
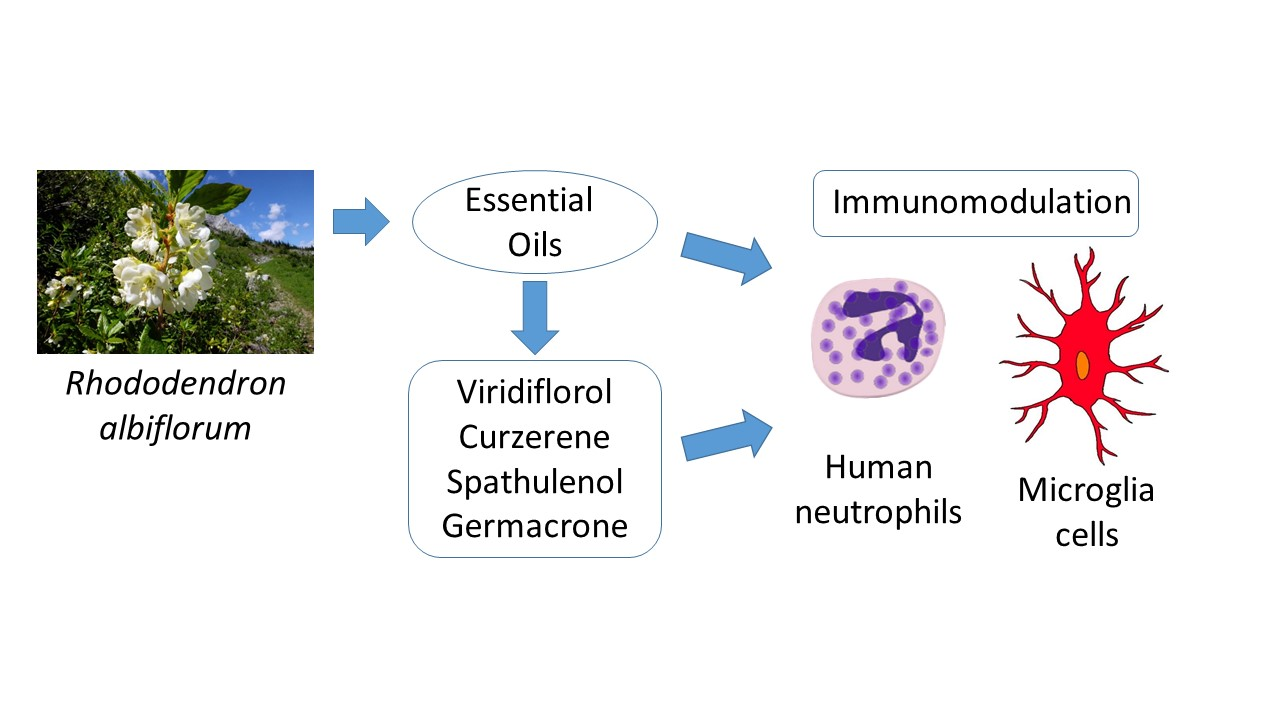
Chemical Composition and Immunomodulatory Activity of Essential Oils from Rhododendron albiflorum

 ,
, 
 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Materials
2.3. Essential Oil Isolation
2.4. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.5. Isolation of Human Neutrophils
2.6. Cell Culture
2.7. Ca2+ Mobilization Assay
2.8. Chemotaxis Assay
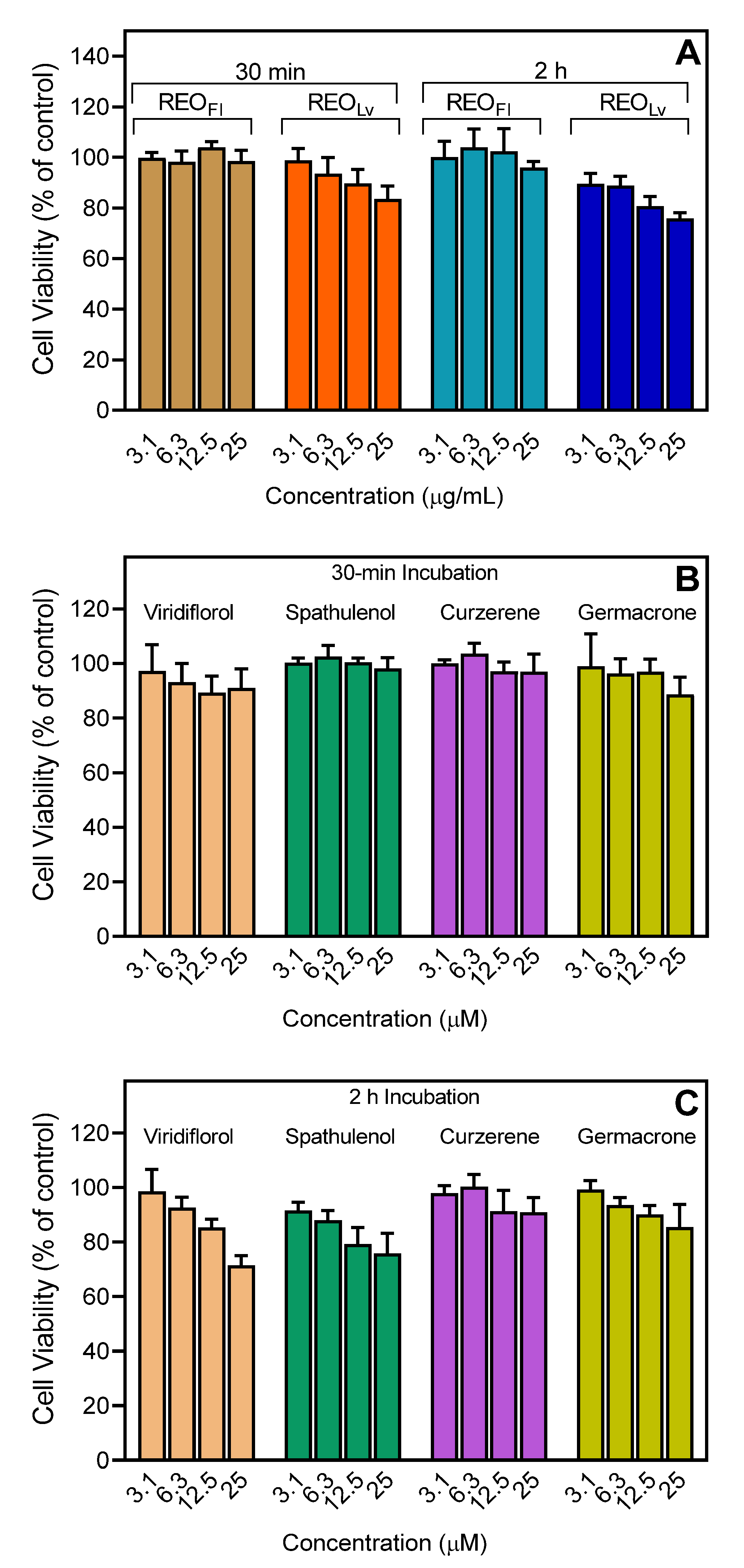
2.9. Cytotoxicity Assay
2.10. Kinase Kd Determination
2.11. Molecular Modeling
2.12. Statistical Analysis
3. Results and Discussion
3.1. Essential Oil Composition
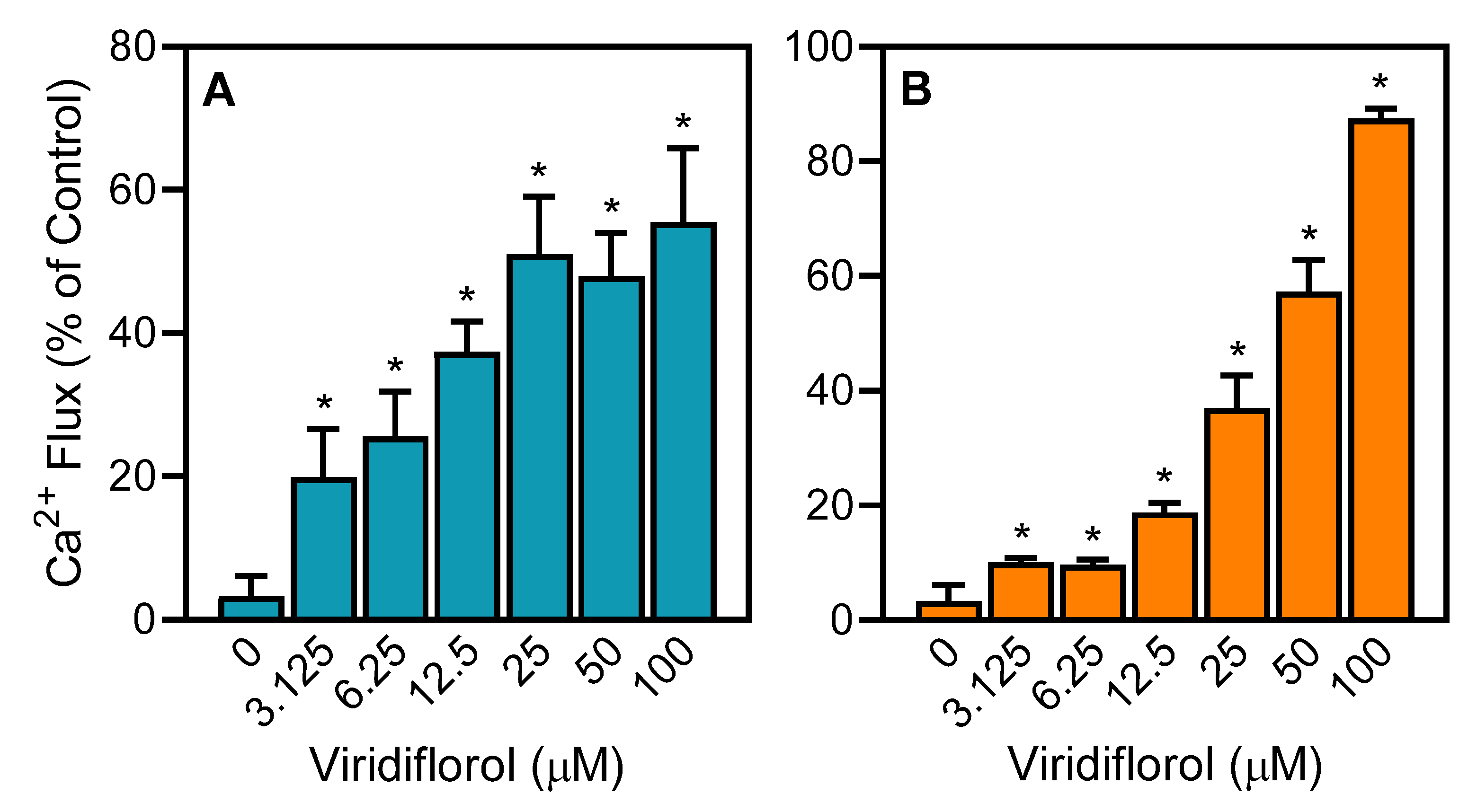
3.2. Effect of the R. albiflorum Essential Oils and Component Compounds on Neutrophil and Microglial [Ca2+]i
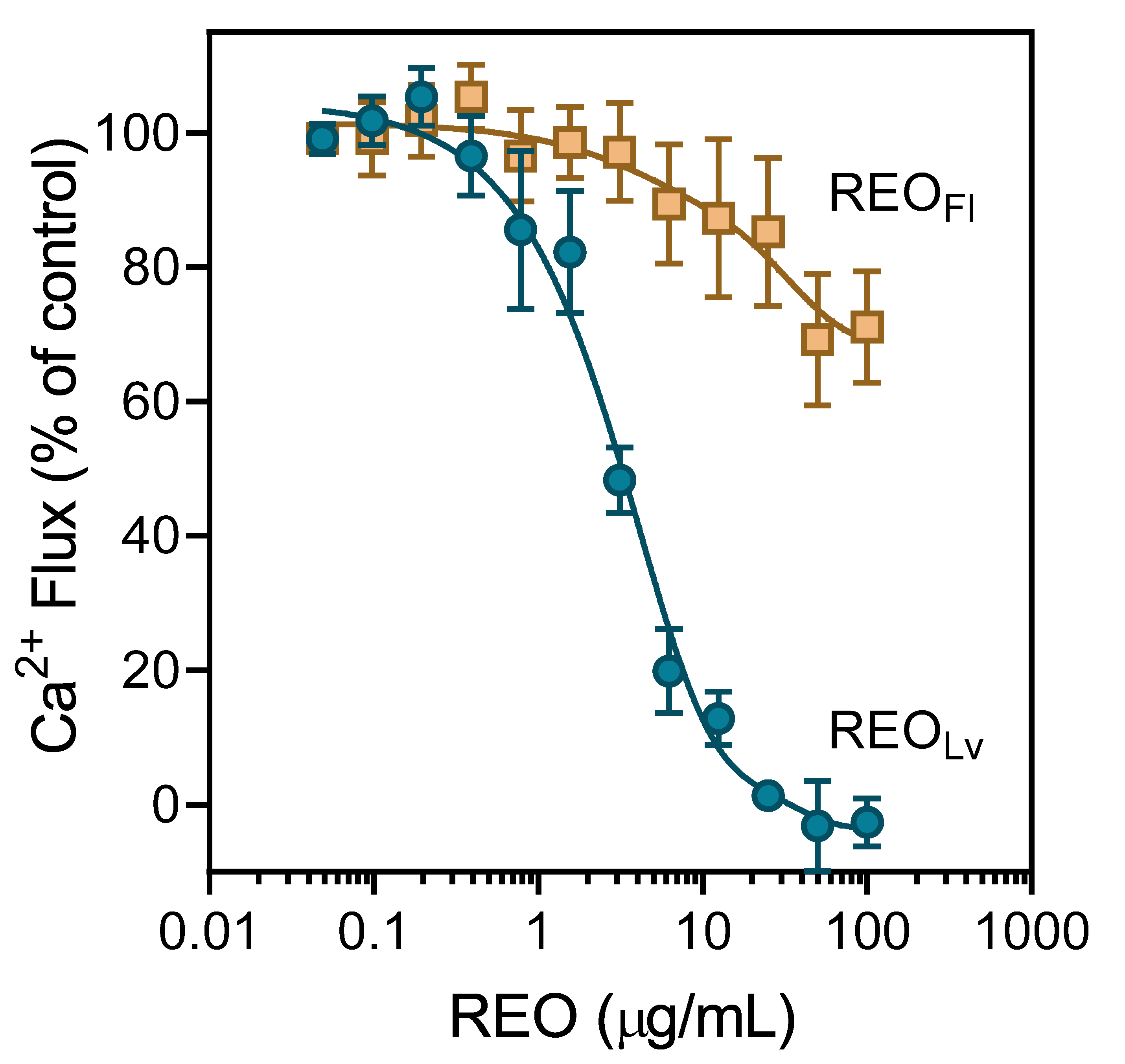
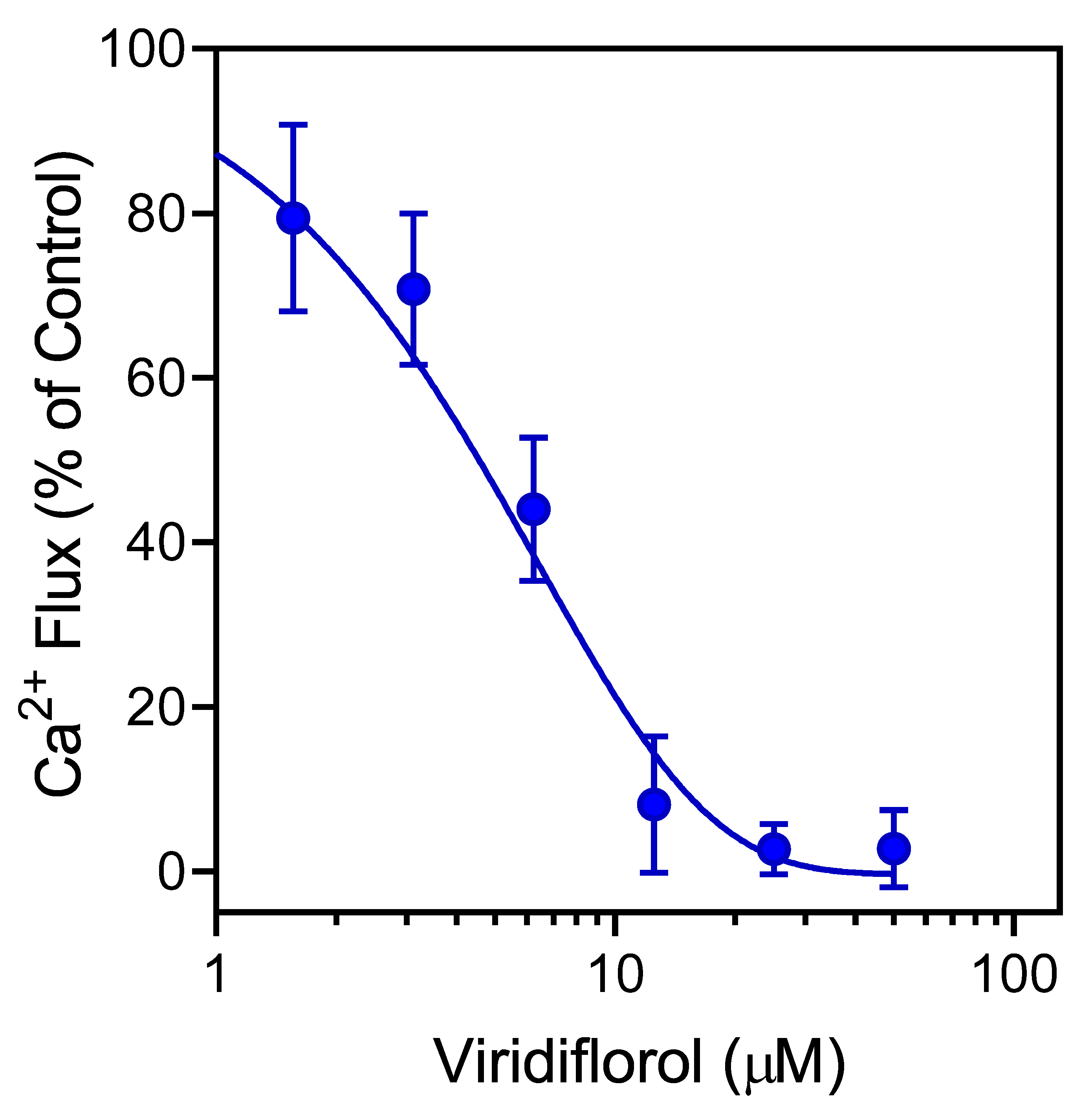
3.3. Effect of R. albiflorum Essential Oils and Component Compounds on Agonist-Induced Ca2+ Influx
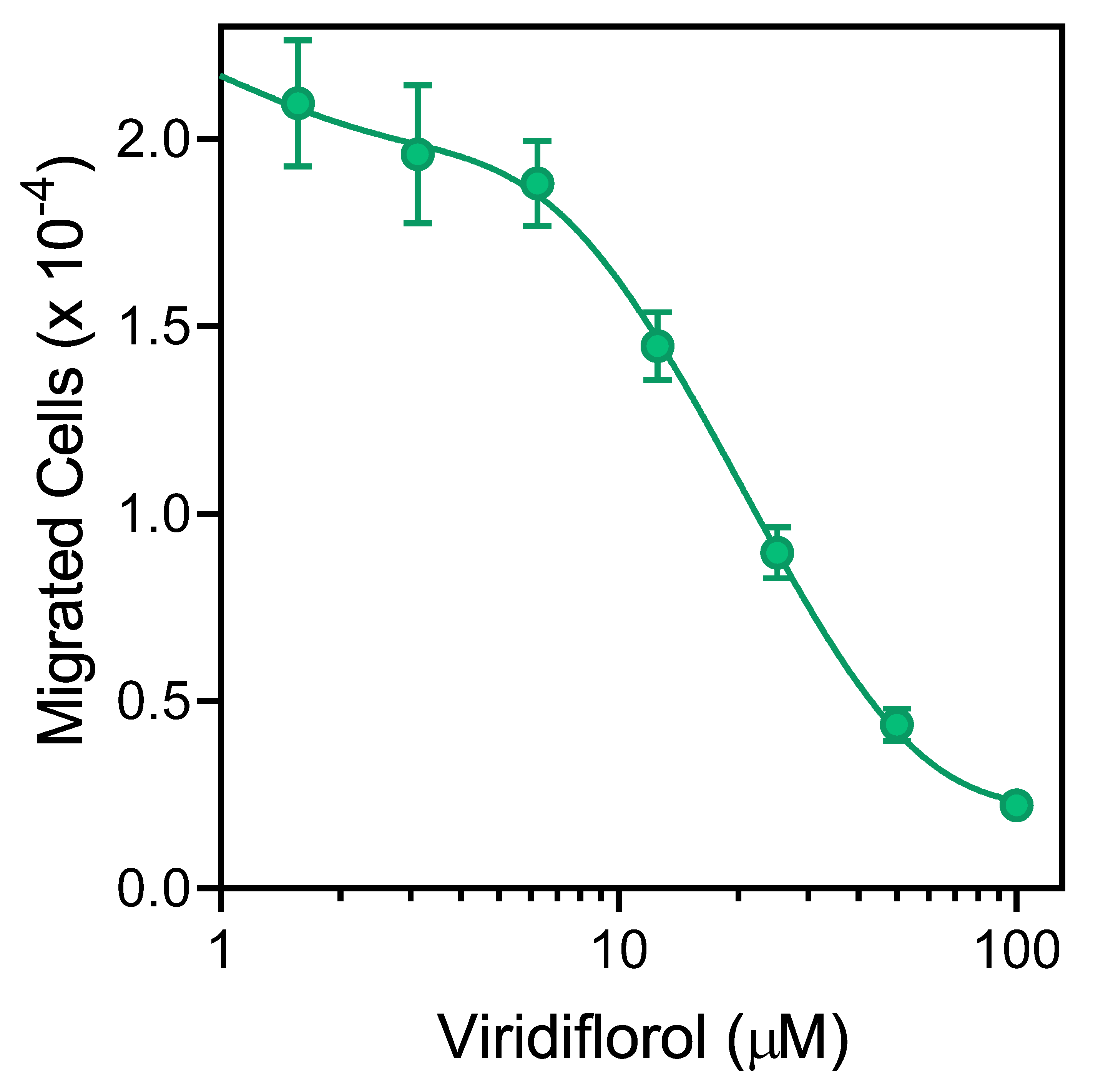
3.4. Effect of R. albiflorum Essential Oils and Component Compounds on Neutrophil Chemotaxis
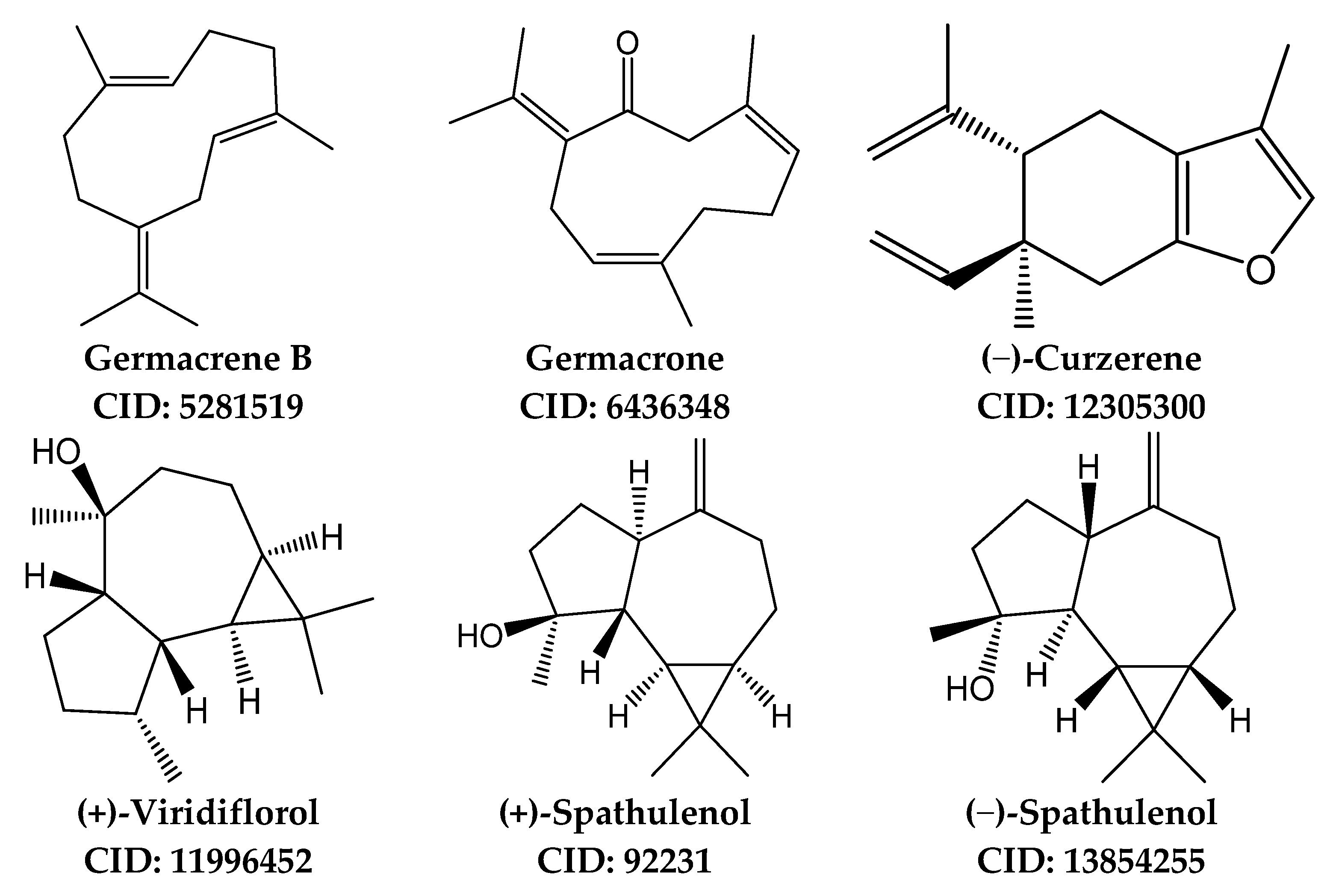
3.5. Identification of Potential Protein Targets for Selected Sesquiterpenes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Popescu, R.; Kopp, B. The genus Rhododendron: An ethnopharmacological and toxicological review. J. Ethnopharmacol. 2013, 147, 42–62. [Google Scholar] [CrossRef]
- Jaiswal, R.; Jayasinghe, L.; Kuhnert, N. Identification and characterization of proanthocyanidins of 16 members of the Rhododendron genus (Ericaceae) by tandem LC-MS. J. Mass Spectrom. 2012, 47, 502–515. [Google Scholar] [CrossRef]
- Prakash, D.; Upadhyay, G.; Singh, B.N.; Dhakarey, R.; Kumar, S.; Singh, K.K. Free-radical scavenging activities of himalayan rhododendrons. Curr. Sci. India 2007, 92, 526–532. [Google Scholar]
- Gautam, V.; Sharma, A.; Arora, S.; Bhardwaj, R.; Ahmad, A.; Ahamad, B.; Ahmad, P. In-vitro antioxidant, antimutagenic and cancer cell growth inhibition activities of Rhododendron arboreum leaves and flowers. Saudi J. Biol. Sci. 2020, 27, 1788–1796. [Google Scholar] [CrossRef]
- Zong, L.; Zhang, J.; Dai, L.; Liu, J.; Yang, Y.; Xie, J.; Luo, X. The anti-inflammatory properties of Rhododendron molle leaf extract in LPS-induced RAW264.7. Chem. Biodivers. 2020, 17, e2000477. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Dai, L.F.; Zhang, H.Y.; Chen, M.N.; Cai, X.F.; Xie, J.K.; Luo, X.D. Unlocking the potential antioxidant and anti-inflammatory activities of Rhododendron molle G. Don. Pak. J. Pharm. Sci. 2019, 32, 2375–2383. [Google Scholar]
- Guo, Y.; Yu, X.M.; Chen, S.; Wen, J.Y.; Chen, Z.W. Total flavones of Rhododendron simsii planch flower protect rat hippocampal neuron from hypoxia-reoxygenation injury via activation of BKCA channel. J. Pharm. Pharmacol. 2020, 72, 111–120. [Google Scholar] [CrossRef]
- Gautam, V.; Kohli, S.K.; Arora, S.; Bhardwaj, R.; Kazi, M.; Ahmad, A.; Raish, M.; Ganaie, M.A.; Ahmad, P. Antioxidant and antimutagenic activities of different fractions from the leaves of Rhododendron arboreum Sm. and their GC-MS profiling. Molecules 2018, 23, 2239. [Google Scholar] [CrossRef] [Green Version]
- Rapinski, M.; Musallam, L.; Arnason, J.T.; Haddad, P.; Cuerrier, A. Adipogenic activity of wild populations of Rhododendron groenlandicum, a medicinal shrub from the James Bay Cree traditional pharmacopeia. Evid. Based Complement. Altern. Med. 2015, 2015, 492458. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.M.; Hsu, Y.M.; Jhan, Y.L.; Tsai, S.J.; Lin, S.X.; Su, C.H.; Chou, C.H. Structure elucidation of procyanidins isolated from Rhododendron formosanum and their anti-oxidative and anti-bacterial activities. Molecules 2015, 20, 12787–12803. [Google Scholar] [CrossRef] [Green Version]
- Moreman, D.E. Native American Medicinal Plants. An Ethnobotanical Dictionary; Timber Press: Portland, OR, USA, 2009; p. 799. [Google Scholar]
- Zuo, X.; Gu, Y.N.; Wang, C.; Zhang, J.R.; Zhang, J.; Wang, G.Q.; Wang, F. A systematic review of the anti-inflammatory and immunomodulatory properties of 16 essential oils of herbs. Evid. Based Compl. Alt. 2020, 2020, 8878927. [Google Scholar] [CrossRef]
- Reis, D.; Jones, T. Aromatherapy using essential oils as a supportive therapy. Clin. J. Oncol. Nurs. 2017, 21, 16–19. [Google Scholar] [CrossRef]
