
Origanum vulgare ssp. hirtum (Lamiaceae) Essential Oil Prevents Behavioral and Oxidative Stress Changes in the Scopolamine Zebrafish Model




Abstract
:1. Introduction
2. Results and Discussion
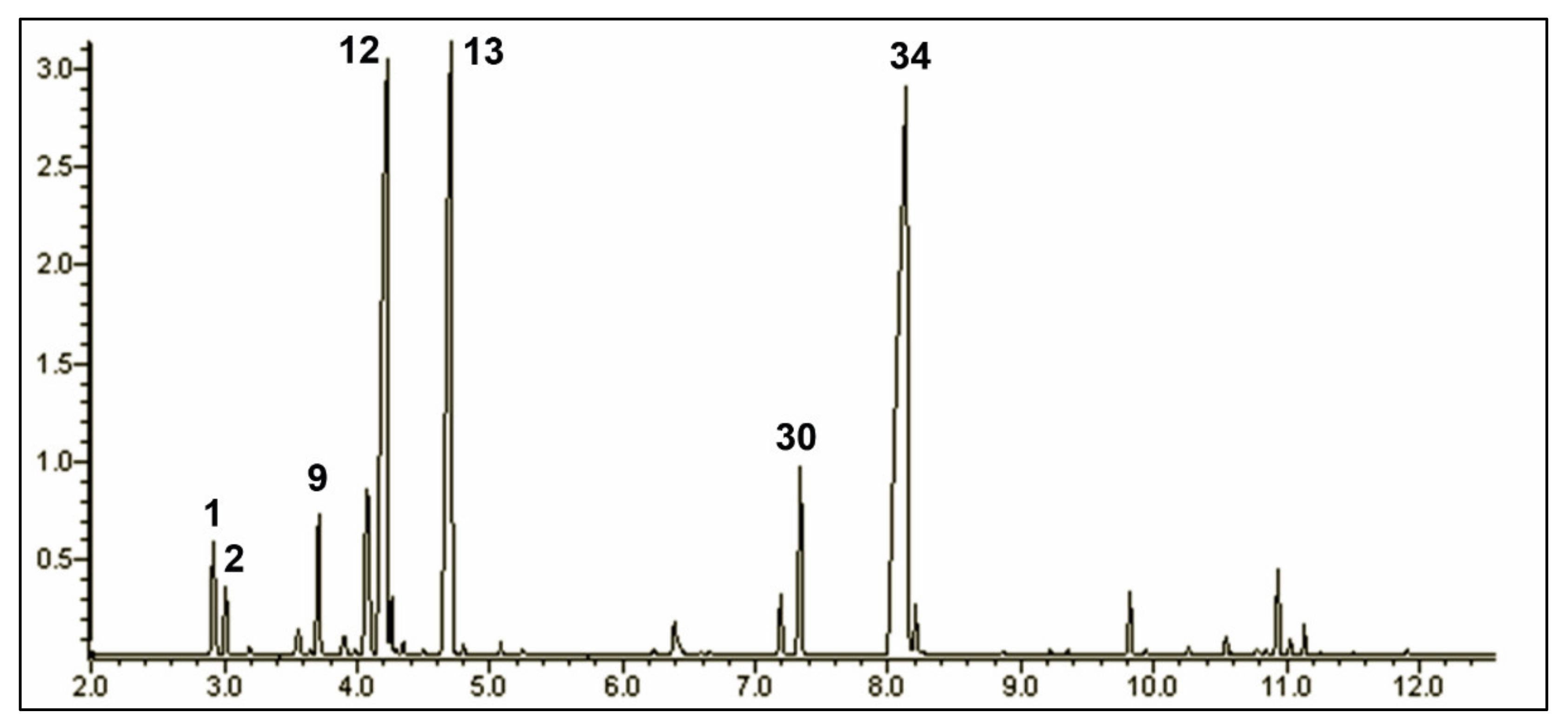
2.1. The Chemical Composition of the Origanum vulgare ssp. hirtum Essential Oil
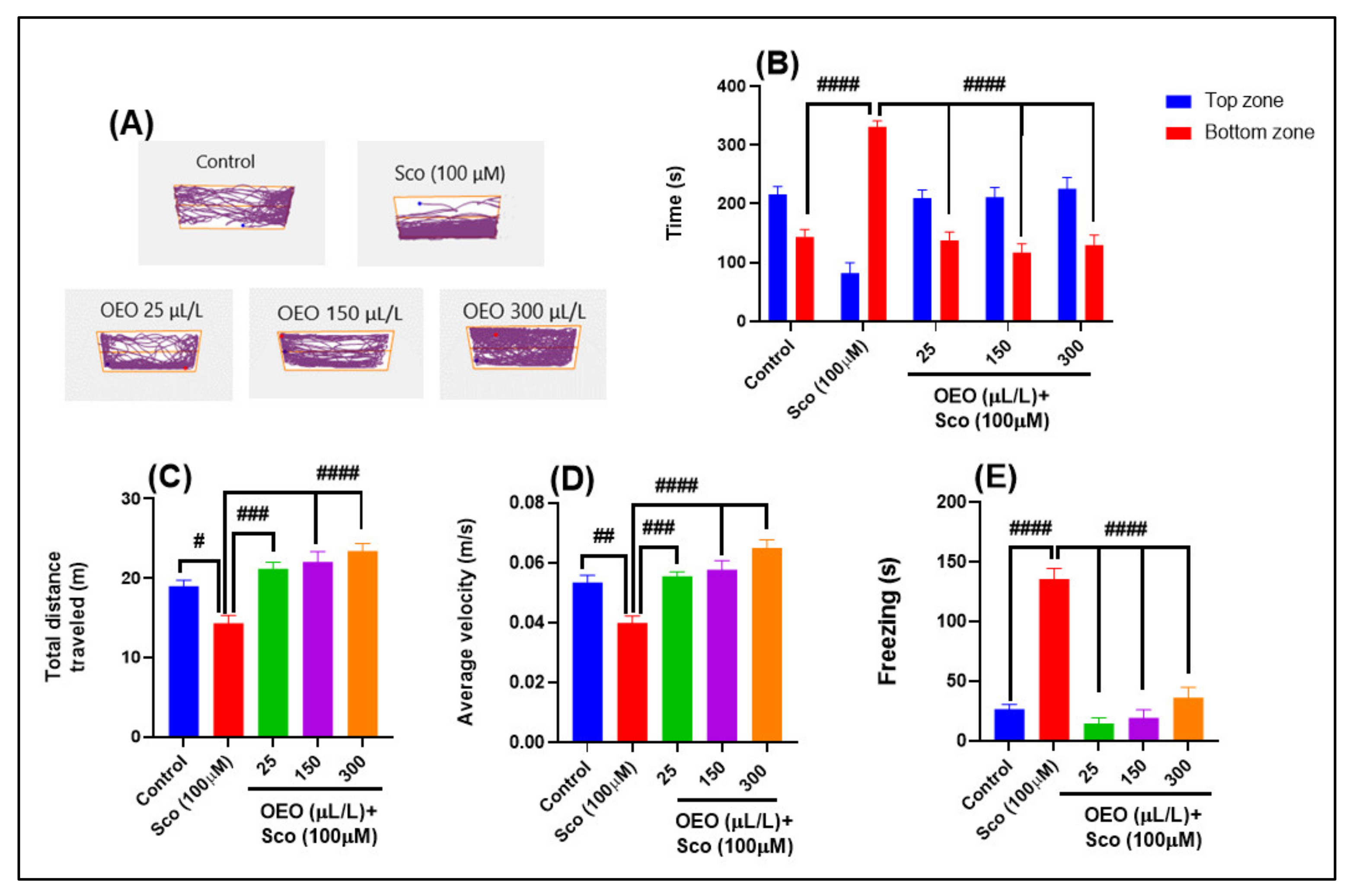
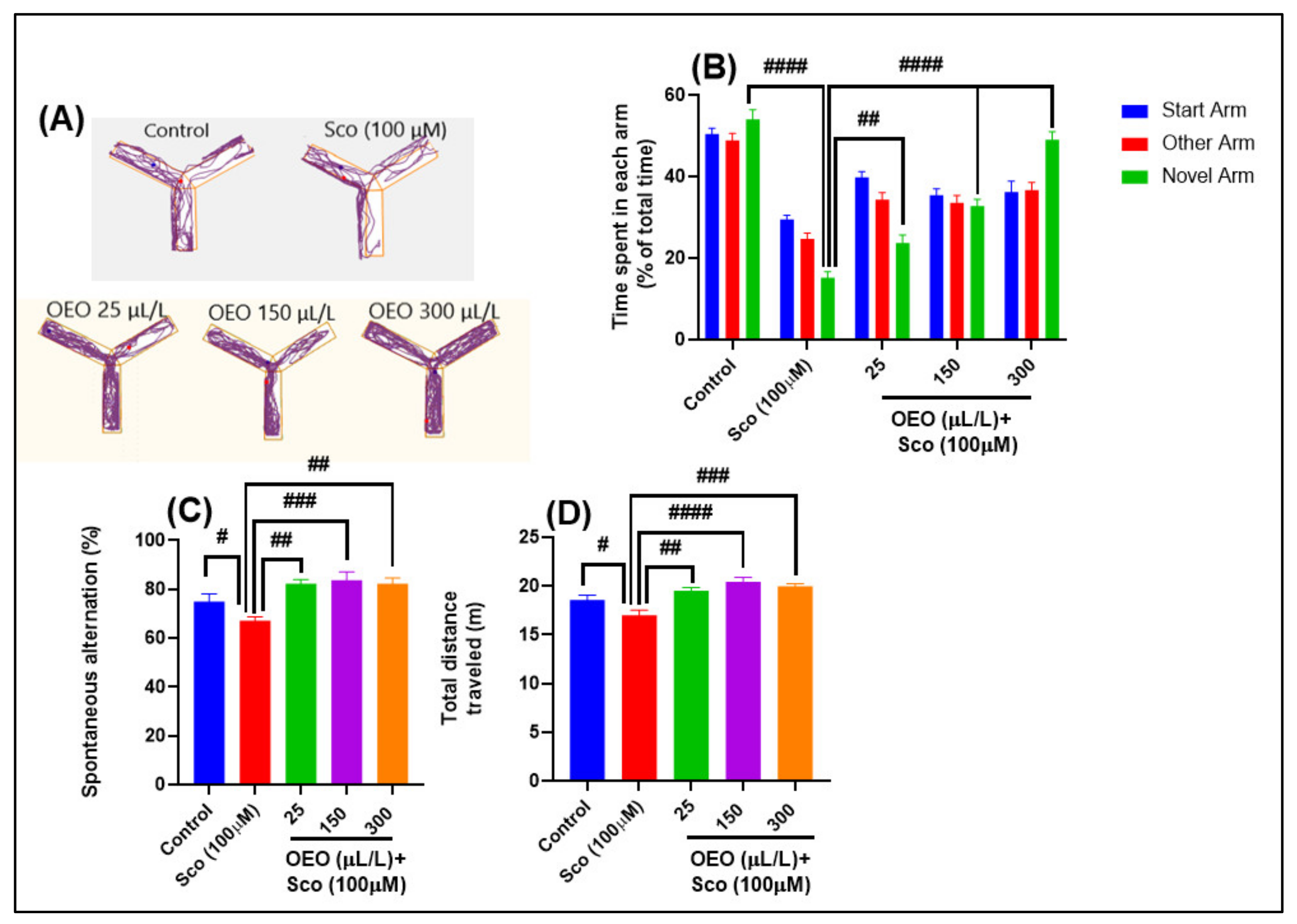
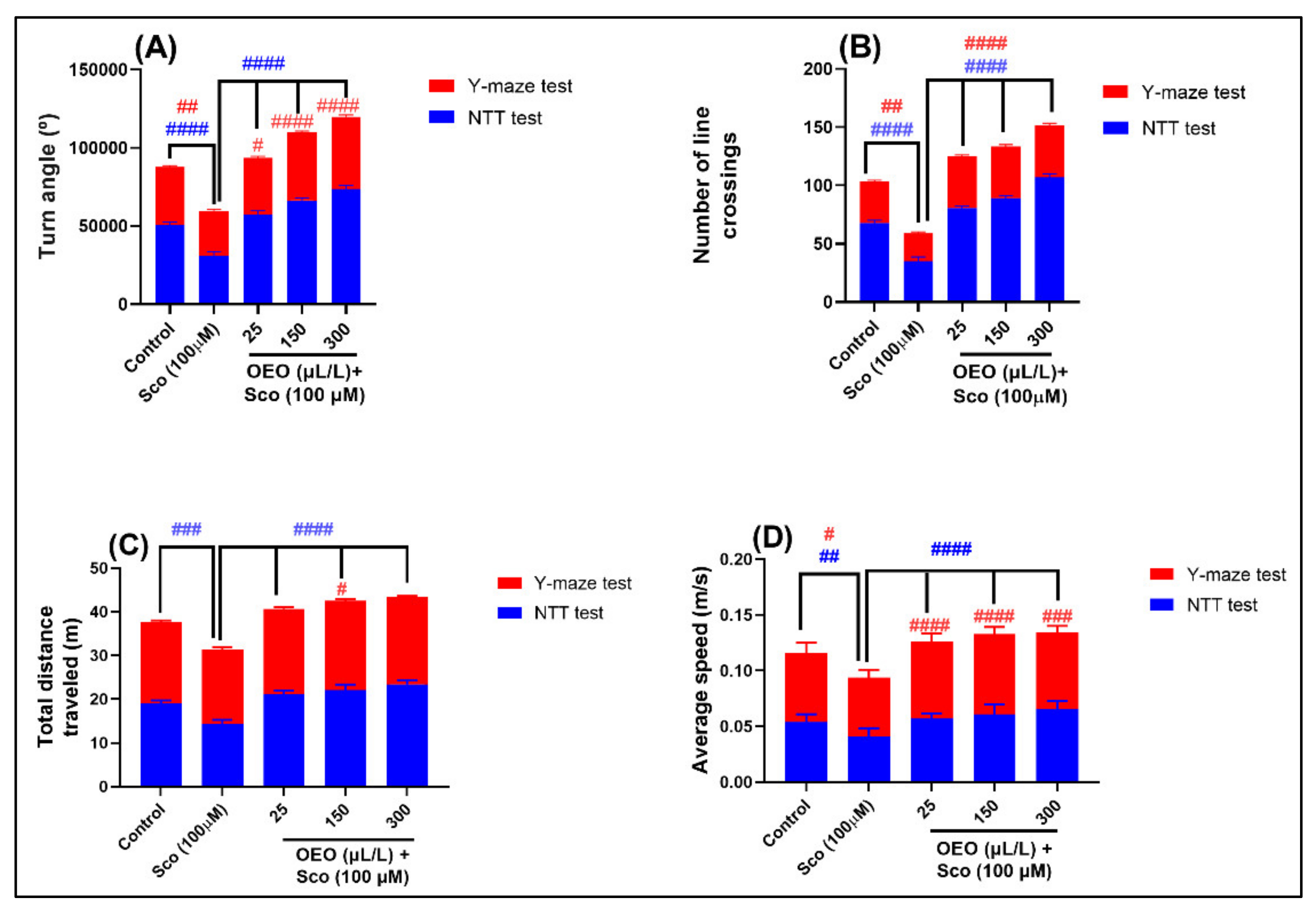
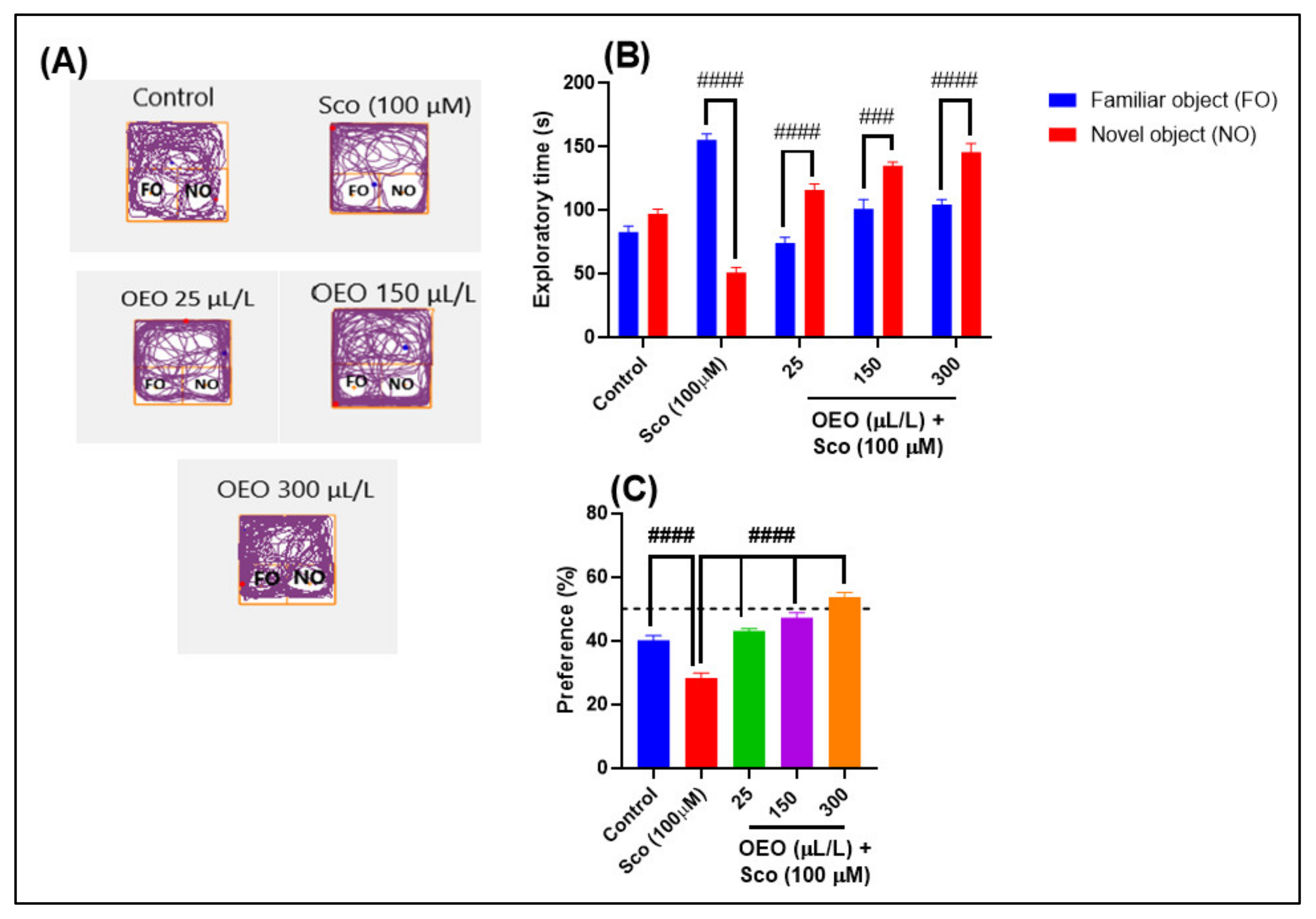
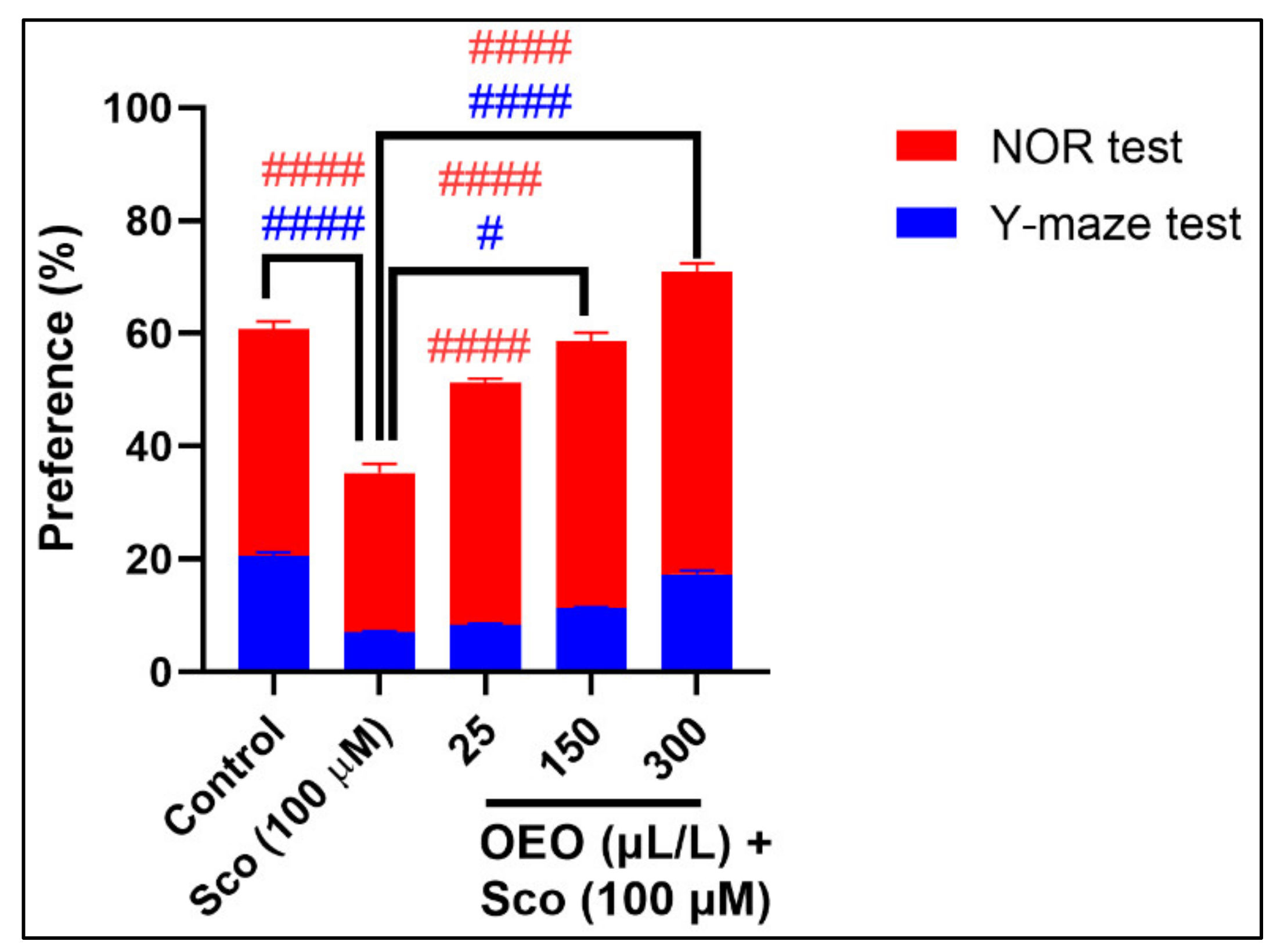
2.2. Improvement of Anxiety-Like Behavior, Spatial Memory, and Response to Novelty in the NTT, Y-Maze and NOR Tests
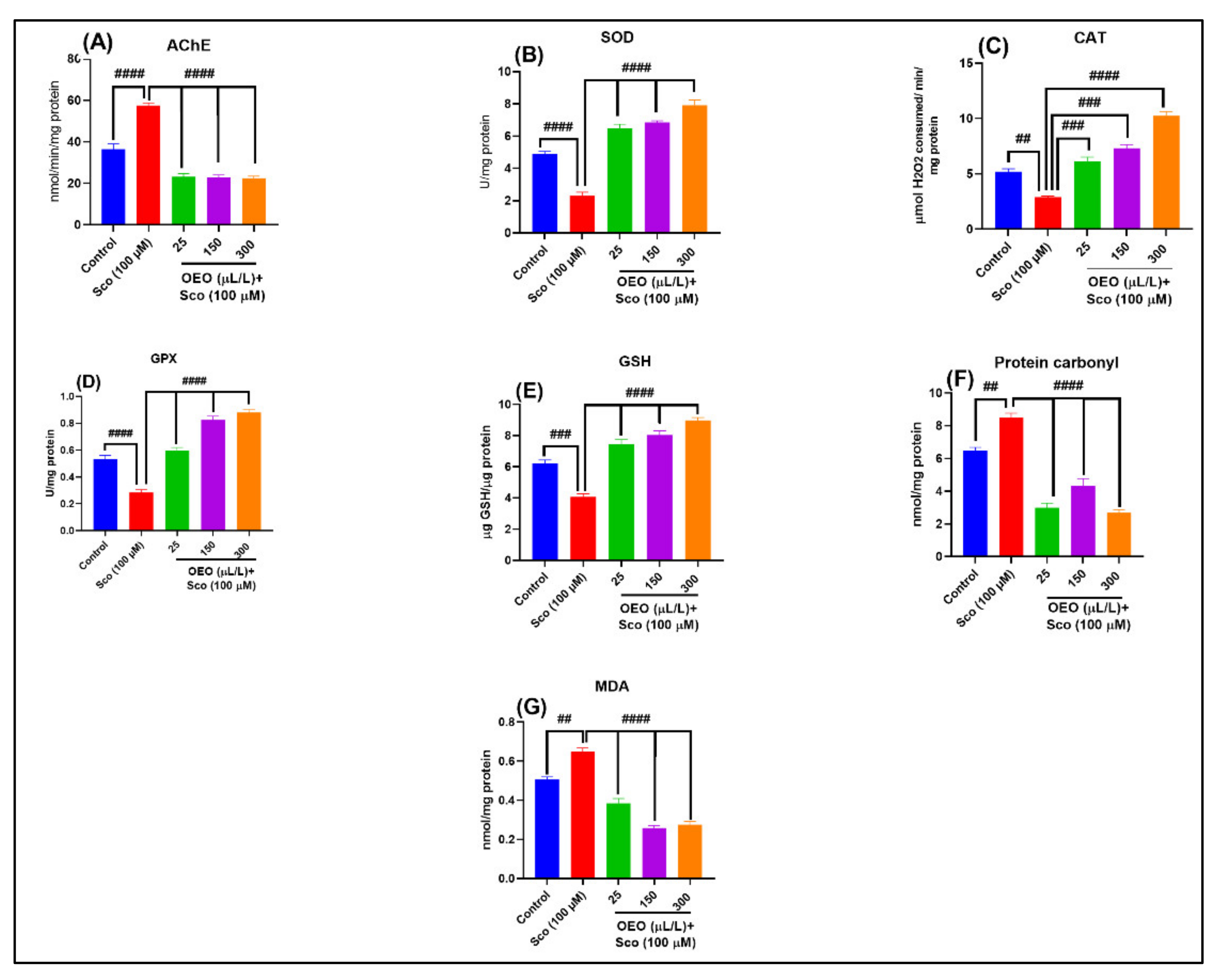
2.3. In Vivo Inhibitory Activity against Acetylcholinesterase Activity
2.4. In Vivo Antioxidant Activity
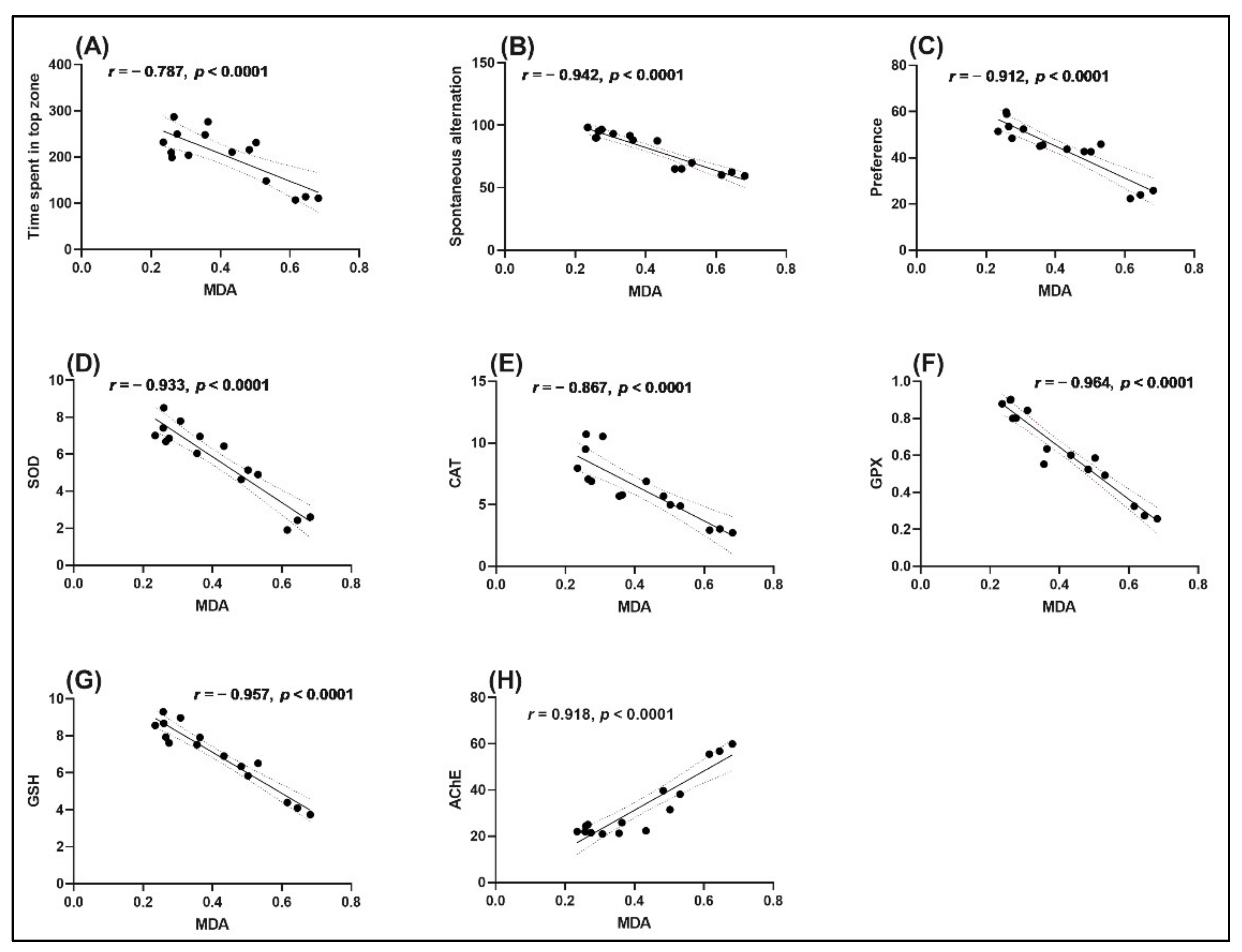
2.5. Correlation between Behavioral Scores, Enzymatic Activities, and Lipid Peroxidation
3. Materials and Methods
3.1. Essential Oil and Chemical Material
3.2. Gas Chromatograph–Mass Spectrometry (GC-MS) Analysis
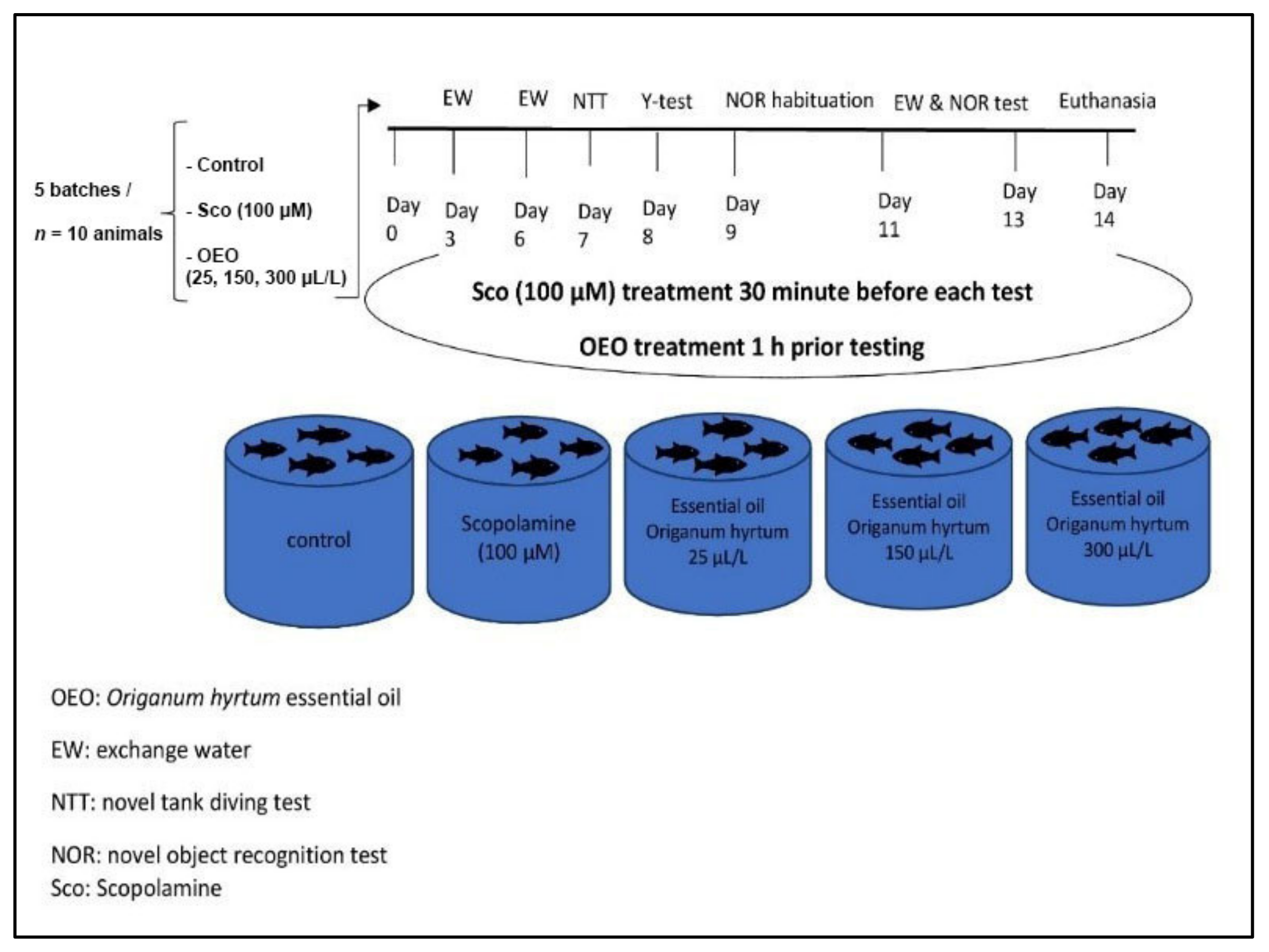
3.3. Zebrafish and Treatment
3.4. Novel Tank-Diving Test (NTT)
3.5. Y-Maze
3.6. Novel Object Recognition Test (NOR)
3.7. Biochemical Assays
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thawkar, B.S.; Kaur, G. Zebrafish as a promising tool for modeling neurotoxin-induced Alzheimer’s disease. Neurotox. Res. 2021, 39, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Capatina, L.; Todirascu-Ciornea, E.; Napoli, E.M.; Ruberto, G.; Hritcu, L.; Dumitru, G. Thymus vulgaris essential oil protects zebrafish against cognitive dysfunction by regulating cholinergic and antioxidants systems. Antioxidants 2020, 9, 1083. [Google Scholar] [CrossRef] [PubMed]
- Brinza, I.; Abd-Alkhalek, A.M.; El-Raey, M.A.; Boiangiu, R.S.; Eldahshan, O.A.; Hritcu, L. Ameliorative effects of rhoifolin in scopolamine-induced amnesic zebrafish (Danio rerio) model. Antioxidants 2020, 9, 580. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Kannan, R.R. Zebrafish: An emerging real-time model system to study Alzheimer’s disease and neurospecific drug discovery. Cell Death Discov. 2018, 4, 45. [Google Scholar] [CrossRef] [Green Version]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of zebrafish in drug discovery toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Jahanshahi, M.; Azami, N.S.; Nickmahzar, E. Effect of scopolamine-based amnesia on the number of astrocytes in the rat’s hippocampus. Int. J. Morphol. 