,b | 800 | 800 | 4.6 | 49.3 | 11.4 | 5 | 20.1 | 10.9 | 5 | 2.8 | 1.4 | 4 | 1.6 | 1.3 | 5 | 11.6 | 10.2 | 5 | 4.8 | 1 | 51.9 | 2.7 | 5 | 15.3 | 3.7 | 5 | |
| Oct-2-ene * | a,b | 810 | 806 | 4.8 | 0.2 | 1 | 0.2 | 0.1 | 3 | |||||||||||||||||||
| 2-Furanmethanol | a,b | 858 | 852 | 5.9 | 0.3 | 0.0 | 2 | 0.4 | 0.2 | 5 | ||||||||||||||||||
| Hexan-1-ol * | a,b | 870 | 865 | 6.3 | 17.9 | 7.2 | 5 | 32.9 | 11.8 | 5 | 72.9 | 12.6 | 5 | 80.5 | 6.3 | 5 | 28.6 | 3.2 | 5 | 60.0 | 21.4 | 5 | ||||||
| Nonane | a,b | 900 | 900 | 7.2 | 19.7 | 7.2 | 5 | 15.6 | 8.1 | 5 | 0.8 | 1 | 0.7 | 0.3 | 2 | 9.3 | 1.3 | 5 | 11.9 | 10.6 | 5 | 14.4 | 1.0 | 5 | 14.5 | 12.8 | 5 | |
| Pentyl acetate * | a,b | est: 924 | 912 | 7.6 | 0.1 | 0.0 | 5 | 0.2 | 0.1 | 5 | ||||||||||||||||||
| Unknown bp: 56 | 925 | 8.0 | 0.1 | 0.0 | 4 | 0.1 | 0.0 | 2 | ||||||||||||||||||||
| Unknown compound 1 with Nitrogen and C=O | a | 955 | 9.1 | 0.0 | 0.0 | 3 | ||||||||||||||||||||||
| Hept-6-en-1-ol | a,b | est: 950 | 959 | 9.3 | 0.3 | 0.1 | 5 | 1.3 | 0.5 | 5 | ||||||||||||||||||
| Unknown1 bp: 95 | 980 | 10.0 | 0.1 | 0.0 | 3 | 0.1 | 0.0 | 2 | ||||||||||||||||||||
| Decane | a,b | 1000 | 1000 | 10.7 | 0.7 | 0.1 | 3 | |||||||||||||||||||||
| Dec-4-ene | a,b | est: 1023 | 1005 | 10.9 | 0.3 | 0.0 | 3 | |||||||||||||||||||||
| Isomer1 of methyldecane | a | 1001 | 1038 | 12.3 | 0.5 | 0.2 | 5 | 0.4 | 0.1 | 5 | ||||||||||||||||||
| 2,6,7-trimethyldecane | a,b | est: 1048 | 1052 | 12.9 | 1.9 | 0.4 | 5 | |||||||||||||||||||||
| Isomer4 of methyldecane | a | 1059 | 13.2 | 0.3 | 0.1 | 3 | 0.2 | 0.0 | 3 | |||||||||||||||||||
| Isomer5 of methyldecane | a | 1069 | 13.6 | 0.5 | 0.4 | 5 | 0.5 | 0.1 | 5 | |||||||||||||||||||
| (5-Ethyl-cyclopent-1-enyl)-methanol | a,b | 1073 | 1073 | 13.7 | 0.8 | 0.4 | 3 | 1.2 | 1 | |||||||||||||||||||
| Isomer1 of Undec-1-ene | a | 1079 | 14.0 | 1.9 | 0.9 | 3 | 1.1 | 0.1 | 3 | |||||||||||||||||||
| Isomer2 of Undec-1-ene | a | 1083 | 14.2 | 8.4 | 3.8 | 3 | 2.0 | 1.2 | 3 | |||||||||||||||||||
| Undec-5-ene | a,b | 1092 | 1105 | 15.1 | 0.3 | 0.0 | 5 | 0.9 | 0.2 | 5 | ||||||||||||||||||
| Heptyl acetate | a,b | 1110 | 1112 | 15.4 | 0.1 | 0.0 | 3 | |||||||||||||||||||||
| Isomer3 of Undec-1-ene | a | 1114 | 15.5 | 1.5 | 0.4 | 5 | ||||||||||||||||||||||
| Isomer1 of methylundecane | a | 1124 | 15.9 | 7.8 | 3.3 | 2 | ||||||||||||||||||||||
| Isomer2 of methylundecane | a | 1124 | 15.9 | 0.4 | 0.2 | 5 | 0.4 | 0.0 | 5 | |||||||||||||||||||
| Non-3-en-1-ol, (Z)- * | a,b | 1158 | 1148 | 17.0 | 0.2 | 1 | 0.1 | 0.1 | 3 | |||||||||||||||||||
| Non-2-en-1-ol, (E)- * | a | 1120 | 1167 | 17.8 | 0.3 | 0.2 | 4 | |||||||||||||||||||||
| Unknown2 bp:95 | 1181 | 18.5 | 0.3 | 0.0 | 4 | |||||||||||||||||||||||
| 2-Nonyloxirane | a,b | est: 1205 | 1210 | 19.7 | 0.2 | 0.0 | 4 | 0.4 | 0.1 | 5 | ||||||||||||||||||
| 4,7-dimethylundecane | a,b | est: 1185 | 1211 | 19.8 | 0.3 | 0.0 | 2 | 5.4 | 1.3 | 4 | 11.7 | 3.8 | 2 | |||||||||||||||