- Rombola, L.; Amantea, D.; Russo, R.; Adornetto, A.; Berliocchi, L.; Tridico, L.; Corasaniti, M.T.; Sakurada, S.; Sakurada, T.; Bagetta, G.; et al. Rational basis for the use of bergamot essential oil in complementary medicine to treat chronic pain. Mini Rev. Med. Chem. 2016, 16, 721–728. [Google Scholar] [CrossRef]
- Di Martile, M.; Garzoli, S.; Ragno, R.; Del Bufalo, D. Essential oils and their main chemical components: The past 20 years of preclinical studies in melanoma. Cancers 2020, 12, 2650. [Google Scholar] [CrossRef]
- de Cassia da Silveira, E.S.R.; Andrade, L.N.; Dos Reis Barreto de Oliveira, R.; de Sousa, D.P. A review on anti-inflammatory activity of phenylpropanoids found in essential oils. Molecules 2014, 19, 1459–1480. [Google Scholar] [CrossRef] [Green Version]
- Judzentiene, A.; Budiene, J.; Svediene, J.; Garjonyte, R. Toxic, radical scavenging, and antifungal activity of Rhododendron tomentosum H. Essential oils. Molecules 2020, 25, 1676. [Google Scholar] [CrossRef] [Green Version]
- Jesionek, A.; Kokotkiewicz, A.; Mikosik-Roczynska, A.; Ciesielska-Figlon, K.; Luczkiewicz, P.; Bucinski, A.; Daca, A.; Witkowski, J.M.; Bryl, E.; Zabiegala, B.; et al. Chemical variability of Rhododendron tomentosum (Ledum Palustre) essential oils and their pro-apoptotic effect on lymphocytes and rheumatoid arthritis synoviocytes. Fitoterapia 2019, 139, 104402. [Google Scholar] [CrossRef]
- Raal, A.; Orav, A.; Gretchushnikova, T. Composition of the essential oil of the Rhododendron tomentosum Harmaja from Estonia. Nat. Prod. Res. 2014, 28, 1091–1098. [Google Scholar] [CrossRef]
- Yang, K.; Zhou, Y.X.; Wang, C.F.; Du, S.S.; Deng, Z.W.; Liu, Q.Z.; Liu, Z.L. Toxicity of Rhododendron anthopogonoides essential oil and its constituent compounds towards Sitophilus zeamais. Molecules 2011, 16, 7320–7330. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Jiao, M.L.; Zang, H.Y.; Guo, S.S.; Wang, Y.; Sang, Y.L.; Du, S.S. Chemical composition of essential oils from four rhododendron species and their repellent activity against three stored-product insects. Environ. Sci. Pollut. Res. 2019, 26, 23198–23205. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Satyal, P.; Pokharel, S.; Setzer, W.N. Chemical composition, enantiomeric distribution, and biological activities of rhododendron anthopogon leaf essential oil from nepal. Nat. Prod. Commun. 2016, 11, 1895–1898. [Google Scholar] [CrossRef] [Green Version]
- Innocenti, G.; Dall’Acqua, S.; Scialino, G.; Banfi, E.; Sosa, S.; Gurung, K.; Barbera, M.; Carrara, M. Chemical composition and biological properties of Rhododendron anthopogon essential oil. Molecules 2010, 15, 2326–2338. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B. Innate immunity: An overview. Mol. Immunol. 2004, 40, 845–859. [Google Scholar] [CrossRef] [PubMed]
- Witko-Sarsat, V.; Rieu, P.; Descamps-Latscha, B.; Lesavre, P.; Halbwachs-Mecarelli, L. Neutrophils: Molecules, functions and pathophysiological aspects. Lab. Investig. 2000, 80, 617–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, S.; Steffy, K.; Averett, D. Masked oral prodrugs of Toll-like receptor 7 agonists: A new approach for the treatment of infectious disease. Curr. Opin. Investig. Drugs 2006, 7, 702–708. [Google Scholar] [PubMed]
- Schepetkin, I.A.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. High-throughput screening for small-molecule activators of neutrophils: Identification of novel n-formyl peptide receptor agonists. Mol. Pharmacol. 2007, 71, 1061–1074. [Google Scholar] [CrossRef] [Green Version]
- Reshetnikov, V.; Hahn, J.; Maueroder, C.; Czegley, C.; Munoz, L.E.; Herrmann, M.; Hoffmann, M.H.; Mokhir, A. Chemical tools for targeted amplification of reactive oxygen species in neutrophils. Front. Immunol. 2018, 9, 1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepetkin, I.A.; Kushnarenko, S.V.; Ozek, G.; Kirpotina, L.N.; Sinharoy, P.; Utegenova, G.A.; Abidkulova, K.T.; Ozek, T.; Baser, K.H.; Kovrizhina, A.R.; et al. Modulation of human neutrophil responses by the essential oils from Ferula akitschkensis and their constituents. J. Agric. Food Chem. 2016, 64, 7156–7170. [Google Scholar] [CrossRef] [Green Version]
