
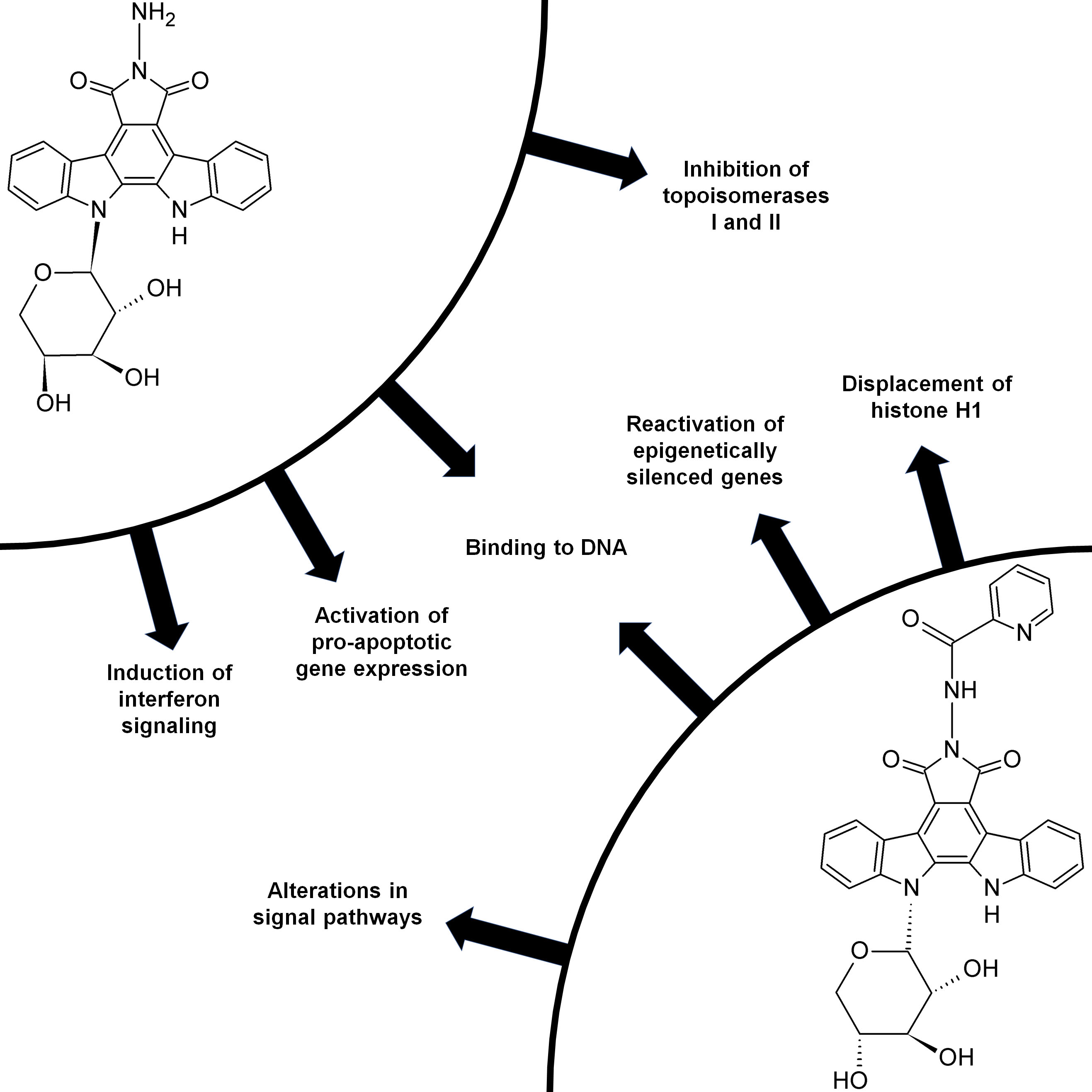
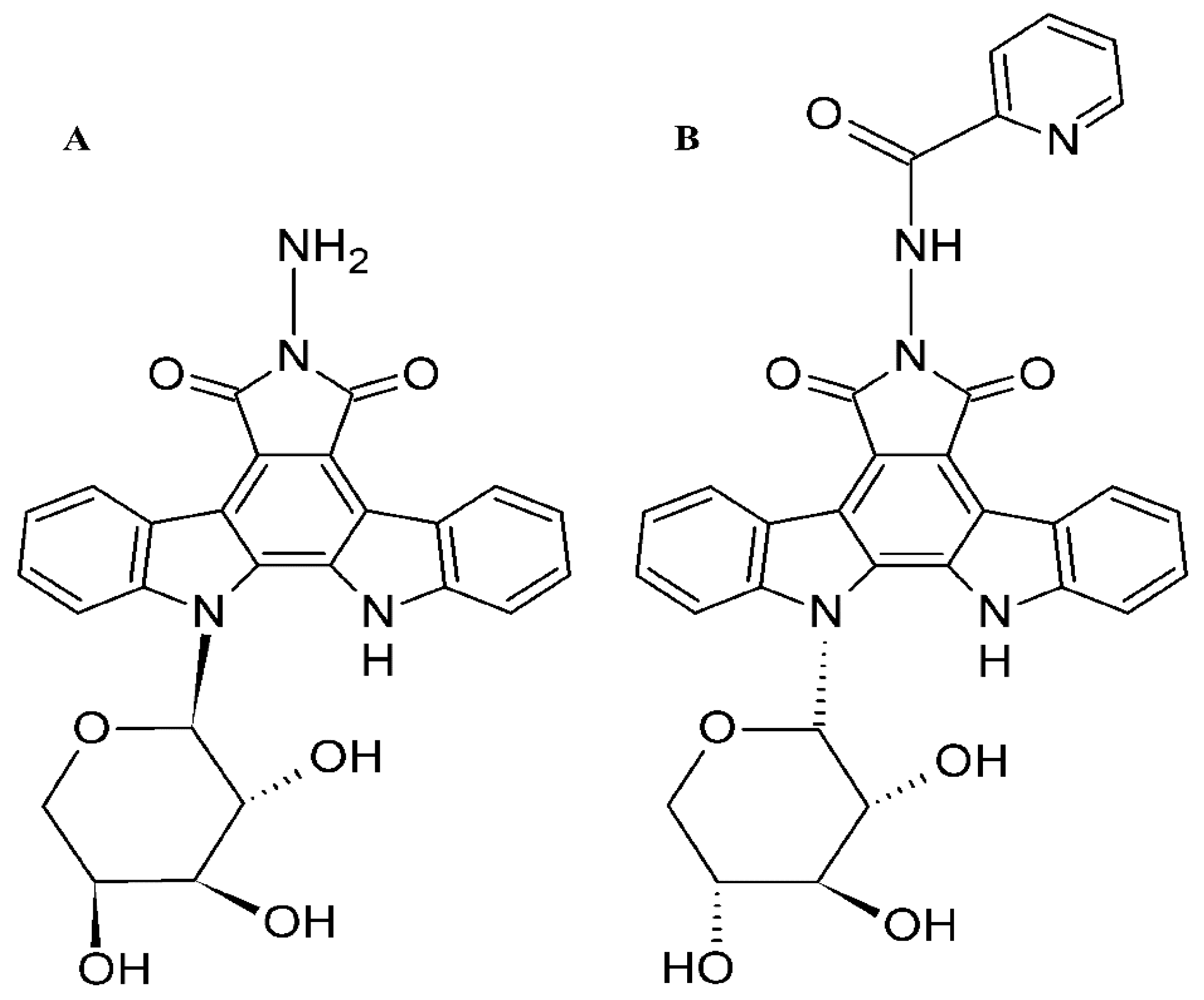
Molecular Mechanisms of Anticancer Activity of N-Glycosides of Indolocarbazoles LCS-1208 and LCS-1269

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Cytotoxicity Assay
2.3. Treatment, RNA Isolation, cDNA Preparation, and qRT-PCR
2.4. Circular Dichroism in Cholesteric Liquid Crystals
2.5. Fluorescent Intercalator Displacement Assay (FID)
2.6. Topoisomerase I Activity Assay
2.7. Topoisomerase II Activity Assay
2.8. Live Cell Microscopy
2.9. Flow Cytometry
2.10. Treatment, Histone Extraction and Western Blot
2.11. Treatment, Nuclear Extraction and Western Blot
2.12. Treatment, DNA-Isolation, DNA Methylation ELISA and Methylation-Sensitive HpaII/MspI Restriction Enzyme Assay (MSRE)
2.13. qRT-PCR for Interferon Signaling and Gene Expression of Histone Methyltransferases, Histone Deacetylases and DNA Methyltransferases
2.14. Statistical Methods
3. Results
3.1. Cytotoxicity of LCS-1208 and LCS-1269 in Human Cancer Cell Lines
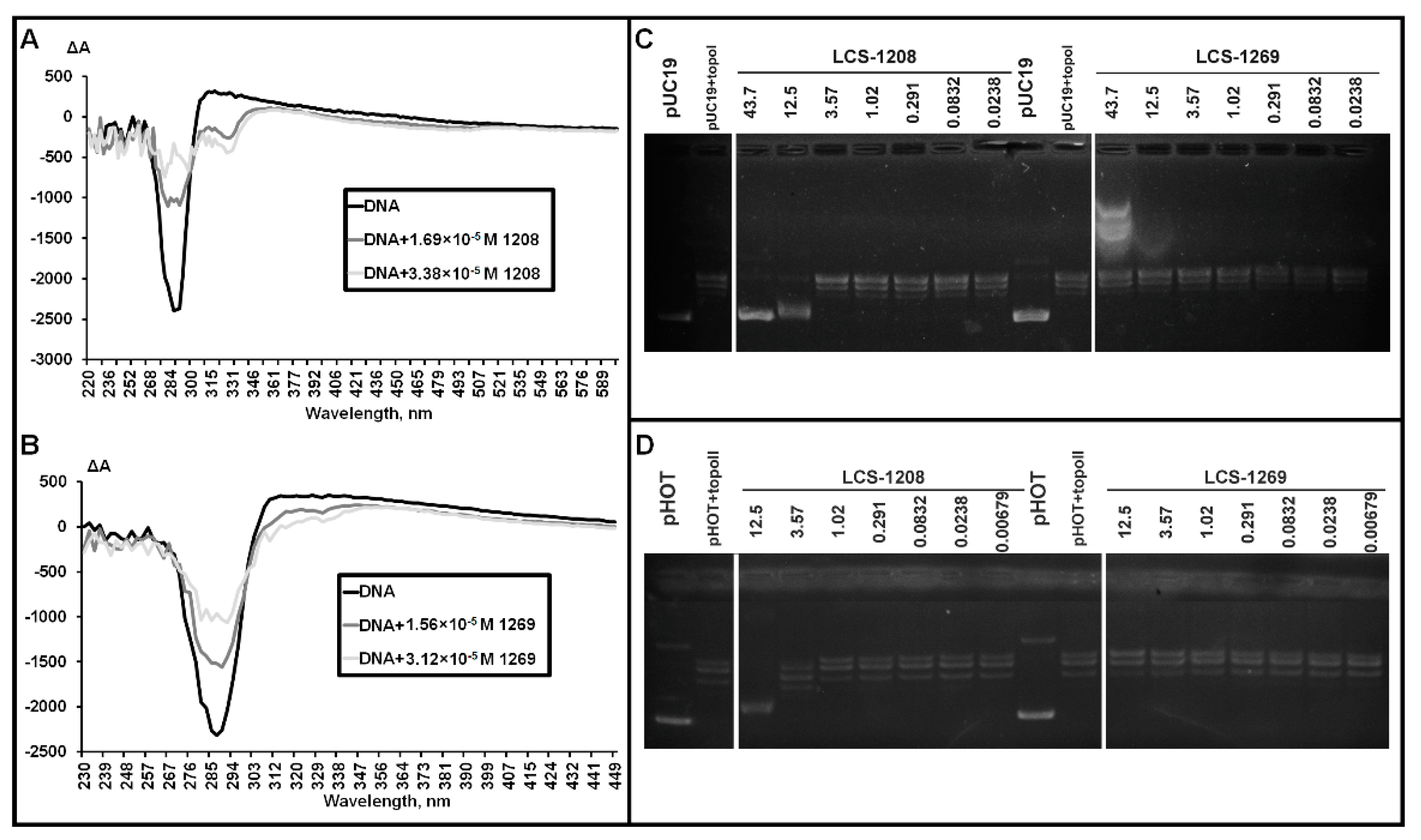
3.2. Affinity of LCS-1208 and LCS-1269 to DNA Duplex
3.3. Analysis of LCS-DNA Complexes by Means of Circular Dichroism in Cholesteric Liquid Crystals
3.4. Topoisomerase I Assay
3.5. Topoisomerase IIα Assay
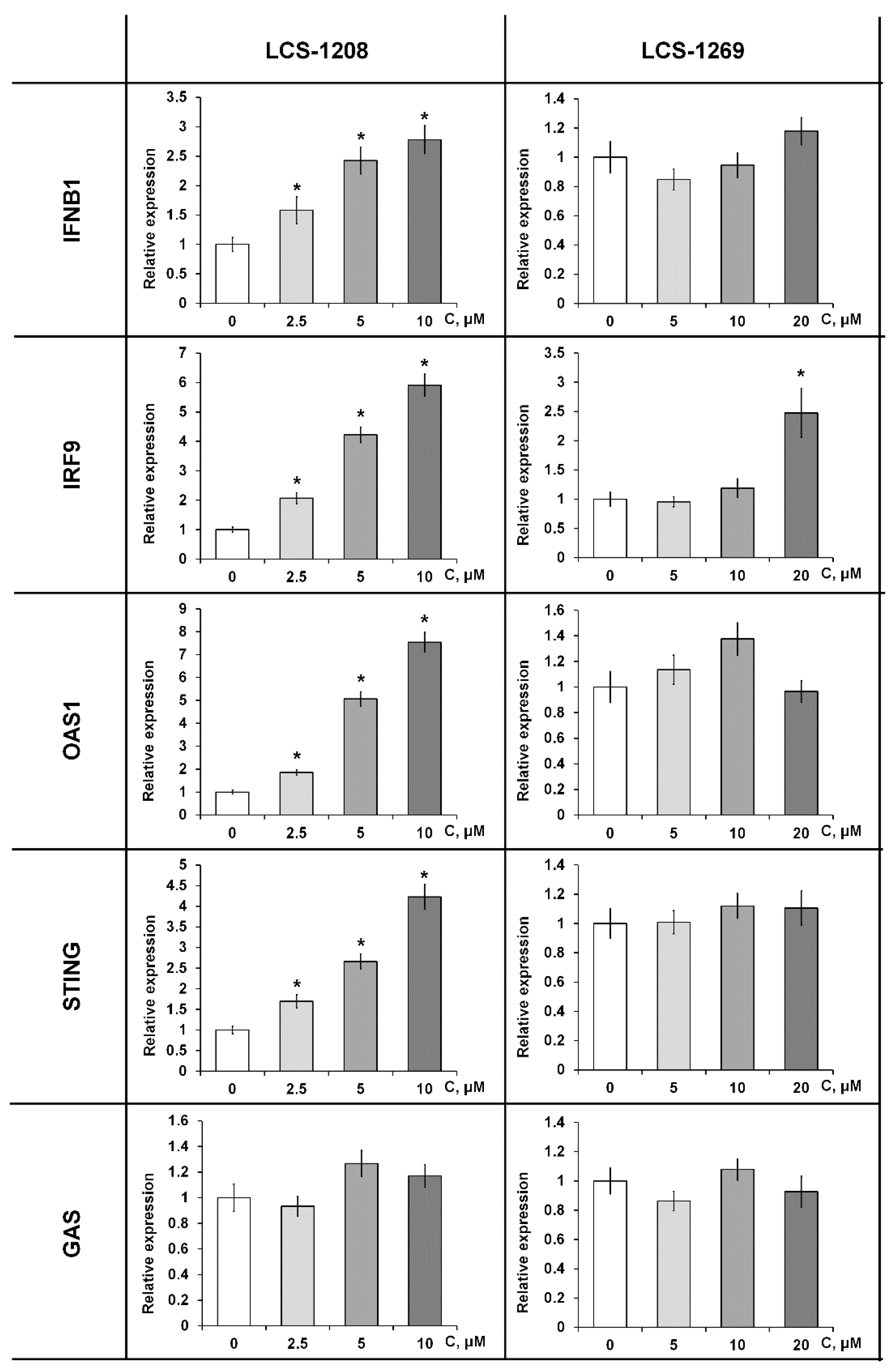
3.6. The Effect of LCS-1208 and LCS-1269 on Interferon Signaling
3.7. Chromatin Remodeling
3.7.1. Linker Histone H1 Displacement
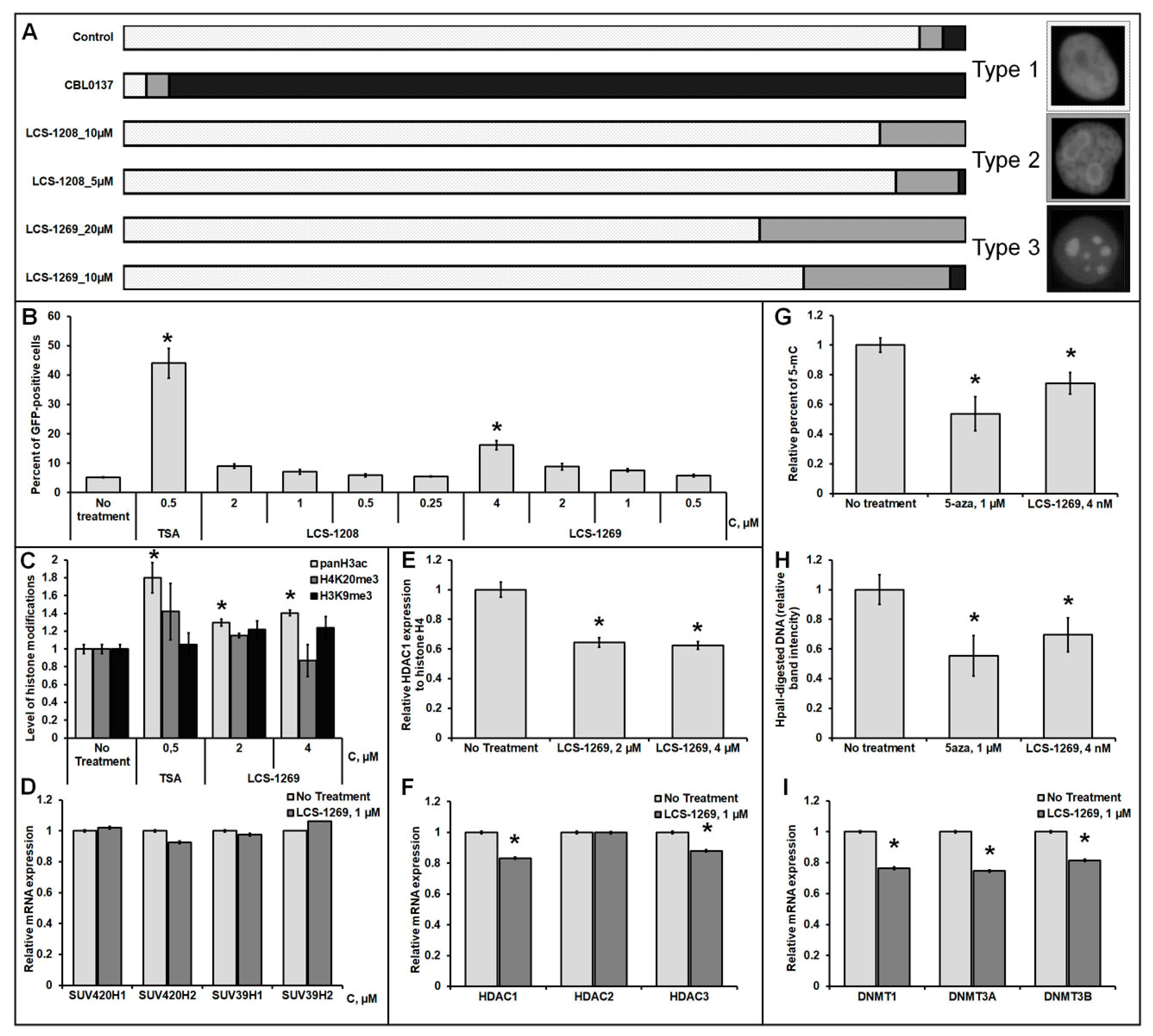
3.7.2. Analysis of the LCS-1208 and LCS-1269 Ability to Reactivate the Expression of the Epigenetically Repressed GFP Gene in HeLa TI Cell-Based Assay
3.7.3. Analysis of LCS-1269 Effect on Histone Modifications
3.7.4. Analysis of LCS-1269 Effect on DNA Methylation
3.8. The Analysis of LCS-Induced Expression Changes in a Gene Set of Neoplastic Transformation
3.9. The Analysis of LCS-Induced Expression Changes in a Set of Target Genes of Signal Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Janosik, T.; Rannug, A.; Rannug, U.; Wahlström, N.; Slätt, J.; Bergman, J. Chemistry and Properties of Indolocarbazoles. Chem. Rev. 2018, 118, 9058–9128. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.; Méndez, C.; Salas, J.A. Indolocarbazole natural products: Occurrence, biosynthesis, and biological activity. Nat. Prod. Rep. 2006, 23, 1007–1045. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.W.; Reddy, K.R.; Knölker, H.J. Occurrence, biogenesis, and synthesis of biologically active carbazole alkaloids. Chem. Rev. 2012, 112, 3193–3328. [Google Scholar] [CrossRef] [PubMed]