2012, 30, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Pagnussat, N.; Almeida, A.S.; Marques, D.M.; Nunes, F.; Chenet, G.C.; Botton, P.H.S.; Mioranzza, S.; Loss, C.M.; Cunha, R.A.; Porciúncula, L.O. Adenosine A 2 A receptors are necessary and sufficient to trigger memory impairment in adult mice. Br. J. Pharmacol. 2015, 172, 3831–3845. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.-Y.; Jang, M.J.; Choi, Y.-H.; Hwang, H.; Rhim, H.; Lee, B.; Choi, C.W.; Kim, M.S. Central administration of afzelin extracted from Ribes fasciculatum improves cognitive and memory function in a mouse model of dementia. Sci. Rep. 2021, 11, 9182. [Google Scholar] [CrossRef]
- Aydin, E.; Hritcu, L.; Dogan, G.; Hayta, S.; Bagci, E. The Effects of inhaled Pimpinella peregrina essential oil on scopolamine-induced memory impairment, anxiety, and depression in laboratory rats. Mol. Neurobiol. 2016, 53, 6557–6567. [Google Scholar] [CrossRef]
- Mitchnick, K.A.; Wideman, C.E.; Huff, A.E.; Palmer, D.; McNaughton, B.L.; Winters, B.D. Development of novel tasks for studying view-invariant object recognition in rodents: Sensitivity to scopolamine. Behav. Brain Res. 2018, 344, 48–56. [Google Scholar] [CrossRef]
- Valu, M.-V.; Soare, L.C.; Ducu, C.; Moga, S.; Negrea, D.; Vamanu, E.; Balseanu, T.-A.; Carradori, S.; Hritcu, L.; Boiangiu, R.S. Hericium erinaceus (Bull.) Pers. ethanolic extract with antioxidant properties on scopolamine-induced memory deficits in a zebrafish model of cognitive impairment. J. Fungi 2021, 7, 477. [Google Scholar] [CrossRef]
- Valu, M.-V.; Ducu, C.; Moga, S.; Negrea, D.; Hritcu, L.; Boiangiu, R.S.; Vamanu, E.; Balseanu, T.A.; Carradori, S.; Soare, L.C. Effects of the Hydroethanolic Extract of Lycopodium selago L. on scopolamine-induced memory deficits in zebrafish. Pharmaceuticals 2021, 14, 568. [Google Scholar] [CrossRef]
- Sahin, F.; Güllüce, M.; Daferera, D.; Sökmen, A.; Sökmen, M.; Polissiou, M.; Agar, G.; Özer, H. Biological activities of the essential oils and methanol extract of Origanum vulgare ssp. vulgare in the Eastern Anatolia region of Turkey. Food Control 2004, 15, 549–557. [Google Scholar] [CrossRef]
- Pezzani, R.; Vitalini, S.; Iriti, M. Bioactivities of Origanum vulgare L.: An update. Phytochem. Rev. 2017, 16, 1253–1268. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; del Contreras, M.M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phyther. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef]
