Transcriptome Analysis Clarified Genes Involved in Betalain Biosynthesis in the Fruit of Red Pitayas (Hylocereus costaricensis)

Abstract
:1. Introduction
2. Results
2.1. Illumina Sequencing and Unigenes Assembly
2.2. Differentially Expressed Unigenes in Red and White Flesh
2.3. Unigenes in the Betalain Biosynthesis Pathway and Their Expression Character
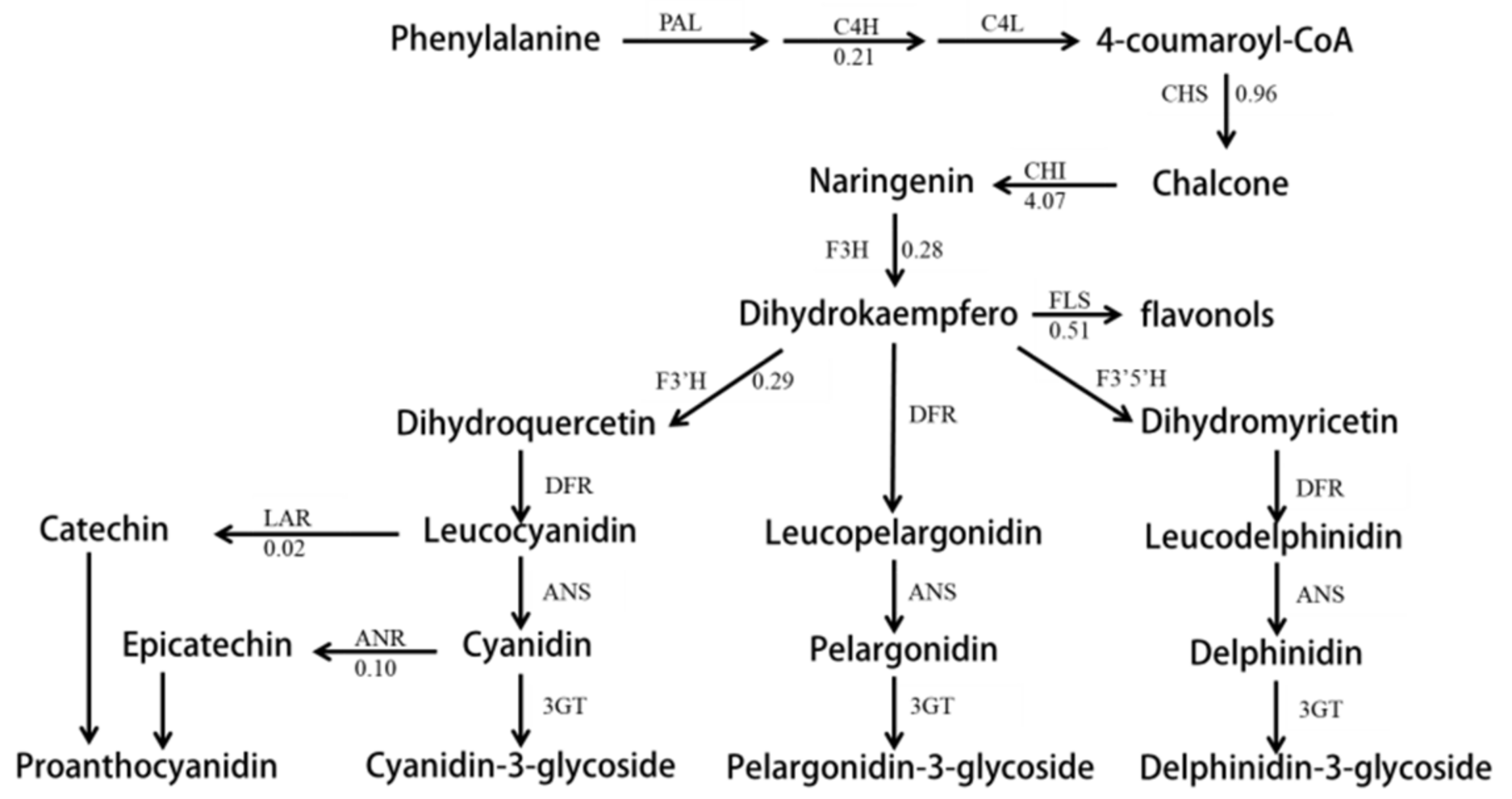
2.4. The Expression Character of the Unigenes Relative to Anthocyanin Biosynthesis
2.5. The Key MYB Regulation Factor Regulating Betalain Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Plant Materials, cDNA Preparation, and Illumina Sequencing
4.2. Sequence Data Filtering and De Novo Assembly
4.3. Analysis of Differential Gene Expression
4.4. Gene Annotation and Classification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nerd, A.; Sitrit, Y.; Kaushik, R.A.; Mizrahi, Y. High summer temperatures inhibit flowering in vine pitaya crops (Hylocereus spp.). Sci. Hortic. 2002, 96, 343–350. [Google Scholar] [CrossRef]
- Ortiz-Hernandez, Y.D. Pitahaya: A New Crop for Mexico (Pitahaya: Un Nuevo Cultivo Para Mexico); Limusa—Grupo Noriega Editores: Mexico DF, Mexico, 1999; p. 111. [Google Scholar]
- Wybraniec, S.; Stalica, P.; Jerz, G.; Klose, B.; Gebers, N.; Winterhalter, P.; Sporna, A.; Szaleniec, M.; Mizrahi, Y. Separation of polar betalain pigments from cacti fruits of Hylocereus costaricensis by ion-pair high-speed countercurrent chromatography. J. Chromatogr. A 2009, 1216, 6890–6899. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Mabry, T. The Betacyanins, a New Class of Red Violet Pigments, and Their Phylogenetic Significance; Roland Press: New York, NY, USA, 1964. [Google Scholar]
- Bate-Smith, E. The phenolic constituents of plants and their taxonomic significance. Bot. J. Linn. Soc. 1962, 58, 95–173. [Google Scholar] [CrossRef]
- Jain, G.; Gould, K.S. Are betalain pigments the functional homologues of anthocyanins in plants? Environ. Exp. Bot. 2015, 119, 48–53. [Google Scholar] [CrossRef]
- Gandia-Herrero, F.; Garcia-Carmona, F. Biosynthesis of betalains: Yellow and violet plant pigments. Trends Plant Sci. 2013, 18, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Polturak, G.; Grossman, N.; Vela-Corcia, D.; Dong, Y.; Nudel, A.; Pliner, M.; Levy, M.; Rogachev, I.; Aharoni, A. Engineered gray mold resistance, antioxidant capacity and pigmentation in betalain-producing crops and ornamentals. Proc. Natl. Acad. Sci. USA 2017, 114, 9062–9067. [Google Scholar] [CrossRef] [PubMed]
- Jain, G.; Gould, K.S. Functional significance of betalain biosynthesis in leaves of Disphyma australe under salinity stress. Environ. Exp. Bot. 2015, 109, 131–140. [Google Scholar] [CrossRef]
- Jain, G.; Schwinn, K.E.; Gould, K.S. Betalain induction by L-DOPA application confers photoprotection to saline-exposed leaves of Disphyma australe. New Phytol. 2015, 207, 1075–1083. [Google Scholar] [CrossRef]