- Zenkov, R.G.; Ektova, L.V.; Vlasova, O.A.; Belitskiy, G.A.; Yakubovskaya, M.G.; Kirsanov, K.I. Indolo[2,3-a]carbazoles: Diversity, biological properties, application in antitumor therapy. Chem. Heterocycl. Compd. 2020, 56, 644–658. [Google Scholar] [CrossRef]
- Bailly, C.; Riou, J.-F.; Colson, P.; Houssier, C.; Rodrigues-Pereira, E.; Prudhomme, M. DNA Cleavage by Topoisomerase I in the Presence of Indolocarbazole Derivatives of Rebeccamycin. Biochemistry 1997, 36, 3917–3929. [Google Scholar] [CrossRef]
- Kojiri, K.; Kondo, H.; Yoshinari, T.; Arakawa, H.; Nakajima, S.; Satoh, F.; Kawamura, K.; Okura, A.; Suda, H.; Okanishi, M. A new antitumor substance BE-13793C, produced by a streptomycete. Taxonomy, fermentation, isolation, structure determination and biological activity. J. Antibiot. 1991, 44, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Ling-Hua, M.; Zhi-Yong, L.; Pommier, Y. Non-camptothecin DNA topoisomerase I inhibitors in cancer therapy. Curr. Top. Med. Chem. 2003, 3, 305–320. [Google Scholar] [CrossRef]
- Long, B.H.; Rose, W.C.; Vyas, D.M.; Matson, J.A.; Forenza, S. Discovery of antitumor indolocarbazoles: Rebeccamycin, NSC 655649, and fluoroindolocarbazoles. Curr. Med. Chem. Anticancer Agents 2002, 2, 255–266. [Google Scholar] [CrossRef]
- Moreau, P.; Anizon, F.; Sancelme, M.; Prudhomme, M.; Sevère, D.; Riou, J.-F.; Goossens, J.-F.; Hénichart, J.-P.; Bailly, C.; Labourier, E.; et al. Synthesis, Mode of Action, and Biological Activities of Rebeccamycin Bromo Derivatives. J. Med. Chem. 1999, 42, 1816–1822. [Google Scholar] [CrossRef] [PubMed]
- Moreau, P.; Gaillard, N.; Marminon, C.; Anizon, F.; Dias, N.; Baldeyrou, B.; Bailly, C.; Pierré, A.; Hickman, J.; Pfeiffer, B.; et al. Semi-synthesis, topoisomerase I and kinases inhibitory properties, and antiproliferative activities of new rebeccamycin derivatives. Bioorg. Med. Chem. 2003, 11, 4871–4879. [Google Scholar] [CrossRef]
- Prudhomme, M. Rebeccamycin analogues as anti-cancer agents. Eur. J. Med. Chem. 2003, 38, 123–140. [Google Scholar] [CrossRef]
- Bailly, C.; Qu, X.; Anizon, F.; Prudhomme, M.; Riou, J.-F.; Chaires, J.B. Enhanced Binding to DNA and Topoisomerase I Inhibition by an Analog of the Antitumor Antibiotic Rebeccamycin Containing an Amino Sugar Residue. Mol. Pharmacol. 1999, 55, 377. [Google Scholar] [CrossRef] [Green Version]
- Staker, B.L.; Feese, M.D.; Cushman, M.; Pommier, Y.; Zembower, D.; Stewart, L.; Burgin, A.B. Structures of Three Classes of Anticancer Agents Bound to the Human Topoisomerase I−DNA Covalent Complex. J. Med. Chem. 2005, 48, 2336–2345. [Google Scholar] [CrossRef]