- Asadbegi, M.; Yaghmaei, P.; Salehi, I.; Komaki, A.; Ebrahim-Habibi, A. Investigation of thymol effect on learning and memory impairment induced by intrahippocampal injection of amyloid beta peptide in high fat diet- fed rats. Metab. Brain Dis. 2017, 32, 827–839. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential oils of oregano: Biological activity beyond their antimicrobial properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.L.; Guo, Y.S.; Wang, C.H.; Li, G.Q.; Xu, J.J.; Chung, H.Y.; Ye, W.C.; Li, Y.L.; Wang, G.C. Phenolic compounds from Origanum vulgare and their antioxidant and antiviral activities. Food Chem. 2014, 152, 300–306. [Google Scholar] [CrossRef]
- Ivanova, D.; Gerova, D.; Chervenkov, T.; Yankova, T. Polyphenols and antioxidant capacity of Bulgarian medicinal plants. J. Ethnopharmacol. 2005, 96, 145–150. [Google Scholar] [CrossRef]
- Avola, R.; Granata, G.; Geraci, C.; Napoli, E.; Graziano, A.C.E.; Cardile, V. Oregano (Origanum vulgare L.) essential oil provides anti-inflammatory activity and facilitates wound healing in a human keratinocytes cell model. Food Chem. Toxicol. 2020, 144, 111586. [Google Scholar] [CrossRef]
- Han, X.; Parker, T.L. Anti-inflammatory, tissue remodeling, immunomodulatory, and anticancer activities of oregano (Origanum vulgare) essential oil in a human skin disease model. Biochim. Open 2017, 4, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Vujicic, M.; Nikolic, I.; Kontogianni, V.G.; Saksida, T.; Charisiadis, P.; Vasic, B.; Stosic-Grujicic, S.; Gerothanassis, I.P.; Tzakos, A.G.; Stojanovic, I. Ethyl Acetate Extract of Origanum vulgare L. ssp. hirtum prevents streptozotocin-induced diabetes in C57BL/6 mice. J. Food Sci. 2016, 81, H1846–H1853. [Google Scholar] [CrossRef] [PubMed]
- Kosakowska, O.; Węglarz, Z.; Pióro-Jabrucka, E.; Przybył, J.L.; Kraśniewska, K.; Gniewosz, M.; Bączek, K. Antioxidant and antibacterial activity of essential oils and hydroethanolic extracts of greek oregano (O. vulgare L. subsp. hirtum (Link) Ietswaart) and common oregano (O. vulgare L. subsp. vulgare). Molecules 2021, 26, 988. [Google Scholar] [CrossRef] [PubMed]
- Kakhki, M.T.; Sedaghat, N.; Mohsenzadeh, M. Chemical composition, antioxidative, antibacterial, and time-kill activities of some selected plant essential oils against foodborne pathogenic and spoilage organisms. Vet. Res. Forum 2020, 11, 339. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Ramos, C.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.F.; Saraiva, J.A.; Nunes, M.L. Chemical composition and bioactivity of different oregano (Origanum vulgare) extracts and essential oil. J. Sci. Food Agric. 2013, 93, 2707–2714. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Y.H.; Wang, K.B.; Fan, L.M.; Su, F.W.; Ye, M. Chemical composition and allelopathic potential of essential oil isolated from Origanum vulgare. J. Appl. Ecol. 2020, 31, 2257–2263. [Google Scholar] [CrossRef]