- Casique-Arroyo, G.; Martinez-Gallardo, N.; de la Vara, L.G.; Delano-Frier, J.P. Betacyanin biosynthetic genes and enzymes are differentially induced by (a)biotic stress in Amaranthus hypochondriacus. PLoS ONE 2014, 9, e99012. [Google Scholar] [CrossRef]
- Hayakawa, K.; Agarie, S. Physiological roles of betacyanin in a halophyte, Suaeda japonica Makino. Plant Prod. Sci. 2010, 13, 351–359. [Google Scholar] [CrossRef]
- Nakashima, T.; Araki, T.; Ueno, O. Photoprotective function of betacyanin in leaves of Amaranthus cruentus L. under water stress. Photosynthetica 2011, 49, 497–506. [Google Scholar] [CrossRef]
- Sepulveda-Jimenez, G.; Rueda-Benitez, P.; Porta, H.; Rocha-Sosa, M. Betacyanin synthesis in red beet (Beta vulgaris) leaves induced by wounding and bacterial infiltration is preceded by an oxidative burst. PhysiolOgical and Molecular. Plant Pathol. 2004, 64, 125–133. [Google Scholar] [CrossRef]
- Ibdah, M.; Krins, A.; Seidlitz, H.K.; Heller, W.; Strack, D.; Vogt, T. Spectral dependence of flavonol and betacyanin accumulation in Mesembryanthemum crystallinum under enhanced ultraviolet radiation. Plant Cell Environ. 2002, 25, 1145–1154. [Google Scholar] [CrossRef]
- Vogt, T.; Grimm, R.; Strack, D. Cloning and expression of a cDNA encoding betanidin 5-O-glucosyltransferase, a betanidin- and flavonoid-specific enzyme with high homology to inducible glucosyltransferases from the Solanaceae. Plant J. 1999, 19, 509–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishima, Y.; Shimaya, A.; Adachi, T. Evidence that blue light induces betalain pigmentation in Portulaca callas. Plant Cell Tissue Organ Cult. 1995, 43, 67–70. [Google Scholar] [CrossRef]
- Nowacki, L.; Vigneron, P.; Rotellini, L.; Cazzola, H.; Merlier, F.; Prost, E.; Ralanairina, R.; Gadonna, J.P.; Rossi, C.; Vayssade, M. Betanin-Enriched Red Beetroot (Beta vulgaris L.) Extract Induces Apoptosis and Autophagic Cell Death in MCF-7 Cells. Phytother. Res. 2015, 29, 1964–1973. [Google Scholar] [CrossRef] [PubMed]
- Kanner, K.; Harel, S.; Granit, R. Betalains A new class of dietary cationized antioxidants. J. Agric. Food Chem. 2001, 49, 5178–5185. [Google Scholar] [CrossRef]
- Escribano, J.; Pedreoño, M.A.; Garcia-Carmona, F.; Munoz, R. Characterization of the antiradical activity of betalains from Beta vulgaris L. roots. Phytochem. Anal. 1998, 9, 124–127. [Google Scholar] [CrossRef]
- Polturak, G.; Breitel, D.; Grossman, N.; Sarrion-Perdigones, A.; Weithorn, E.; Pliner, M.; Orzaez, D.; Granell, A.; Rogachev, I.; Aharoni, A. Elucidation of the first committed step in betalain biosynthesis enables the heterologous engineering of betalain pigments in plants. New Phytol. 2016, 210, 269–283. [Google Scholar] [CrossRef]
- Sunnadeniya, R.; Bean, A.; Brown, M.; Akhavan, N.; Hatlestad, G.; Gonzalez, A.; Symonds, V.V.; Lloyd, A. Tyrosine Hydroxylation in Betalain Pigment Biosynthesis Is Performed by Cytochrome P450 Enzymes in Beets (Beta vulgaris). PLoS ONE 2016, 11, e0149417. [Google Scholar] [CrossRef] [PubMed]