- Ozek, G.; Schepetkin, I.A.; Utegenova, G.A.; Kirpotina, L.N.; Andrei, S.R.; Ozek, T.; Baser, K.H.C.; Abidkulova, K.T.; Kushnarenko, S.V.; Khlebnikov, A.I.; et al. Chemical composition and phagocyte immunomodulatory activity of Ferula iliensis essential oils. J. Leukoc. Biol. 2017, 101, 1361–1371. [Google Scholar] [CrossRef] [Green Version]
- Schepetkin, I.A.; Kushnarenko, S.V.; Ozek, G.; Kirpotina, L.N.; Utegenova, G.A.; Kotukhov, Y.A.; Danilova, A.N.; Ozek, T.; Baser, K.H.; Quinn, M.T. Inhibition of human neutrophil responses by the essential oil of Artemisia kotuchovii and its constituents. J. Agric. Food. Chem. 2015, 63, 4999–5007. [Google Scholar] [CrossRef] [Green Version]
- Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical composition and immunomodulatory activity of Hypericum perforatum essential oils. Biomolecules 2020, 10, 916. [Google Scholar] [CrossRef]
- Cunningham, C. Microglia and neurodegeneration: The role of systemic inflammation. Glia 2013, 61, 71–90. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Li, G.; Yu, J.; Xu, K.; Wu, W. Receptors, channel proteins, and enzymes involved in microglia-mediated neuroinflammation and treatments by targeting microglia in ischemic stroke. Neuroscience 2021, 460, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Yang, J.M.; Wang, X.Y.; Yang, P.; Zhao, Y.; Li, K.; Chen, Y.J.; Xu, Y. Herbal compounds play a role in neuroprotection through the inhibition of microglial activation. J. Immunol. Res. 2018, 2018, 9348046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Darwish, M.S.; Cabral, C.; Goncalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Efferth, T.; Salgueiro, L. Artemisia herba-alba essential oil from Buseirah (South Jordan): Chemical characterization and assessment of safe antifungal and anti-inflammatory doses. J. Ethnopharmacol. 2015, 174, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhang, X.; Ren, F.; Yan, T.; Wu, B.; Bi, K.; Bi, W.; Jia, Y. Essential oil of Schisandra chinensis ameliorates cognitive decline in mice by alleviating inflammation. Food Funct. 2019, 10, 5827–5842. [Google Scholar] [CrossRef]
- Li, Y.; Lv, O.; Zhou, F.; Li, Q.; Wu, Z.; Zheng, Y. Linalool inhibits LPS-induced inflammation in BV2 microglia cells by activating Nrf2. Neurochem. Res. 2015, 40, 1520–1525. [Google Scholar] [CrossRef]
- Ozek, G.; Ishmuratova, M.; Tabanca, N.; Radwan, M.M.; Goger, F.; Ozek, T.; Wedge, D.E.; Becnel, J.J.; Cutler, S.J.; Can Baser, K.H. One-step multiple component isolation from the oil of Crinitaria tatarica (Less.) Sojak by preparative capillary gas chromatography with characterization by spectroscopic and spectrometric techniques and evaluation of biological activity. J. Sep. Sci. 2012, 35, 650–660. [Google Scholar] [CrossRef]
- Garcia-Mesa, Y.; Jay, T.R.; Checkley, M.A.; Luttge, B.; Dobrowolski, C.; Valadkhan, S.; Landreth, G.E.; Karn, J.; Alvarez-Carbonell, D. Immortalization of primary microglia: A new platform to study hiv regulation in the central nervous system. J. Neurovirol. 2017, 23, 47–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, M.W.; Herrgard, S.; Treiber, D.K.; Gallant, P.; Atteridge, C.E.; Campbell, B.T.; Chan, K.W.; Ciceri, P.; Davis, M.I.; Edeen, P.T.; et al. A quantitative analysis of kinase inhibitor selectivity. Nat. Biotechnol. 2008, 26, 127–132. [Google Scholar] [CrossRef]
- Liu, X.; Ouyang, S.; Yu, B.; Liu, Y.; Huang, K.; Gong, J.; Zheng, S.; Li, Z.; Li, H.; Jiang, H. Pharmmapper server: A web server for potential drug target identification using pharmacophore mapping approach. Nucleic. Acids Res. 2010, 38, W609–W614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Schaff, U.Y.; Yamayoshi, I.; Tse, T.; Griffin, D.; Kibathi, L.; Simon, S.I. Calcium flux in neutrophils synchronizes β2 integrin adhesive and signaling events that guide inflammatory recruitment. Ann. Biomed. Eng. 2008, 36, 632–646. [Google Scholar] [CrossRef] [Green Version]