- Hamada, I.; Al-Waili, N.; Aboulghazi, A.; Abdellaoui, A.; Al-Waili, T.; Lyoussi, B. Chemical composition and antioxidant content of Thymus vulgaris honey and Origanum vulgare essential oil; their effect on carbon tetrachloride-induced toxicity. Vet. World 2021, 14, 292–301. [Google Scholar] [CrossRef]
- Amaral, S.C.; Pruski, B.B.; de Freitas, S.B.; Allend, S.O.; Ferreira, M.R.A.; Moreira, C.; Pereira, D.I.B.; Junior, A.S.V.; Hartwig, D.D. Origanum vulgare essential oil: Antibacterial activities and synergistic effect with polymyxin B against multidrug-resistant Acinetobacter baumannii. Mol. Biol. Rep. 2020, 47, 9615–9625. [Google Scholar] [CrossRef]
- Azizi, Z.; Ebrahimi, S.; Saadatfar, E.; Kamalinejad, M.; Majlessi, N. Cognitive-enhancing activity of thymol and carvacrol in two rat models of dementia. Behav. Pharmacol. 2012, 23, 241–249. [Google Scholar] [CrossRef]
- Sudeep, H.V.; Venkatakrishna, K.; Amritharaj; Gouthamchandra, K.; Reethi, B.; Naveen, P.; Lingaraju, H.B.; Shyamprasad, K. A standardized black pepper seed extract containing β-caryophyllene improves cognitive function in scopolamine-induced amnesia model mice via regulation of brain-derived neurotrophic factor and MAPK proteins. J. Food Biochem. 2021, e13994. [Google Scholar] [CrossRef]
- Napoli, E.; Siracusa, L.; Ruberto, G. New Tricks for Old Guys: Recent developments in the chemistry, biochemistry, applications and exploitation of selected species from the Lamiaceae family. Chem. Biodivers. 2020, 17, e1900677. [Google Scholar] [CrossRef]
- Napoli, E.; Giovino, A.; Carrubba, A.; How Yuen Siong, V.; Rinoldo, C.; Nina, O.; Ruberto, G. Variations of essential oil constituents in oregano (Origanum vulgare subsp. viridulum (= O. heracleoticum) over cultivation cycles. Plants 2020, 9, 1174. [Google Scholar] [CrossRef]
- Licata, M.; Tuttolomondo, T.; Dugo, G.; Ruberto, G.; Leto, C.; Napoli, E.M.; Rando, R.; Fede, M.R.; Virga, G.; Leone, R.; et al. Study of quantitative and qualitative variations in essential oils of Sicilian oregano biotypes. J. Essent. Oil Res. 2015, 27, 293–306. [Google Scholar] [CrossRef]
- Mancini, E.; Camele, I.; Elshafie, H.S.; De Martino, L.; Pellegrino, C.; Grulova, D.; De Feo, V. Chemical composition and biological activity of the essential oil of Origanum vulgare ssp. hirtum from different areas in the southern Apennines (Italy). Chem. Biodivers. 2014, 11, 639–651. [Google Scholar] [CrossRef]
- Stešević, D.; Jaćimović, Ž.; Šatović, Z.; Šapčanin, A.; Jančan, G.; Kosović, M.; Damjanović-Vratnica, B. Chemical characterization of wild growing Origanum vulgare populations in Montenegro. Nat. Prod. Commun. 2018, 13, 1934578X1801301. [Google Scholar] [CrossRef] [Green Version]