- Civenni, G.; Longoni, N.; Costales, P.; Dallavalle, C.; García Inclán, C.; Albino, D.; Nuñez, L.E.; Morís, F.; Carbone, G.M.; Catapano, C.V. EC-70124, a Novel Glycosylated Indolocarbazole Multikinase Inhibitor, Reverts Tumorigenic and Stem Cell Properties in Prostate Cancer by Inhibiting STAT3 and NF-κB. Mol. Cancer Ther. 2016, 15, 806–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathi, A.T.; Levis, M. Lestaurtinib: A multi-targeted FLT3 inhibitor. Expert Rev. Hematol. 2009, 2, 17–26. [Google Scholar] [CrossRef]
- Garcia, J.S.; Percival, M.E. Midostaurin for the treatment of adult patients with newly diagnosed acute myeloid leukemia that is FLT3 mutation-positive. Drugs Today 2017, 53, 531–543. [Google Scholar] [CrossRef]
- Lawrie, A.M.; Noble, M.E.; Tunnah, P.; Brown, N.R.; Johnson, L.N.; Endicott, J.A. Protein kinase inhibition by staurosporine revealed in details of the molecular interaction with CDK2. Nat. Struct. Biol. 1997, 4, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Maroney, A.C.; Finn, J.P.; Connors, T.J.; Durkin, J.T.; Angeles, T.; Gessner, G.; Xu, Z.; Meyer, S.L.; Savage, M.J.; Greene, L.A.; et al. Cep-1347 (KT7515), a semisynthetic inhibitor of the mixed lineage kinase family. J. Biol. Chem. 2001, 276, 25302–25308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meggio, F.; Donella Deana, A.; Ruzzene, M.; Brunati, A.M.; Cesaro, L.; Guerra, B.; Meyer, T.; Mett, H.; Fabbro, D.; Furet, P.; et al. Different susceptibility of protein kinases to staurosporine inhibition. Kinetic studies and molecular bases for the resistance of protein kinase CK2. Eur. J. Biochem. 1995, 234, 317–322. [Google Scholar] [CrossRef]
- Omura, S.; Sasaki, Y.; Iwai, Y.; Takeshima, H. Staurosporine, a potentially important gift from a microorganism. J. Antibiot. 1995, 48, 535–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Fujita, N.; Tsuruo, T. Interference with PDK1-Akt survival signaling pathway by UCN-01 (7-hydroxystaurosporine). Oncogene 2002, 21, 1727–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senderowicz, A.M. The cell cycle as a target for cancer therapy: Basic and clinical findings with the small molecule inhibitors flavopiridol and UCN-01. Oncologist 2002, 7, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Strock, C.J.; Park, J.-I.; Rosen, M.; Dionne, C.; Ruggeri, B.; Jones-Bolin, S.; Denmeade, S.R.; Ball, D.W.; Nelkin, B.D. CEP-701 and CEP-751 Inhibit Constitutively Activated RET Tyrosine Kinase Activity and Block Medullary Thyroid Carcinoma Cell Growth. Cancer Res. 2003, 63, 5559. [Google Scholar] [PubMed]