- Krause, K.H.; Campbell, K.P.; Welsh, M.J.; Lew, D.P. The calcium signal and neutrophil activation. Clin. Biochem. 1990, 23, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Richardon, R.M.; Ali, H.; Tomhave, E.D.; Haribabu, B.; Snyderman, R. Cross-desensitization of chemoattractant receptors occurs at multiple levels—Evidence for a role for inhibition of phospholipase-c activity. J. Biol. Chem. 1995, 270, 27829–27833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fachini-Queiroz, F.C.; Kummer, R.; Estevao-Silva, C.F.; Carvalho, M.D.; Cunha, J.M.; Grespan, R.; Bersani-Amado, C.A.; Cuman, R.K. Effects of thymol and carvacrol, constituents of Thymus vulgaris L. essential oil, on the inflammatory response. Evid. Based Complement. Altern. Med. 2012, 2012, 657026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielli, L.J.; de Souza, T.J.T.; Maciel, A.J.; Ferrao, M.F.; Fuentefria, A.M.; Apel, M.A. Influence of monoterpenes in biological activities of Nectandra megapotamica (Spreng.) Mez essential oils. Biomolecules 2019, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Trevizan, L.N.F.; do Nascimento, K.F.; Santos, J.A.; Kassuya, C.A.L.; Cardoso, C.A.L.; Vieira, M.D.; Moreira, F.M.F.; Croda, J.; Formagio, A.S.N. Anti-inflammatory, antioxidant and anti-mycobacterium tuberculosis activity of viridiflorol: The major constituent of Allophylus edulis (A. St.-Hil., A. Juss. & Cambess.) Radlk. J. Ethnopharmacol. 2016, 192, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Gilabert, M.; Marcinkevicius, K.; Andujar, S.; Schiavone, M.; Arena, M.E.; Bardon, A. Sesqui and triterpenoids from the liverwort Lepidozia chordulifera inhibitors of bacterial biofilm and elastase activity of human pathogenic bacteria. Phytomedicine 2015, 22, 77–85. [Google Scholar] [CrossRef]
- Wang, Y.D.; Li, J.H.; Guo, J.Q.; Wang, Q.Y.; Zhu, S.G.; Gao, S.Y.; Yang, C.; Wei, M.; Pan, X.D.; Zhu, W.; et al. Cytotoxic and antitumor effects of curzerene from Curcuma longa. Planta Med. 2017, 83, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, P.L.B.; Pinto, L.C.; da Costa, J.S.; da Silva, A.R.C.; Mourao, R.H.V.; Montenegro, R.C.; da Silva, J.K.R.; Maia, J.G.S. Composition, antioxidant capacity and cytotoxic activity of Eugenia uniflora L. chemotype-oils from the amazon. J. Ethnopharmacol. 2019, 232, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, E.; Radai, J.A.S.; do Nascimento, K.F.; Formagio, A.S.N.; de Matos Balsalobre, N.; Ziff, E.B.; Castelon Konkiewitz, E.; Kassuya, C.A.L. Contribution of spathulenol to the anti-nociceptive effects of Psidium guineense. Nutr. Neurosci. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, K.F.; Moreira, F.M.F.; Alencar Santos, J.; Kassuya, C.A.L.; Croda, J.H.R.; Cardoso, C.A.L.; Vieira, M.D.C.; Gois Ruiz, A.L.T.; Ann Foglio, M.; de Carvalho, J.E.; et al. Antioxidant, anti-inflammatory, antiproliferative and antimycobacterial activities of the essential oil of Psidium guineense Sw. and spathulenol. J. Ethnopharmacol. 2018, 210, 351–358. [Google Scholar] [CrossRef]
- Bomfim, L.M.; Menezes, L.R.; Rodrigues, A.C.; Dias, R.B.; Rocha, C.A.; Soares, M.B.; Neto, A.F.; Nascimento, M.P.; Campos, A.F.; Silva, L.C.; et al. Antitumour activity of the microencapsulation of Annona vepretorum essential oil. Basic Clin. Pharmacol. Toxicol. 2016, 118, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Perez-Hernandez, N.; Ponce-Monter, H.; Medina, J.A.; Joseph-Nathan, P. Spasmolytic effect of constituents from Lepechinia caulescens on rat uterus. J. Ethnopharmacol. 2008, 115, 30–35. [Google Scholar] [CrossRef]
- Riaz, A.; Rasul, A.; Kanwal, N.; Hussain, G.; Shah, M.A.; Sarfraz, I.; Ishfaq, R.; Batool, R.; Rukhsar, F.; Adem, S. Germacrone: A potent secondary metabolite with therapeutic potential in metabolic diseases, cancer and viral infections. Curr. Drug Metab. 2020, 21, 1079–1090. [Google Scholar] [CrossRef]
- Liao, Q.; Qian, Z.; Liu, R.; An, L.; Chen, X. Germacrone inhibits early stages of influenza virus infection. Antivir. Res. 2013, 100, 578–588. [Google Scholar] [CrossRef]
- He, W.; Zhai, X.; Su, J.; Ye, R.; Zheng, Y.; Su, S. Antiviral activity of germacrone against pseudorabies virus in vitro. Pathogens 2019, 8, 258. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Bai, X.; Cui, T.; Zhou, H.; Chen, Y.; Xie, J.; Shi, Q.; Wang, H.; Zhang, G. In vitro antiviral activity of germacrone against porcine reproductive and respiratory syndrome virus. Curr. Microbiol. 2016, 73, 317–323. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W.; Fang, B.; Ma, F.; Zheng, Q.; Deng, P.; Zhao, S.; Chen, M.; Yang, G.; He, G. Anti-tumor effect of germacrone on human hepatoma cell lines through inducing G2/M cell cycle arrest and promoting apoptosis. Eur. J. Pharmacol. 2013, 698, 95–102. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Zheng, Q.; Fang, B.; Wang, W.; Ma, F.Y.; Roshan, S.; Banafa, A.; Chen, M.J.; Chang, J.L.; Deng, X.M.; et al. Germacrone induces apoptosis in human hepatoma HEPG2 cells through inhibition of the JAK2/STAT3 signalling pathway. J. Huazhong. Univ. Sci. Technolog. Med. Sci. 2013, 33, 339–345. [Google Scholar] [CrossRef] [PubMed]