- Pasias, I.N.; Ntakoulas, D.D.; Raptopoulou, K.; Gardeli, C.; Proestos, C. Chemical composition of essential oils of aromatic and medicinal herbs cultivated in Greece—Benefits and drawbacks. Foods 2021, 10, 2354. [Google Scholar] [CrossRef]
- Qiao, Y.; Yu, Z.; Bai, L.; Li, H.; Zhang, S.; Liu, J.; Gao, Z.; Yang, X. Chemical composition of essential oils from Thymus mongolicus, Cinnamomum verum, and Origanum vulgare and their acaricidal effects on Haemaphysalis longicornis (Acari: Ixodidae). Ecotoxicol. Environ. Saf. 2021, 224, 112672. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Y.H.; Ye, M.; Wang, K.B.; Fan, L.M.; Su, F.W. Chemical composition and antifungal activity of essential oil from Origanum vulgare against Botrytis cinerea. Food Chem. 2021, 365, 130506. [Google Scholar] [CrossRef]
- Wong, K.; Elegante, M.; Bartels, B.; Elkhayat, S.; Tien, D.; Roy, S.; Goodspeed, J.; Suciu, C.; Tan, J.; Grimes, C.; et al. Analyzing habituation responses to novelty in zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 450–457. [Google Scholar] [CrossRef]
- Blaser, R.E.; Rosemberg, D.B. Measures of anxiety in zebrafish (Danio rerio): Dissociation of black/white preference and novel tank test. PLoS ONE 2012, 7, e36931. [Google Scholar] [CrossRef] [Green Version]
- Kalueff, A.V.; Cachat, J.M. Zebrafish Models in Neurobehavioral Research; Humana Press: Totowa, NJ, USA, 2011. [Google Scholar]
- Amiresmaeili, A.; Roohollahi, S.; Mostafavi, A.; Askari, N. Effects of oregano essential oil on brain TLR4 and TLR2 gene expression and depressive-like behavior in a rat model. Res. Pharm. Sci. 2018, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Abbasi-Maleki, S.; Kadkhoda, Z.; Taghizad-Farid, R. The antidepressant-like effects of Origanum majorana essential oil on mice through monoaminergic modulation using the forced swimming test. J. Tradit. Complement. Med. 2020, 10, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, A.; Mousavi, G.; Nazeri, M.; Jafari, B.; Ebadi, A.; Ahmadeh, C.; Habibi, E. Comparative study of sedative, pre-anesthetic and anti-anxiety effect of Origanum majorana extract with diazepam on rats. Res. J. Biol. Sci. 2011, 6, 611–614. [Google Scholar] [CrossRef] [Green Version]
- Mombeini, T.; Mazloumi, S.; Shams, J. Pharmacological effects of Origanum vulgare L. in the elevated plus-maze and open field tests in the rat. J. Basic Clin. Pathophysiol. 2015, 3, 29–36. [Google Scholar]