- Osada, H.; Koshino, H.; Kudo, T.; Onose, R.; Isono, K. A new inhibitor of protein kinase C, RK-1409 (7-oxostaurosporine). I. Taxonomy and biological activity. J. Antibiot. 1992, 45, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Rüegg, U.T.; Burgess, G.M. Staurosporine, K-252 and UCN-01: Potent but nonspecific inhibitors of protein kinases. Trends Pharmacol. Sci. 1989, 10, 218–220. [Google Scholar] [CrossRef]
- Komander, D.; Kular, G.S.; Bain, J.; Elliott, M.; Alessi, D.R.; Van Aalten, D.M. Structural basis for UCN-01 (7-hydroxystaurosporine) specificity and PDK1 (3-phosphoinositide-dependent protein kinase-1) inhibition. Biochem. J. 2003, 375, 255–262. [Google Scholar] [CrossRef]
- Rodrigues Pereira, E.; Belin, L.; Sancelme, M.; Prudhomme, M.; Ollier, M.; Rapp, M.; Sevère, D.; Riou, J.-F.; Fabbro, D.; Meyer, T. Structure−Activity Relationships in a Series of Substituted Indolocarbazoles: Topoisomerase I and Protein Kinase C Inhibition and Antitumoral and Antimicrobial Properties. J. Med. Chem. 1996, 39, 4471–4477. [Google Scholar] [CrossRef] [PubMed]
- Speck, K.; Magauer, T. The chemistry of isoindole natural products. Beilstein J. Org. Chem. 2013, 9, 2048–2078. [Google Scholar] [CrossRef] [PubMed]
- Conseil, G.; Perez-Victoria, J.M.; Jault, J.-M.; Gamarro, F.; Goffeau, A.; Hofmann, J.; Di Pietro, A. Protein Kinase C Effectors Bind to Multidrug ABC Transporters and Inhibit Their Activity. Biochemistry 2001, 40, 2564–2571. [Google Scholar] [CrossRef]
- Robey, R.W.; Shukla, S.; Steadman, K.; Obrzut, T.; Finley, E.M.; Ambudkar, S.V.; Bates, S.E. Inhibition of ABCG2-mediated transport by protein kinase inhibitors with a bisindolylmaleimide or indolocarbazole structure. Mol. Cancer Ther. 2007, 6, 1877. [Google Scholar] [CrossRef] [Green Version]
- Kiseleva, M.P.; Smirnova, Z.S.; Borisova, L.M.; Kubasova, I.Y.; Ektova, L.V.; Miniker, T.D.; Plikhtiak, I.L.; Medvedeva, L.A.; Eremina, V.A.; Tikhonova, N.I. Search for new antitumor compounds among N-glycoside indolo[2,3-a]carbazole derivatives. Ross. Onkol. Zhurnal 2015, 20, 33–37. (In Russian) [Google Scholar]
- Golubeva, I.S.; Eremina, V.A.; Moiseeva, N.I.; Ektova, L.V.; Yavorskaya, N.P. Derivative of the Class of n-glycosides indolo[2,3-a]pyrrolo[3,4-c]carbazole-5,7-dione-n-{12-(β-d-xylopyranosyl)-5,7-dioxo-indolo[2,3-a]pyrrolo[3,4-c]carbazol-6-yl} pyridine-2-carboxamide, Which has Cytotoxic and Antitumor Activity. Russia Patent 2667906c1, 21 March 2017. [Google Scholar]
- Luthra, P.; Aguirre, S.; Yen, B.C.; Pietzsch, C.A.; Sanchez-Aparicio, M.T.; Tigabu, B.; Morlock, L.K.; García-Sastre, A.; Leung, D.W.; Williams, N.S.; et al. Topoisomerase II Inhibitors Induce DNA Damage-Dependent Interferon Responses Circumventing Ebola Virus Immune Evasion. mBio 2017, 8, e00368-17. [Google Scholar] [CrossRef] [Green Version]