- An, J.F.; Sun, Y.; Zhang, Q.L.; Zhang, F.L.; Zhang, J.L. The effects of germacrone on lipopolysaccharide-induced acute lung injury in neonatal rats. Cell Mol. Biol. 2014, 60, 8–12. [Google Scholar] [PubMed]
- Zhong, Z.; Chen, X.; Tan, W.; Xu, Z.; Zhou, K.; Wu, T.; Cui, L.; Wang, Y. Germacrone inhibits the proliferation of breast cancer cell lines by inducing cell cycle arrest and promoting apoptosis. Eur. J. Pharmacol. 2011, 667, 50–55. [Google Scholar] [CrossRef]
- Lim, M.S.; Choung, S.Y.; Jeong, K.W. Germacrone inhibits estrogen receptor α-mediated transcription in MCF-7 breast cancer cells. Phytother. Res. 2016, 30, 2036–2043. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Xu, J.; Shao, M.; Zou, J. Germacrone induces apoptosis as well as protective autophagy in human prostate cancer cells. Cancer Manag. Res. 2020, 12, 4009–4016. [Google Scholar] [CrossRef]
- Wang, Z.; Zhuo, F.; Chu, P.; Yang, X.; Zhao, G. Germacrone alleviates collagen-induced arthritis via regulating Th1/Th2 balance and NF-κB activation. Biochem. Biophys. Res. Commun. 2019, 518, 560–564. [Google Scholar] [CrossRef]
- Wu, T.; Yin, F.; Kong, H.; Peng, J. Germacrone attenuates cerebral ischemia/reperfusion injury in rats via antioxidative and antiapoptotic mechanisms. J. Cell Biochem. 2019, 120, 18901–18909. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, W.; Jin, L.; Zhang, G.; Yang, H.; Yu, B. Inhibitory activities of curzerenone, curdione, furanodienone, curcumol and germacrone on Ca2+-activated chloride channels. Fitoterapia 2020, 147, 104736. [Google Scholar] [CrossRef]
- Dolesjs, L.; Motl, O.; Soucek, M.; Herout, V.; Sorm, F. On terpenes. CVIII. Epimeric aromadendrenes. Stereoisomerism of ledol, viridiflorol and globulol. Collect. Czech. Chem. Commun. 1960, 25, 1483–1491. [Google Scholar] [CrossRef]
- Buckingham, J. (Ed.) Dictionary of Natural Products; CRC Press: Boca Raton, FL, USA, 1997; Volume 11, p. 615. [Google Scholar]
- Xiang, C.P.; Han, J.X.; Li, X.C.; Li, Y.H.; Zhang, Y.; Chen, L.; Qu, Y.; Hao, C.Y.; Li, H.Z.; Yang, C.R.; et al. Chemical composition and acetylcholinesterase inhibitory activity of essential oils from piper species. J. Agric. Food Chem. 2017, 65, 3702–3710. [Google Scholar] [CrossRef]
- Ghavam, M.; Manca, M.L.; Manconi, M.; Bacchetta, G. Chemical composition and antimicrobial activity of essential oils obtained from leaves and flowers of Salvia hydrangea Dc. Ex Benth. Sci. Rep. 2020, 10, 15647. [Google Scholar] [CrossRef]
- Benovit, S.C.; Silva, L.L.; Salbego, J.; Loro, V.L.; Mallmann, C.A.; Baldisserotto, B.; Flores, E.M.; Heinzmann, B.M. Anesthetic activity and bio-guided fractionation of the essential oil of Aloysia gratissima (Gillies & Hook.) Tronc. in silver catfish Rhamdia quelen. An. Acad. Bras. Cienc. 2015, 87, 1675–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beattie, K.D.; Waterman, P.G.; Forster, P.I.; Thompson, D.R.; Leach, D.N. Chemical composition and cytotoxicity of oils and eremophilanes derived from various parts of Eremophila mitchellii Benth. (Myoporaceae). Phytochemistry 2011, 72, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Luciardi, M.C.; Perez Hernandez, M.V.; Muruaga, N.; Bardon, A.; Arena, M.E.; Cartagena, E. Volatiles from subtropical Convolvulaceae that interfere with bacterial cell-to-cell communication as potential antipathogenic drugs. Evid. Based Complement. Altern. Med. 2016, 2016, 7890260. [Google Scholar] [CrossRef] [Green Version]
- Hulley, I.M.; van Vuuren, S.F.; Sadgrove, N.J.; van Wyk, B.E. Antimicrobial activity of Elytropappus rhinocerotis (Asteraceae) against micro-organisms associated with foot odour and skin ailments. J. Ethnopharmacol. 2019, 228, 92–98. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Peng, C.X.; Hu, Y.; Bu, C.; Guo, S.C.; Li, X.; Chen, Y.; Chen, J.W. Studies on chemical constituents and anti-hepatoma effects of essential oil from Annona squamosa L. Pericarps. Nat. Prod. Res. 2017, 31, 1305–1308. [Google Scholar] [CrossRef]