- Da Machado, K.C.; Paz, M.F.C.J.; de Oliveira Santos, J.V.; da Silva, F.C.C.; Tchekalarova, J.D.; Salehi, B.; Islam, M.T.; Setzer, W.N.; Sharifi-Rad, J.; de Castro e Sousa, J.M.; et al. Anxiety therapeutic interventions of β-caryophyllene: A laboratory-based study. Nat. Prod. Commun. 2020, 15, 1934578X2096222. [Google Scholar] [CrossRef]
- Hanrahan, J.R.; Chebib, M.; Johnston, G.A.R. Flavonoid modulation of GABA A receptors. Br. J. Pharmacol. 2011, 163, 234–245. [Google Scholar] [CrossRef] [Green Version]
- Bianchini, A.E.; Garlet, Q.I.; Da Cunha, J.A.; Bandeira Junior, G.; Brusque, I.C.M.; Salbego, J.; Heinzmann, B.M.; Baldisserotto, B. Monoterpenoids (thymol, carvacrol and S-(+)-linalool) with anesthetic activity in silver catfish (Rhamdia quelen): Evaluation of acetylcholinesterase and GABAergic activity. Brazilian J. Med. Biol. Res. 2017, 50, e6346. [Google Scholar] [CrossRef] [Green Version]
- Dougnon, G.; Ito, M. Role of Ascaridole and p-cymene in the sleep-promoting effects of Dysphania ambrosioides essential oil via the GABAergic system in a ddY mouse inhalation model. J. Nat. Prod. 2021, 84, 91–100. [Google Scholar] [CrossRef]
- Wang, Z.J.; Heinbockel, T. Essential oils and their constituents targeting the GABAergic system and sodium channels as treatment of neurological diseases. Molecules 2018, 23, 1061. [Google Scholar] [CrossRef] [Green Version]
- Brinza, I.; Boiangiu, R.S.; Hancianu, M.; Cioanca, O.; Orhan, I.E.; Hritcu, L. Bay leaf (Laurus nobilis L.) incense improved scopolamine-induced amnesic rats by restoring cholinergic dysfunction and brain antioxidant status. Antioxidants 2021, 10, 259. [Google Scholar] [CrossRef]
- Gaspary, K.V.; Reolon, G.K.; Gusso, D.; Bonan, C.D. Novel object recognition and object location tasks in zebrafish: Influence of habituation and NMDA receptor antagonism. Neurobiol. Learn. Mem. 2018, 155, 249–260. [Google Scholar] [CrossRef]
- Ghaderi, A.; Karimi, S.A.; Talaei, F.; Shahidi, S.; Faraji, N.; Komaki, A. The effects of aqueous extract of Origanum vulgare on learning and memory in male rats. J. Herbmed Pharmacol. 2020, 9, 239–244. [Google Scholar] [CrossRef]
- Haghpanah, T.; Bezanjani, E.K.; Khaki, M.R.A.; Sheibani, V.; Abbasnejad, M.; Ardakani, Y.M. Effect of intra-hippocampal injection of Origanum vulgare L. ssp. viridis leaf extract on spatial learning and memory consolidation. KAUMS J. (FEYZ) 2011, 14, 380–387. [Google Scholar]
- Maryam, A.; Farhad, V.; Bagher, S. Investigation of Origanum vulgare L. Leaf Extract on Ethanol-Induced Impairment of Working Memory on Male Rat; Civilica: Teheran, Iran, 2021; Available online: https://civilica.com/doc/976478/ (accessed on 1 October 2021).