- Gandia-Herrero, F.; Garcia-Carmona, F. Characterization of recombinant Beta vulgaris 4,5-DOPA-extradiol-dioxygenase active in the biosynthesis of betalains. Planta 2012, 236, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.L. Beyond brown: Polyphenol oxidases as enzymes of plant specialized metabolism. Front. Plant Sci. 2014, 5, 783. [Google Scholar] [CrossRef] [PubMed]
- Hatlestad, G.J.; Sunnadeniya, R.M.; Akhavan, N.A.; Gonzalez, A.; Goldman, I.L.; McGrath, J.M.; Lloyd, A.M. The beet R locus encodes a new cytochrome P450 required for red betalain production. Nat. Genet. 2012, 44, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T.; Zimmermann, E.; Grimm, R.; Meyer, M.; Strack, D. Are the characteristics of betanidin glucosyltransferases from cell-suspension cultures of Dorotheanthus bellidiformis indicative of their phylogenetic relationship with flavonoid glucosyltransferases? Planta 1997, 203, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T.; Ibdah, M.; Schmidt, J.; Wray, V.; Nimtz, M.; Strack, D. Light-induced betacyanin and flavonol accumulation in bladder cells of Mesembryanthemum crystallinum. Phytochemistry 1999, 52, 583–592. [Google Scholar] [CrossRef]
- Sasaki, N.; Wada, K.; Koda, T.; Kasahara, K.; Adachi, T.; Ozeki, Y. Isolation and characterization of cDNAs encoding an enzyme with glucosyltransferase activity for cyclo-DOPA from four o’clocks and feather cockscombs. Plant Cell Physiol. 2005, 46, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Schliemann, W.; Kobayashi, N.; Strack, D. The decisive step in betaxanthin biosynthesis is a spontaneous reaction. Plant Physiol. 1999, 119, 1217–1232. [Google Scholar] [CrossRef] [PubMed]
- Strack, D.; Vogt, T.; Schliemann, W. Recent advances in betalain research. Phytochemistry 2003, 62, 247–269. [Google Scholar] [CrossRef]
- Sasaki, N.; Adachi, T.; Koda, T.; Ozeki, Y. Detection of UDP-glucose:cyclo-DOPA 5-O-glucosyltransferase activity in four o’clocks (Mirabilis jalapa L.). FEBS Lett. 2004, 568, 159–162. [Google Scholar] [CrossRef]
- Hatlestad, G.J.; Akhavan, N.A.; Sunnadeniya, R.M.; Elam, L.; Cargile, S.; Hembd, A.; Gonzalez, A.; McGrath, J.M.; Lloyd, A.M. The beet Y locus encodes an anthocyanin MYB-like protein that activates the betalain red pigment pathway. Nat. Genet. 2015, 47, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Brockington, S.F.; Yang, Y.; Gandia-Herrero, F.; Covshoff, S.; Hibberd, J.M.; Sage, R.F.; Wong, G.K.; Moore, M.J.; Smith, S.A. Lineage-specific gene radiations underlie the evolution of novel betalain pigmentation in Caryophyllales. New Phytol. 2015, 207, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Bean, A.; Sunnadeniya, R.; Akhavan, N.; Campbell, A.; Brown, M.; Lloyd, A. Gain-of-function mutations in beet DODA2 identify key residues for betalain pigment evolution. New Phytol. 2018, 219, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Park, C.M. MYB96-mediated abscisic acid signals induce pathogen resistance response by promoting salicylic acid biosynthesis in Arabidopsis. New Phytol. 2010, 186, 471–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, P.J.; Xiang, F.; Qiao, M.; Park, J.-Y.; Lee, Y.N.; Kim, S.-G.; Lee, Y.-H.; Park, W.J.; Park, C.-M. The MYB96 transcription factor mediates abscisic acid signaling during drought stress response in Arabidopsis. Plant Physiol. 2009, 151, 275–289. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef]