- Xiao, Z.; Fan, B.; Niu, Y.; Wu, M.; Liu, J.; Ma, S. Characterization of odor-active compounds of various chrysanthemum essential oils by gas chromatography-olfactometry, gas chromatography-mass spectrometry and their correlation with sensory attributes. J. Chromatogr. B 2016, 1009–1010, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, B.; Yang, F.; Sun, Q.; Yang, Z.; Zhu, L. Chemical compositionand anti-acetyl cholinesterase activity of flower essential oils of Artemisia annua at different flowering stage. Iran. J. Pharm. Res. 2011, 10, 265–271. [Google Scholar] [CrossRef] [Green Version]
- McNeil, M.J.; Porter, R.B.; Williams, L.A.; Rainford, L. Chemical composition and antimicrobial activity of the essential oils from Cleome spinosa. Nat. Prod. Commun. 2010, 5, 1301–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixit, N.; Kim, M.H.; Rossaint, J.; Yamayoshi, I.; Zarbock, A.; Simon, S.I. Leukocyte function antigen-1, kindlin-3, and calcium flux orchestrate neutrophil recruitment during inflammation. J. Immunol. 2012, 189, 5954–5964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, F.; Deng, J.; Wang, G.; Ye, R.D.; Christman, J.W. Pivotal role of mitogen-activated protein kinase-activated protein kinase 2 in inflammatory pulmonary diseases. Curr. Protein Pept. Sci. 2016, 17, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.F.; Chen, C.Y.; Lin, I.W.; Leu, Y.L.; Yang, S.C.; Syu, Y.T.; Chen, P.J.; Hwang, T.L. Imperatorin alleviates psoriasiform dermatitis by blocking neutrophil respiratory burst, adhesion, and chemotaxis through selective phosphodiesterase 4 inhibition. Antioxid. Redox Signal. 2020. [Google Scholar] [CrossRef] [PubMed]
- Polesskaya, O.; Wong, C.; Lebron, L.; Chamberlain, J.M.; Gelbard, H.A.; Goodfellow, V.; Kim, M.; Daiss, J.L.; Dewhurst, S. MLK3 regulates fMLP-stimulated neutrophil motility. Mol. Immunol. 2014, 58, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Bialopiotrowicz, E.; Gorniak, P.; Noyszewska-Kania, M.; Pula, B.; Makuch-Lasica, H.; Nowak, G.; Bluszcz, A.; Szydlowski, M.; Jablonska, E.; Piechna, K.; et al. Microenvironment-induced PIM kinases promote CXCR4-triggered mTOR pathway required for chronic lymphocytic leukaemia cell migration. J. Cell Mol. Med. 2018, 22, 3548–3559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. Ilogp: A simple, robust, and efficient description of n-octanol/water partition coefficient for drug design using the GB/SA approach. J. Chem. Inf. Model 2014, 54, 3284–3301. [Google Scholar] [CrossRef]
- Giner, B.; Lafuente, C.; Lapena, D.; Errazquin, D.; Lomba, L. Qsar study for predicting the ecotoxicity of nades towards aliivibrio fischeri. Exploring the use of mixing rules. Ecotox. Environ. Safe 2020, 191, 110004. [Google Scholar] [CrossRef]
- Ren, Y.; Yu, J.; Kinghorn, A.D. Development of anticancer agents from plant-derived sesquiterpene lactones. Curr. Med. Chem. 2016, 23, 2397–2420. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Plotnikov, M.B.; Khlebnikov, A.I.; Plotnikova, T.M.; Quinn, M.T. Oximes: Novel therapeutics with anticancer and anti-inflammatory potential. Biomolecules 2021, 11, 777. [Google Scholar] [CrossRef] [PubMed]
- Thomé, R.; de Carvalho, A.C.; Alves da Costa, T.; Ishikawa, L.L.; Fraga-Silva, T.F.; Sartori, A.; de Oliveira, A.L.; Verinaud, L. Artesunate ameliorates experimental autoimmune encephalomyelitis by inhibiting leukocyte migration to the central nervous system. CNS Neurosci. Ther. 2016, 22, 707–714. [Google Scholar] [CrossRef]
- Kiss, E.; Kins, S.; Zöller, Y.; Schilling, S.; Gorgas, K.; Groß, D.; Schlicksupp, A.; Rosner, R.; Kirsch, J.; Kuhse, J. Artesunate restores the levels of inhibitory synapse proteins and reduces amyloid-β and c-terminal fragments (ctfs) of the amyloid precursor protein in an ad-mouse model. Mol. Cell Neurosci. 2021, 113, 103624. [Google Scholar] [CrossRef] [PubMed]
- Kiss, E.; Kins, S.; Gorgas, K.; Orlik, M.; Fischer, C.; Endres, K.; Schlicksupp, A.; Kirsch, J.; Kuhse, J. Artemisinin-treatment in pre-symptomatic app-ps1 mice increases gephyrin phosphorylation at ser270: A modification regulating postsynaptic gaba(a)r density. Biol. Chem. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.R.; Tay, K.C.; Su, Y.X.; Wong, C.K.; Tan, W.N.; Khaw, K.Y. Potential of naturally derived alkaloids as multi-targeted therapeutic agents for neurodegenerative diseases. Molecules 2021, 26, 728. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Major Compounds (%) | Ref. |