| Isomer2 of methyldodecane | a | 1237 | 20.9 | 1.2 | 0.6 | 3 | 0.8 | 1 | ||||||||||||||||||||
| Isomer3 of methyldodecane | a | 1245 | 21.3 | 0.2 | 0.0 | 2 | 1.8 | 0.6 | 3 | 0.9 | 1 | 0.1 | 0.1 | 4 | 0.3 | 0.1 | 5 | |||||||||||
| Isomer of Decenol | a | 1268 | 22.3 | 0.4 | 0.3 | 4 | ||||||||||||||||||||||
| Isomer5 of methyldodecane | a | 1274 | 22.6 | 0.6 | 0.7 | 2 | ||||||||||||||||||||||
| Undecan-2-one * | a,b | 1294 | 1292 | 23.4 | 16.7 | 8.7 | 4 | |||||||||||||||||||||
| Isomer6 of methyldodecane | a | 1295 | 23.5 | 1.0 | 0.0 | 2 | 1.5 | 1 | ||||||||||||||||||||
| Isomer2 of methyldecanol | a | 1295 | 23.5 | 4.7 | 2.2 | 2 | 12.2 | 2.0 | 2 | |||||||||||||||||||
| Unknown furanic | a | 1301 | 23.7 | 0.7 | 0.5 | 2 | ||||||||||||||||||||||
| Isomer1 of trimethyldodecane | a | 1302 | 23.8 | 2.2 | 0.7 | 3 | 1.2 | 0.2 | 2 | |||||||||||||||||||
| Isomer2 of trimethyldodecane | a | 1320 | 24.6 | 1.0 | 0.2 | 3 | 0.7 | 1 | ||||||||||||||||||||
| Isomer of methylundecanol | a | 1323 | 24.7 | 1.8 | 0.5 | 3 | 1.0 | 1 | ||||||||||||||||||||
| Isomer1 of methyltridecane | a | 1336 | 25.3 | 0.5 | 0.2 | 4 | 1.6 | 1.2 | 3 | |||||||||||||||||||
| Isomer2 of methyltridecane | a | 1343 | 25.6 | 1.2 | 0.4 | 3 | 0.8 | 1 | ||||||||||||||||||||
| Isomer3 of methyltridecane | a | 1354 | 26.0 | 0.8 | 0.0 | 2 | ||||||||||||||||||||||
| Unknown3 bp:95 | 1381 | 27.2 | 0.3 | 0.1 | 4 | |||||||||||||||||||||||
| Isomer of methyltetradecane | a | 1496 | 31.9 | 1.2 | 0.4 | 3 | ||||||||||||||||||||||
| γ-Dodecalactone | a,b | 1682 | 1679 | 37.5 | 0.3 | 0.2 | 3 | 0.4 | 0.1 | 4 | 0.6 | 1 | 0.9 | 0.1 | 3 | |||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez, M.; Celis, A.M.; Guevara-Suarez, M.I.; Molina, J.; Carazzone, C. Yeast Smell Like What They Eat: Analysis of Volatile Organic Compounds of Malassezia furfur in Growth Media Supplemented with Different Lipids. Molecules 2019, 24, 419. https://doi.org/10.3390/molecules24030419
Gonzalez M, Celis AM, Guevara-Suarez MI, Molina J, Carazzone C. Yeast Smell Like What They Eat: Analysis of Volatile Organic Compounds of Malassezia furfur in Growth Media Supplemented with Different Lipids. Molecules. 2019; 24(3):419. https://doi.org/10.3390/molecules24030419
Chicago/Turabian StyleGonzalez, Mabel, Adriana M. Celis, Marcela I. Guevara-Suarez, Jorge Molina, and Chiara Carazzone. 2019. "Yeast Smell Like What They Eat: Analysis of Volatile Organic Compounds of Malassezia furfur in Growth Media Supplemented with Different Lipids" Molecules 24, no. 3: 419. https://doi.org/10.3390/molecules24030419
APA StyleGonzalez, M., Celis, A. M., Guevara-Suarez, M. I., Molina, J., & Carazzone, C. (2019). Yeast Smell Like What They Eat: Analysis of Volatile Organic Compounds of Malassezia furfur in Growth Media Supplemented with Different Lipids. Molecules, 24(3), 419. https://doi.org/10.3390/molecules24030419