- Sheibani, V.; Afarinesh, M.; Hajializadeh, Z.; Abbasnejad, M.; Haghpanah, T.; Arabnezhad, R.; Sepehri, G. Evaluation of Origanum vulgare L. ssp. viridis leaves extract effect on discrimination learning and LTP induction in the CA1 region of the rat hippocampus. Iran. J. Basic Med. Sci. 2011, 14, 177–184. [Google Scholar] [CrossRef]
- Seifi-Nahavandi, B.; Yaghmaei, P.; Ahmadian, S.; Ghobeh, M.; Ebrahim-Habibi, A. Cymene consumption and physical activity effect in Alzheimer’s disease model: An in vivo and in vitro study. J. Diabetes Metab. Disord. 2020, 19, 1381–1389. [Google Scholar] [CrossRef]
- Kim, K.; Bu, Y.; Jeong, S.; Lim, J.; Kwon, Y.; Cha, D.S.; Kim, J.; Jeon, S.; Eun, J.; Jeon, H. Memory-enhancing effect of a supercritical carbon dioxide fluid extract of the needles of Abies koreana on scopolamine-induced amnesia in mice. Biosci. Biotechnol. Biochem. 2006, 70, 1821–1826. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, A.; Perez, E.; Cutillas, A.-B.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Origanum vulgare and Thymbra capitata essential oils from Spain: Determination of aromatic profile and bioactivities. Nat. Prod. Commun. 2016, 11, 113–120. [Google Scholar] [CrossRef]
- Lombrea, A.; Antal, D.; Ardelean, F.; Avram, S.; Pavel, I.Z.; Vlaia, L.; Mut, A.-M.; Diaconeasa, Z.; Dehelean, C.A.; Soica, C.; et al. A Recent insight regarding the phytochemistry and bioactivity of Origanum vulgare L. essential oil. Int. J. Mol. Sci. 2020, 21, 9653. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Zengin, G.; Oskay, M.; Uysal, S.; Ceylan, R.; Aktumsek, A. Composition, antioxidant, antimicrobial and enzyme inhibition activities of two Origanum vulgare subspecies (subsp. vulgare and subsp. hirtum) essential oils. Ind. Crops Prod. 2015, 70, 178–184. [Google Scholar] [CrossRef]
- Sohn, E.; Kim, Y.J.; Kim, J.-H.; Jeong, S.-J. Ficus erecta Thunb leaves alleviate memory loss induced by scopolamine in mice via regulation of oxidative stress and cholinergic system. Mol. Neurobiol. 2021, 58, 3665–3676. [Google Scholar] [CrossRef]
- Paloczi, J.; Varga, Z.V.; Hasko, G.; Pacher, P. Neuroprotection in oxidative stress-related neurodegenerative diseases: Role of endocannabinoid system modulation. Antioxid. Redox Signal. 2018, 29, 75–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-C.; Xu, Y.-M.; Li, H.-Y.; Wu, C.-Y.; Xu, T.-T.; Luo, N.-C.; Zhang, S.-J.; Wang, Q.; Quan, S.-J. Jiao-Tai-Wan improves cognitive dysfunctions through cholinergic pathway in scopolamine-treated mice. Biomed. Res. Int. 2018, 2018, 3538763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rigi, A.; Heidarian, E.; Amini, S.A. Protective and anti-inflammatory effects of hydroalcoholic leaf extract of Origanum vulgare on oxidative stress, TNF-α gene expression and liver histological changes in paraquat-induced hepatotoxicity in rats. Arch. Physiol. Biochem. 2019, 125, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Wang, J.; Peng, J.; Wei, H. Oregano Essential Oil Induces SOD1 and GSH Expression through Nrf2 activation and alleviates hydrogen peroxide-induced oxidative damage in IPEC-J2 Cells. Oxid. Med. Cell. Longev. 2016, 2016, 5987183. [Google Scholar] [CrossRef]
- Sun, Q.-F.; Chen, S.-X.; Tang, Z.-F.; Song, X.-Y.; Jing, F.; Wu, H.-T.; Ding, Z.-Y.; El-kott, A.; Massoud, D.; Khalifa, H. Origanum vulgare L. leaf extract alleviates finasteride-induced oxidative stress in mouse liver and kidney. Asian Pac. J. Trop. Biomed. 2021, 11, 194. [Google Scholar] [CrossRef]
- Namiecińska, E.; Sadowska, B.; Więckowska-Szakiel, M.; Dołęga, A.; Pasternak, B.; Grazul, M.; Budzisz, E. Anticancer and antimicrobial properties of novel η6-p-cymene ruthenium(II) complexes containing a N,S-type ligand, their structural and theoretical characterization. RSC Adv. 2019, 9, 38629–38645. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.N.; Redhwi, H.H.; Tsagkalias, I.; Vouvoudi, E.C.; Achilias, D.S. Development of bio-composites with enhanced antioxidant activity based on poly(lactic acid) with thymol, carvacrol, limonene, or cinnamaldehyde for active food packaging. Polymers 2021, 13, 3652. [Google Scholar] [CrossRef]
- Formiga, R.d.O.; Júnior, E.B.A.; Vasconcelos, R.C.; Guerra, G.C.B.; de Araújo, A.A.; de Carvalho, T.G.; Garcia, V.B.; Junior, R.F.d.A.; Gadelha, F.A.A.F.; Vieira, G.C.; et al. P-cymene and rosmarinic acid ameliorate tnbs-induced intestinal inflammation upkeeping zo-1 and muc-2: Role of antioxidant system and immunomodulation. Int. J. Mol. Sci. 2020, 21, 5870. [Google Scholar] [CrossRef]
- Memari-Tabrizi, E.F.; Yousefpour-Dokhanieh, A.; Babashpour-Asl, M. Foliar-applied silicon nanoparticles mitigate cadmium stress through physio-chemical changes to improve growth, antioxidant capacity, and essential oil profile of summer savory (Satureja hortensis L.). Plant Physiol. Biochem. 2021, 165, 71–79. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Mumivand, H.; Craker, L.E.; Maggi, F. Chemical composition and antioxidant activity of essential oils in Origanum vulgare subsp. gracile at different phenological stages and plant parts. J. Food Process. Preserv. 2018, 42, e13516. [Google Scholar] [CrossRef]
- Qneibi, M.; Jaradat, N.; Hawash, M.; Zaid, A.N.; Natsheh, A.R.; Yousef, R.; AbuHasan, Q. The neuroprotective role of Origanum syriacum L. and Lavandula dentata L. essential oils through their effects on AMPA receptors. Biomed. Res. Int. 2019, 2019, 5640173. [Google Scholar] [CrossRef] [Green Version]
- Napoli, E.M.; Curcuruto, G.; Ruberto, G. Screening the essential oil composition of wild Sicilian oregano. Biochem. Syst. Ecol. 2009, 37, 484–493. [Google Scholar] [CrossRef]
- NIST Standard Reference Database 1A. Available online: https://www.nist.gov/system/files/documents/srd/NIST1aVer22Man.pdf (accessed on 10 January 2020).