|---|---|---|
| R. tomentosum | Sabinene (0–33), myrcene (0–55.7), p-cymene (0–51.7), limonene (0–50.3), γ-terpineol (0–31.2), bornyl acetate (0–10.8), ascaridol isomers (0–49.2), palustrol (0–53.5), ledol (0–36.5), lepalol (3.3–7.9), lepalone (0.7–6.5), and cyclocolorenone isomers (4.1) | [17,18,19] |
| R. anthopogonoides | 4-Phenyl-2-butanone (27.2), nerolidol (8.1), 1,4-cineole (7.9), caryophyllene (7.6), γ-elemene (6.1), α-farnesene (4.4), and spathulenol (4.2) | [20] |
| R. capitatum | Cedrene (22.2), 1,4,7,-cycloundecatriene,1,5,9,9-tetramethyl-,Z,Z,Z (18.5), α-gurjunene (5.1), α-selinene (4.8), and eremophilene (7.7) | [21] |
| R. przewalskii | Bisabolol oxide II (10.4), 4-(2,3,4,6-tetramethylphenyl)-3-buten-2-one (27.7), and manoyl oxide (10.8) | [21] |
| R. mucronulatum | Borneol (36.6), β-caryophyllene, α-humulene (15.4), and germacrene D (5.3) | [21] |
| R. micranthum R. micranthum | Germacrene D (27.6), α-humulene (6.1), α-muurolene (4.6), δ-cadinene, spathulenol (5.1), 15-copaenol (5.4), α-cadinol (6.3), and τ-muurolol (6.1) | [21] |
| R. anthopogon | α-Pinene (21.5–37.4), δ-cadinene (9.1–13.8), β-pinene (9.5–16.0), limonene (5.9–13.3), cis-ocimene (5.3), δ-amorphene (4.6), α-muurolene (4.5), and (E)-caryophyllene (3.2) | [22,23] |
| No | RRI | Compound | REOLv | REOFl | N° | RRI | Compound | REOLv | REOFl |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 1032 | α-Pinene | t | 1.2 | 33 | 1612 | β-Caryophyllene | 0.4 | |
| 2 | 1076 | Camphene | t | 0.1 | 34 | 1650 | γ-Elemene | 2.4 | 0.6 |
| 3 | 1118 | β-Pinene | t | 1.1 | 35 | 1661 | Alloaromadendrene | t | |
| 4 | 1132 | Sabinene | 0.9 | 36 | 1662 | Pulegone | t | ||
| 5 | 1174 | Myrcene | 1.2 | 37 | 1668 | (Z)-β-Farnesene | t | ||
| 6 | 1176 | α-Phellandrene | 0.6 | 38 | 1687 | α-Humulene | 0.9 | ||
| 7 | 1188 | α-Terpinene | 0.4 | 39 | 1704 | γ-Curcumene | 0.4 | ||
| 8 | 1203 | Limonene | t | 14.2 | 40 | 1719 | Borneol | 1.1 | |
| 9 | 1218 | β-Phellandrene | 8.9 | 41 | 1726 | Germacrene D | 0.5 | ||
| 10 | 1246 | (Z)-β-Ocimene | 6.5 | 42 | 1742 | β-Selinene | t | ||
| 11 | 1255 | γ-Terpinene | 7.1 | 43 | 1744 | α-Selinene | t | ||
| 12 | 1266 | (E)-β-Ocimene | 3.4 | 44 | 1755 | Bicyclogermacrene | 8.9 | 0.4 | |
| 13 | 1280 | p-Cymene | 2.8 | 45 | 1786 | ar-Curcumene | 0.6 | ||
| 14 | 1290 | Terpinolene | t | 37.7 | 46 | 1815 | 2-Tridecanone | 0.5 | |
| 15 | 1382 | cis-Alloocimene | 0.5 | 47 | 1854 | Germacrene B | 6.8 | ||
| 16 | 1398 | 2-Nonanone | t | 48 | 1886 | Curzerene | 17.8 | 2.2 | |
| 17 | 1437 | α-Thujone | t | 49 | 2050 | (E)-Nerolidol | 0.2 | ||
| 18 | 1443 | 2,5-Dimethylstyrene | 0.4 | 50 | 2096 | Elemol | 1.4 | ||
| 19 | 1451 | β-Thujone | 1.2 | 51 | 2104 | Viridiflorol | 22.0 | 1.2 | |
| 20 | 1477 | 4,8-Epoxyterpinolene | 0.9 | 52 | 2106 | β-Elemenone | 5.3 | 1.7 | |
| 21 | 1479 | δ-Elemene | t | 53 | 2144 | Spathulenol | 14.4 | 0.3 | |
| 22 | 1495 | Bicycloelemene | t | 54 | 2147 | Germacrone | 3.3 | ||
| 23 | 1528 | α-Bourbonene | t | 55 | 2198 | Thymol | 0.3 | ||
| 24 | 1535 | β-Bourbonene | t | 56 | 2199 | Alismol | 1.3 | ||
| 25 | 1536 | Italicene | t | 57 | 2203 | β-Eudesmol | 0.3 | ||
| 26 | 1541 | Benzaldehyde | t | 58 | 2219 | Porosadienol | 0.6 | ||
| 27 | 1545 | cis-α-Bergamotene | t | 59 | 2217 | Alismol isomer | 1.2 | ||
| 28 | 1553 | Linalool | 0.5 | 60 | 2368 | Eudesma-4(15),7-diene-1-β-ol | t | ||
| 29 | 1590 | Bornyl acetate | 0.6 | 0.2 | |||||
| 30 | 1600 | β-Elemene | 2.2 | 61 | 2400 | Tetracosane | 0.5 | ||
| 31 | 1604 | 2-Undecanone | 0.2 | 62 | 2500 | Pentacosane | 0.9 | ||
| 32 | 1611 | Terpinen-4-ol | 0.8 | 63 | 2656 | Furanoeremophil-1-one | t |
| Major Components | REOLv | REOFl |
|---|---|---|
| % | ||
| Monoterpene hydrocarbons | <0.1 | 87.0 |
| Oxygenated monoterpenes | 0.6 | 5.0 |
| Sesquiterpene hydrocarbons | 40.9 | 3.2 |
| Oxygenated sesquiterpenes | 50.0 | 3.2 |
| Miscellaneous compounds | 0.7 | 1.4 |
| Total | 92.2 | 99.8 |
| Essential Oil or Pure Compound | Ca2+ Influx | |||
|---|---|---|---|---|
| Neutrophils | C20 cells | |||
| EC50 (µg/mL) | ||||
| REOLv | 18.6 ± 5.8 | 22.8 ± 1.6 | ||
| REOFl | N.A. | N.A. | ||
| REOLv | REOFl | EC50 (µM) | ||
| Composition (%) | ||||
| β-Phellandrene | 0 | 8.9 | N.A. | N.A. |
| Viridiflorol | 22.0 | 1.2 | 6.8 ± 2.3 | 27.8 ± 4.6 |