- Pépin, G.; Nejad, C.; Thomas, B.J.; Ferrand, J.; McArthur, K.; Bardin, P.G.; Williams, B.R.; Gantier, M.P. Activation of cGAS-dependent antiviral responses by DNA intercalating agents. Nucleic Acids Res. 2017, 45, 198–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharjee, S.; Chakraborty, S.; Sengupta, P.K.; Bhowmik, S. Exploring the Interactions of the Dietary Plant Flavonoids Fisetin and Naringenin with G-Quadruplex and Duplex DNA, Showing Contrasting Binding Behavior: Spectroscopic and Molecular Modeling Approaches. J. Phys. Chem. B 2016, 120, 8942–8952. [Google Scholar] [CrossRef] [PubMed]
- Shechter, D.; Dormann, H.L.; Allis, C.D.; Hake, S.B. Extraction, purification and analysis of histones. Nat. Protoc. 2007, 2, 1445–1457. [Google Scholar] [CrossRef]
- Monchaud, D.; Allain, C.; Teulade-Fichou, M.P. Development of a fluorescent intercalator displacement assay (G4-FID) for establishing quadruplex-DNA affinity and selectivity of putative ligands. Bioorg. Med. Chem. Lett. 2006, 16, 4842–4845. [Google Scholar] [CrossRef] [PubMed]
- Nordén, B. Applications of linear Dichroism Spectroscopy. Appl. Spectrosc. Rev. 1978, 14, 157–248. [Google Scholar] [CrossRef]
- Yevdokimov, Y.M.; Skuridin, S.G.; Lortkipanidze, G.B. Invited Article Liquid-crystalline dispersions of nucleic acids. Liq. Cryst. 1992, 12, 1–16. [Google Scholar] [CrossRef]
- Yevdokimov, Y.M.; Skuridin, S.G.; Nechipurenko, Y.D.; Zakharov, M.A.; Salyanov, V.I.; Kurnosov, A.A.; Kuznetsov, V.D.; Nikiforov, V.N. Nanoconstructions based on double-stranded nucleic acids. Int. J. Biol. Macromol. 2005, 36, 103–115. [Google Scholar] [CrossRef]
- Poleshko, A.; Einarson, M.B.; Shalginskikh, N.; Zhang, R.; Adams, P.D.; Skalka, A.M.; Katz, R.A. Identification of a functional network of human epigenetic silencing factors. J. Biol. Chem. 2010, 285, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Maksimova, V.; Shalginskikh, N.; Vlasova, O.; Usalka, O.; Beizer, A.; Bugaeva, P.; Fedorov, D.; Lizogub, O.; Lesovaya, E.; Katz, R.; et al. HeLa TI cell-based assay as a new approach to screen for chemicals able to reactivate the expression of epigenetically silenced genes. PLoS ONE 2021, 16, e0252504. [Google Scholar] [CrossRef] [PubMed]
- Kiseleva, M.P.; Shprakh, Z.S.; Borisova, L.M.; Kubasova, I.Y.; Dezhenkova, L.G.; Kaluzhny, D.N.; Lantsova, A.V.; Sanarova, E.V.; Shtil, A.A.; Oborotova, N.A.; et al. Preclinical study of antitumor activity of indolocarbazoles N-glycosides derivative LCS-1208. Report II. Russ. J. Biother. 2015, 14, 41–48. (In Russian) [Google Scholar] [CrossRef]