- Sparkman, O.D. Identification of essential oil components by gas chromatography/quadrupole mass spectroscopy Robert P. Adams. J. Am. Soc. Mass Spectrom. 2005, 16, 1902–1903. [Google Scholar] [CrossRef] [Green Version]
- Franscescon, F.; Souza, T.P.; Müller, T.E.; Michelotti, P.; Canzian, J.; Stefanello, F.V.; Rosemberg, D.B. Taurine prevents MK-801-induced shoal dispersion and altered cortisol responses in zebrafish. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 111, 110399. [Google Scholar] [CrossRef]
- Dos Santos, A.C.; Junior, G.B.; Zago, D.C.; Zeppenfeld, C.C.; da Silva, D.T.; Heinzmann, B.M.; Baldisserotto, B.; da Cunha, M.A. Anesthesia and anesthetic action mechanism of essential oils of Aloysia triphylla and Cymbopogon flexuosus in silver catfish (Rhamdia quelen). Vet. Anaesth. Analg. 2017, 44, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Cachat, J.M.; Canavello, P.R.; Elkhayat, S.I.; Bartels, B.K.; Hart, P.C.; Elegante, M.F.; Beeson, E.C.; Laffoon, A.L.; Haymore, W.A.M.; Tien, D.H.; et al. Video-aided analysis of zebrafish locomotion and anxiety-related behavioral responses. Neuromethods 2011, 51, 1–14. [Google Scholar] [CrossRef]
- Rosemberg, D.B.; Rico, E.P.; Mussulini, B.H.M.; Piato, Â.L.; Calcagnotto, M.E.; Bonan, C.D.; Dias, R.D.; Blaser, R.E.; Souza, D.O.; de Oliveira, D.L. Differences in spatio-temporal behavior of zebrafish in the open tank paradigm after a short-period confinement into dark and bright environments. PLoS ONE 2011, 6, e19397. [Google Scholar] [CrossRef] [Green Version]
- De Cognato, G.P.; Bortolotto, J.W.; Blazina, A.R.; Christoff, R.R.; Lara, D.R.; Vianna, M.R.; Bonan, C.D. Y-Maze memory task in zebrafish (Danio rerio): The role of glutamatergic and cholinergic systems on the acquisition and consolidation periods. Neurobiol. Learn. Mem. 2012, 98, 321–328. [Google Scholar] [CrossRef]
- Brinza, I.; Ayoub, I.M.; Eldahshan, O.A.; Hritcu, L. Baicalein 5,6-dimethyl ether prevents memory deficits in the scopolamine zebrafish model by regulating cholinergic and antioxidant systems. Plants 2021, 10, 1245. [Google Scholar] [CrossRef]
- Kielkopf, C.L.; Bauer, W.; Urbatsch, I.L. Bradford assay for determining protein concentration. Cold Spring Harb. Protoc. 2020, 2020, prot102269. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.J.; Featherstone, R.M.; Feather-Stone, R. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # a | KI b | KI c | Class/Compound | % d |
|---|---|---|---|---|
| Monoterpene Hydrocarbons | 49.68 | |||
| 1 | 925 | 930 | α-Thujene | 1.52 (± 0.00) |
| 2 | 933 | 939 | α-Pinene | 0.84 (± 0.00) |
| 3 | 944 | 960 | Thuja-2,4(10)-diene | 0.02 (± 0.00) |
| 4 | 949 | 954 | Camphene | 0.10 (± 0.00) |
| 5 | 973 | 968 | Verbenene | 0.01 (± 0.00) |
| 6 | 976 | 979 | β-Pinene | 0.14 (± 0.03) |
| 9 | 988 | 991 | β-Myrcene | 2.09 (± 0.00) |
| 10 | 1002 | 1003 | α-Phellandrene | 0.34 (± 0.00) |
| 11 | 1009 | 1004 | p-Menth-1(7),8-diene | 0.11 (± 0.00) |
| 12 | 1016 | 1017 | α-Terpinene | 3.51 (± 0.01) |
| 13 | 1027 | 1025 | p-Cymene | 20.28 (± 0.03) |
| 14 | 1030 | 1029 | Limonene | 0.67 (± 0.06) |
| 16 | 1037 | 1037 | cis-β-Ocimene | 0.18 (± 0.00) |
| 17 | 1048 | 1050 | trans-β-Ocimene | 0.09 (± 0.00) |
| 18 | 1062 | 1060 | γ-Terpinene | 19.58 (± 0.08) |
| 20 | 1088 | 1089 | Terpinolene | 0.20 (± 0.00) |
| Oxygenated Monoterpenes | 44.93 | |||
| 15 | 1033 | 1031 | 1,8-Cineol | 0.03 (± 0.00) |
| 19 | 1069 | 1070 | cis-Sabinene hydrate | 0.17 (± 0.00) |
| 22 | 1099 | 1098 | trans-Sabinene hydrate | 0.07 (± 0.00) |
| 23 | 1169 | 1169 | Borneol | 0.11 (± 0.01) |
| 24 | 1179 | 1177 | Terpinen-4-ol | 0.80 (± 0.00) |
| 25 | 1187 | 1183 | p-Cymen-8-ol | 0.02 (± 0.00) |
| 26 | 1196 | 1189 | α-Terpineol | 0.09 (± 0.00) |
| 27 | 1203 | 1201 | trans-Dihydro Carvone | 0.01 (± 0.00) |
| 28 | 1218 | 1215 | cis-Dihydro Carvone | 0.03 (± 0.00) |
| 29 | 1237 | 1235 | Thymol methyl ether | 0.92 (± 0.00) |
| 30 | 1248 | 1245 | Carvacrol methyl ether | 3.11 (± 0.00) |
| 31 | 1260 | 1243 | Carvone | 0.03 (± 0.00) |
| 32 | 1287 | 1289 | Bornyl acetate | 0.02 (± 0.00) |
| 33 | 1292 | 1291 | p-Cymen-7-ol | 0.03 (± 0.00) |
| 34 | 1306 | 1290 | Thymol | 38.82 (± 0.05) |
| 35 | 1311 | 1299 | Carvacrol | 0.59 (± 0.00) |
| 36 | 1358 | 1252 | Thymol acetate | 0.08 (± 0.00) |
| Sesquiterpenes | 3.94 | |||
| 37 | 1377 | 1375 | α-Ylangene | 0.03 (± 0.00) |
| 38 | 1382 | 1377 | α-Copaene | 0.07 (± 0.00) |
| 39 | 1390 | 1388 | β-Bourbonene | 0.05 (± 0.00) |
| 40 | 1426 | 1419 | β-Caryophyllene | 0.97 (± 0.00) |
| 41 | 1435 | 1432 | β-Copaene | 0.08 (± 0.00) |
| 42 | 1440 | 1435 | β-Bergamotene | 0.03 (± 0.00) |
| 43 | 1460 | 1455 | α-Humulene | 0.11 (± 0.00) |
| 44 | 1481 | 1480 | γ-Muurolene | 0.27 (± 0.00) |
| 45 | 1485 | 1485 | α-Amorphene | 0.03 (± 0.00) |
| 46 | 1492 | 1490 | β-Selinene | 0.02 (± 0.00) |
| 47 | 1498 | 1496 | γ-Amorphene | 0.10 (± 0.00) |
| 48 | 1504 | 1500 | α-Muurolene | 0.08 (± 0.00) |
| 49 | 1512 | 1506 | β-Bisabolene | 1.27 (± 0.01) |
| 50 | 1520 | 1514 | γ-Cadinene | 0.22 (± 0.00) |
| 51 | 1529 | 1523 | δ-Cadinene | 0.48 (± 0.00) |
| 52 | 1544 | 1539 | α-Cadinene | 0.02 (± 0.00) |
| 53 | 1549 | 1546 | α-Calacorene | 0.01 (± 0.00) |
| 54 | 1590 | 1583 | Caryophyllene oxide | 0.10 (± 0.00) |
| Others | 0.41 | |||
| 7 | 978 | 979 | 1-Octen-3-ol | 0.33 (± 0.03) |
| 8 | 984 | 984 | 3-Octanone | 0.07 (± 0.00) |
| 21 | 1094 | 1091 | Methyl benzoate | 0.01 (± 0.00) |
| Total | 98.96 | |||
| Monoterpene hydrocarbons | 49.68 | |||
| Oxygenated monoterpenes | 44.93 | |||
| Sesquiterpenes | 3.94 | |||
| Others | 0.41 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capatina, L.; Napoli, E.M.; Ruberto, G.; Hritcu, L. Origanum vulgare ssp. hirtum (Lamiaceae) Essential Oil Prevents Behavioral and Oxidative Stress Changes in the Scopolamine Zebrafish Model. Molecules 2021, 26, 7085. https://doi.org/10.3390/molecules26237085
Capatina L, Napoli EM, Ruberto G, Hritcu L. Origanum vulgare ssp. hirtum (Lamiaceae) Essential Oil Prevents Behavioral and Oxidative Stress Changes in the Scopolamine Zebrafish Model. Molecules. 2021; 26(23):7085. https://doi.org/10.3390/molecules26237085
Chicago/Turabian StyleCapatina, Luminita, Edoardo Marco Napoli, Giuseppe Ruberto, and Lucian Hritcu. 2021. "Origanum vulgare ssp. hirtum (Lamiaceae) Essential Oil Prevents Behavioral and Oxidative Stress Changes in the Scopolamine Zebrafish Model" Molecules 26, no. 23: 7085. https://doi.org/10.3390/molecules26237085
APA StyleCapatina, L., Napoli, E. M., Ruberto, G., & Hritcu, L. (2021). Origanum vulgare ssp. hirtum (Lamiaceae) Essential Oil Prevents Behavioral and Oxidative Stress Changes in the Scopolamine Zebrafish Model. Molecules, 26(23), 7085. https://doi.org/10.3390/molecules26237085