| Spathulenol | 14.4 | 0.3 | 39.4 ± 9.5 | N.A. |
| Curzerene | 17.8 | 2.2 | 37.6 ± 8.4 | 25.9 ± 5.2 |
| Germacrone | 3.3 | 0 | 24.0 ± 4.6 | 27.7 ± 2.5 |
| Essential Oil or Pure Compound | FPR1- HL60 a | FPR2- HL60 b | C20 Cells a | Neutro-phils a | Neutrophils c | ||
|---|---|---|---|---|---|---|---|
| Ca2+ Influx | Chemotaxis | ||||||
| IC50 (µg/mL) | |||||||
| REOLv | 12.3 ± 2.5 | 7.6 ± 2.3 | 8.0 ± 0.1 | 2.7 ± 0.6 | 3.3 ± 0.5 | ||
| REOFl | N.A. | N.A. | N.A. | N.A. | N.A. | ||
| Composition (%) | |||||||
| REOLv | REOFl | IC50 (µM) | |||||
| β-Phellandrene | 0 | 8.9 | N.A. | N.A. | N.A. | N.A. | N.A. |
| Viridiflorol | 22.0 | 1.2 | 19.5 ± 4.7 | 10.7 ± 3.8 | 22.6 ± 3.1 | 7.8 ± 2.3 | 18.3 ± 4.1 |
| Spathulenol | 14.4 | 0.3 | 32.2 ± 6.4 | 31.6 ± 5.3 | 9.8 ± 3.4 | 36.2 ± 8.2 | 4.9 ± 0.8 |
| Curzerene | 17.8 | 2.2 | 21.8 ± 6.1 | 16.7 ± 5.5 | 30.7 ± 4.4 | 11.0 ± 3.8 | 37.9 ± 2.2 |
| Germacrone | 3.3 | 0 | 27.7 ± 2.9 | 25.0 ± 7.2 | 10.7 ± 2.3 | 27.9 ± 8.9 | 8.5 ± 0.6 |
| Rank | PDB ID | Target Name | Fit Score | Rank | PDB ID | Target Name | Fit Score |
|---|---|---|---|---|---|---|---|
| Germacrene B | Germacrone | ||||||
| 1 | 1J96 | AKR1C2 | 0.9912 | 1 | 1J96 | AKR1C2 | 0.9926 |
| 2 | 1REU | BMP2 | 0.9846 | 2 | 1PMV | JNK3 | 0.9911 |
| 3 | 2P3G | MAPKAPK2 | 0.9817 | 3 | 1UKI | JNK1 | 0.9909 |
| 4 | 2PG2 | KIF11 | 0.9735 | 4 | 2PIN | NR1A2 | 0.9712 |
| 5 | 1P49 | Steroid sulfatase | 0.9567 | 5 | 1P49 | Steroid sulfatase | 0.9586 |
| 6 | 1SHJ | Caspase-7 | 0.9481 | 6 | 2PG2 | KIF11 | 0.9537 |
| 7 | 1E7E | Serum albumin | 0.9419 | 7 | 1L6L | Apo A-II | 0.9489 |
| 8 | 2O65 | PIM1 | 0.9295 | 8 | 1RS0 | CFB | 0.9415 |
| (−)-Curzerene | (+)-Viridiflorol | ||||||
| 1 | 1REU | BMP2 | 0.9904 | 1 | 1XDD | Integrin α-L | 3 |
| 2 | 2PIN | NR1A2 | 0.9873 | 2 | 1J96 | AKR1C2 | 3 |
| 3 | 2O65 | PIM1 | 0.9861 | 3 | 3BMP | BMP2 | 3 |
| 4 | 2P3G | MAPKAPK2 | 0.9764 | 4 | 2P3G | MAPKAPK2 | 2.906 |
| 5 | 1UKI | JNK1 | 0.975 | 5 | 1IF4 | CA2 | 2.886 |
| 6 | 1PMV | JNK3 | 0.9671 | 6 | 3CJF | VEGFR2 | 2.881 |
| 7 | 1RS0 | CFB | 0.9652 | 7 | 1SHJ | Caspase-7 | 2.708 |
| 8 | 1SHJ | Caspase-7 | 0.9594 | 8 | 3CGF | JNK3 | 2.568 |
| (−)-Spathulenol | (+)-Spathulenol | ||||||
| 1 | 1XDD | Integrin α-L | 2.968 | 1 | 2P3G | MAPKAPK2 | 2.952 |
| 2 | 1NO9 | Prothrombin | 2.948 | 2 | 3BMP | BMP2 | 2.947 |
| 3 | 3BMP | BMP2 | 2.947 | 3 | 1XDD | Integrin α-L | 2.895 |
| 4 | 1J96 | AKR1C2 | 2.921 | 4 | 1NO9 | Prothrombin | 2.802 |
| 5 | 1E7A | Serum albumin | 2.804 | 5 | 1SHJ | Caspase-7 | 2.793 |
| 6 | 1PMV | JNK3 | 2.789 | 6 | 1PMV | JNK3 | 2.736 |
| 7 | 2P3G | MAPKAPK2 | 2.749 | 7 | 2O65 | PIM1 | 2.723 |
| 8 | 1P49 | Steroid sulfatase | 2.739 | 8 | 1J96 | AKR1C2 | 2.722 |
| Property | Germacrene B | Germacrone | Curzerene | Viridiflorol | Spathulenol |
|---|---|---|---|---|---|
| Formula | C15H24 | C15H22O | C15H20O | C15H26O | C15H24O |
| M.W. | 204.35 | 218.33 | 216.32 | 222.37 | 220.35 |
| Heavy atoms | 15 | 16 | 16 | 16 | 16 |
| Fraction Csp3 | 0.60 | 0.53 | 0.47 | 1.00 | 0.87 |
| Rotatable bonds | 0 | 0 | 2 | 0 | 0 |
| H-bond acceptors | 0 | 1 | 1 | 1 | 1 |
| H-bond donors | 0 | 1 | 0 | 1 | 1 |
| MR | 70.68 | 70.88 | 68.74 | 68.82 | 68.34 |
| tPSA | 0.00 | 17.07 | 13.14 | 20.23 | 20.23 |
| iLogP | 3.27 | 2.97 | 3.10 | 3.08 | 3.04 |
| BBB permeation | No | Yes | Yes | Yes | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical Composition and Immunomodulatory Activity of Essential Oils from Rhododendron albiflorum. Molecules 2021, 26, 3652. https://doi.org/10.3390/molecules26123652
Schepetkin IA, Özek G, Özek T, Kirpotina LN, Khlebnikov AI, Quinn MT. Chemical Composition and Immunomodulatory Activity of Essential Oils from Rhododendron albiflorum. Molecules. 2021; 26(12):3652. https://doi.org/10.3390/molecules26123652
Chicago/Turabian StyleSchepetkin, Igor A., Gulmira Özek, Temel Özek, Liliya N. Kirpotina, Andrei I. Khlebnikov, and Mark T. Quinn. 2021. "Chemical Composition and Immunomodulatory Activity of Essential Oils from Rhododendron albiflorum" Molecules 26, no. 12: 3652. https://doi.org/10.3390/molecules26123652
APA StyleSchepetkin, I. A., Özek, G., Özek, T., Kirpotina, L. N., Khlebnikov, A. I., & Quinn, M. T. (2021). Chemical Composition and Immunomodulatory Activity of Essential Oils from Rhododendron albiflorum. Molecules, 26(12), 3652. https://doi.org/10.3390/molecules26123652