- Strekowski, L.; Wilson, B. Noncovalent interactions with DNA: An overview. Mutat. Res. 2007, 623, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Härtlova, A.; Erttmann, S.F.; Raffi, F.A.; Schmalz, A.M.; Resch, U.; Anugula, S.; Lienenklaus, S.; Nilsson, L.M.; Kröger, A.; Nilsson, J.A.; et al. DNA damage primes the type I interferon system via the cytosolic DNA sensor STING to promote anti-microbial innate immunity. Immunity 2015, 42, 332–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paludan, S.R.; Bowie, A.G. Immune sensing of DNA. Immunity 2013, 38, 870–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Käs, E.; Izaurralde, E.; Laemmli, U.K. Specific inhibition of DNA binding to nuclear scaffolds and histone H1 by distamycin. The role of oligo(dA).oligo(dT) tracts. J. Mol. Biol. 1989, 210, 587–599. [Google Scholar] [CrossRef]
- Pang, B.; Qiao, X.; Janssen, L.; Velds, A.; Groothuis, T.; Kerkhoven, R.; Nieuwland, M.; Ovaa, H.; Rottenberg, S.; van Tellingen, O.; et al. Drug-induced histone eviction from open chromatin contributes to the chemotherapeutic effects of doxorubicin. Nat. Commun. 2013, 4, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Sasikala, W.D. Drug-DNA intercalation: From discovery to the molecular mechanism. Adv. Protein Chem. Struct. Biol. 2013, 92, 1–62. [Google Scholar] [CrossRef]
- Leonova, K.; Safina, A.; Nesher, E.; Sandlesh, P.; Pratt, R.; Burkhart, C.; Lipchick, B.; Gitlin, I.; Frangou, C.; Koman, I.; et al. TRAIN (Transcription of Repeats Activates INterferon) in response to chromatin destabilization induced by small molecules in mammalian cells. eLife 2018, 7, e30842. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.H.; Goutte-Gattat, D.; Becker, N.; Meyer, S.; Shukla, M.S.; Hayes, J.J.; Everaers, R.; Angelov, D.; Bednar, J.; Dimitrov, S. Single-base resolution mapping of H1-nucleosome interactions and 3D organization of the nucleosome. Proc. Natl. Acad. Sci. USA 2010, 107, 9620–9625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb. Perspect. Med. 2016, 6, a026831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klutstein, M.; Nejman, D.; Greenfield, R.; Cedar, H. DNA Methylation in Cancer and Aging. Cancer Res. 2016, 76, 3446–3450. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.K.; Majumder, H.K.; Roychoudhury, S. Natural Compounds as Anticancer Agents Targeting DNA Topoisomerases. Curr. Genomics 2017, 18, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Chen, Y. Lymphangiogenesis and colorectal cancer. Saudi Med. J. 2017, 38, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Wiesmann, M.; Rohan, M.; Chan, V.; Jefferson, A.B.; Guo, L.; Sakamoto, D.; Caothien, R.H.; Fuller, J.H.; Reinhard, C.; et al. Elevated expression of axin2 and hnkd mRNA provides evidence that Wnt/beta -catenin signaling is activated in human colon tumors. Proc. Natl. Acad. Sci. USA 2001, 98, 14973–14978. [Google Scholar] [CrossRef] [Green Version]
- Schatoff, E.M.; Leach, B.I.; Dow, L.E. Wnt Signaling and Colorectal Cancer. Curr. Colorectal Cancer Rep. 2017, 13, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | |
|---|---|---|
| RPL27 | 5′-ACCGCTACCCCCGCAAAGTG-3′ | 5′-CCCGTCGGGCCTTGCGTTTA-3′ |
| IFNB1 | 5′-ACGCCGCATTGACCATCTAT-3′ | 5′-GTCTCATTCCAGCCAGTGCT-3′ |
| IRF9 | 5′-CAACTGAGGCCCCCTTTCAA-3′ | 5′-CGCCCGTTGTAGATGAAGGT-3′ |
| OAS1 | 5′-TGGAGACCCAAAGGGTTGGA-3′ | 5′-AGGAAGCAGGAGGTCTCACC-3′ |
| GAS | 5′-ACGTGCTGTGAAAACAAAGAAG-3′ | 5′-GTCCCACTGACTGTCTTGAGG-3′ |
| STING | 5′-ATATCTGCGGCTGATCCTGC-3′ | 5′-GGTCTGCTGGGGCAGTTTAT-3′ |
| HDAC1 | 5′-CACCCATTCTTCCCGTTCTT-3′ | 5′-GGCATTTCAGGAGTTTGTCTTAT-3′ |
| HDAC2 | 5′-CTTATTGTGTGTCTGCCCATTT-3′ | 5′-ATTTGTCTGCTTCCTGCTACT-3′ |
| HDAC3 | 5′-AATGCCTTCAACGTAGGCGA-3′ | 5′-GGGTTGCTCCTTGCAGAGAT-3′ |
| DNMT1 | 5′-AGCACAGAAGTCAACCCAAA-3′ | 5′-TGCGTCTCTTCTCCTCCTTT-3′ |
| DNMT3A | 5′-AGCCCAAGGTCAAGGAGATT-3′ | 5′-TACGCACACTCCAGAAAGC-3′ |
| DNMT3B | 5′-CAACAGCATCGGCAGGAA-3′ | 5′-GTCCTCTGTGTCGTCTGTGA-3′ |
| SUV420H1 | 5′-CCCGTGTAGCATAAAAGCAGC-3′ | 5′-CCAGTTTCACCAAGGAACCAG-3′ |
| SUV420H2 | 5′-CGTGCTTGGAAGAAGAATGA-3′ | 5′-GCAGTCATGGTTGATGAAGG-3′ |
| SUV39H1 | 5′-GCTAGGCCCGAATGTCGTTA-3′ | 5′-TAGAGATACCGAGGGCAGGG-3′ |
| SUV39H2 | 5′-GCAGGACGAACTCAACAGAA-3′ | 5′-CAACCAAAGGTGGCTTCATT-3′ |
| LCS-1269 | LCS-1208 | Histogenesis/Cancer Type | |
|---|---|---|---|
| Adhesion lines | |||
| HeLa | 26.6 ± 2.1 | 28.1 ± 1.8 | Cervical adenocarcinoma |
| MCF-7 | 31 ± 1 | 5.5 ± 0.7 | Breast adenocarcinoma |
| HepG2 | 2.5 ± 0.5 | 1.7 ± 0.3 | Hepatocellular carcinoma |
| U251 | 1.2 ± 0.06 | 0.36 ± 0.08 | Glioblastoma |
| A549 | 3.2 ± 0.3 | 1.0 ± 0.01 | Lung adenocarcinoma |
| PC-3 | 24 ± 4 | 0.97 ± 0.01 | Prostatic adenocarcinoma |
| HT29 | 1.4 ± 0.5 | 0.13 ± 0.01 | Colorectal adenocarcinoma |
| HaCaT | 29 ± 2 | 31 ± 3 | Spontaneously immortalized keratinocytes |
| Suspension lines | |||
| CCRF CEM | 6.8 ± 0.7 | 2.0 ± 1.2 | Acute lymphoblastic leukemia |
| K562 | >500 | 6.0 ± 0.06 | Chronic myelogenous leukemia |
| KG-1 | 7.1 ± 2.3 | 0.6 ± 0.2 | Acute myelogenous leukemia |
| Granta-519 | 0.60 ± 0.02 | 0.071 ± 0.008 | B-cell lymphoma |