- Cone, K.C.; Burr, F.A.; Burr, B. Molecular analysis of the maize anthocyanin regulatory locus Cl. Proc. Natl. Acad. Sci. USA 1986, 83, 9631–9635. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2013, 2013 29, 644–652. [Google Scholar]
- Romualdi, C.; Bortoluzzi, S.; d’Alessi, F.; Danieli, G.A. IDEG6: A web tool for detection of differentially expressed genes in multiple tag sampling experiments. Physiol. Genomics 2013, 12, 159–162. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinf. 2003, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaeffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Iseli, C.; Jongeneel, C.V.; Bucher, P. ESTScan: A program for detecting, evaluating, and reconstructing potential coding regions in EST sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1999, 23, 138–148. [Google Scholar]
- Gene Ontology Consortium. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [PubMed]
- Rivals, I.; Personnaz, L.; Taing, L.; Potier, M.-C. Enrichment or depletion of a GO category within a class of genes: Which test? Bioinformatics 2007, 23, 401–407. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Values | Total | Nr a | Nt b | Swissprot c | KEGG d | KOG e | GO f | Overall |
|---|---|---|---|---|---|---|---|---|
| Number | 79,049 | 51,093 | 39,651 | 36,059 | 38,851 | 39,492 | 14,378 | 52,618 |
| Percentage | 100% | 64.63% | 50.16% | 45.62% | 49.15% | 49.96% | 18.19% | 66.56% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, X.; Zong, Y.; Li, S.; Cao, D.; Sun, X.; Liu, B. Transcriptome Analysis Clarified Genes Involved in Betalain Biosynthesis in the Fruit of Red Pitayas (Hylocereus costaricensis). Molecules 2019, 24, 445. https://doi.org/10.3390/molecules24030445
Xi X, Zong Y, Li S, Cao D, Sun X, Liu B. Transcriptome Analysis Clarified Genes Involved in Betalain Biosynthesis in the Fruit of Red Pitayas (Hylocereus costaricensis). Molecules. 2019; 24(3):445. https://doi.org/10.3390/molecules24030445
Chicago/Turabian StyleXi, Xingyuan, Yuan Zong, Shiming Li, Dong Cao, Xuemei Sun, and Baolong Liu. 2019. "Transcriptome Analysis Clarified Genes Involved in Betalain Biosynthesis in the Fruit of Red Pitayas (Hylocereus costaricensis)" Molecules 24, no. 3: 445. https://doi.org/10.3390/molecules24030445
APA StyleXi, X., Zong, Y., Li, S., Cao, D., Sun, X., & Liu, B. (2019). Transcriptome Analysis Clarified Genes Involved in Betalain Biosynthesis in the Fruit of Red Pitayas (Hylocereus costaricensis). Molecules, 24(3), 445. https://doi.org/10.3390/molecules24030445