| LCS-1208 | LCS-1269 | |||
|---|---|---|---|---|
| Interaction with DNA | Mechanism of interaction (CD in CLC) | Intercalation | Intercalation or groove binding | |
| Binding constant (FID) | 1.24 × 106 M−1 | 2.52 × 105 M−1 | ||
| Decrease of DNA electrophoretic mobility (Electrophoresis) | + | - | ||
| Inhibition of topoisomerase I | + | - | ||
| Inhibition of topoisomerase IIα | + | - | ||
| Activation of interferon signaling | + | - | ||
| Displacement of histone H1 | - | + | ||
| Influence on the following mechanisms of epigenetic regulation of gene expression: | Expression of epigenetically repressed GFP gene (HeLa TI cell-based assay) | - | Reactivation | |
| DNA methylation | Decrease | |||
| DNA methyltransferase gene expression | Decrease | |||
| Histone modifications associated with actively transcribed chromatin and repressed transcription | Increase in H3 acetylation (actively transcribed chromatin) | |||
| Expression of genes of histone deacetylases | Decrease in HDAC1 and HDAC3 expression | |||
| Expression of genes encoding histone methyltransferases | - | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zenkov, R.G.; Vlasova, O.A.; Maksimova, V.P.; Fetisov, T.I.; Karpechenko, N.Y.; Ektova, L.V.; Eremina, V.A.; Popova, V.G.; Usalka, O.G.; Lesovaya, E.A.; et al. Molecular Mechanisms of Anticancer Activity of N-Glycosides of Indolocarbazoles LCS-1208 and LCS-1269. Molecules 2021, 26, 7329. https://doi.org/10.3390/molecules26237329
Zenkov RG, Vlasova OA, Maksimova VP, Fetisov TI, Karpechenko NY, Ektova LV, Eremina VA, Popova VG, Usalka OG, Lesovaya EA, et al. Molecular Mechanisms of Anticancer Activity of N-Glycosides of Indolocarbazoles LCS-1208 and LCS-1269. Molecules. 2021; 26(23):7329. https://doi.org/10.3390/molecules26237329
Chicago/Turabian StyleZenkov, Roman G., Olga A. Vlasova, Varvara P. Maksimova, Timur I. Fetisov, Natalia Y. Karpechenko, Lidiya V. Ektova, Vera A. Eremina, Valeriia G. Popova, Olga G. Usalka, Ekaterina A. Lesovaya, and et al. 2021. "Molecular Mechanisms of Anticancer Activity of N-Glycosides of Indolocarbazoles LCS-1208 and LCS-1269" Molecules 26, no. 23: 7329. https://doi.org/10.3390/molecules26237329
APA StyleZenkov, R. G., Vlasova, O. A., Maksimova, V. P., Fetisov, T. I., Karpechenko, N. Y., Ektova, L. V., Eremina, V. A., Popova, V. G., Usalka, O. G., Lesovaya, E. A., Belitsky, G. A., Yakubovskaya, M. G., & Kirsanov, K. I. (2021). Molecular Mechanisms of Anticancer Activity of N-Glycosides of Indolocarbazoles LCS-1208 and LCS-1269. Molecules, 26(23), 7329. https://doi.org/10.3390/molecules26237329