
An Analysis of Variability in the Content of Phenolic Acids and Flavonoids in Camelina Seeds Depending on Weather Conditions, Functional Form, and Genotypes

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
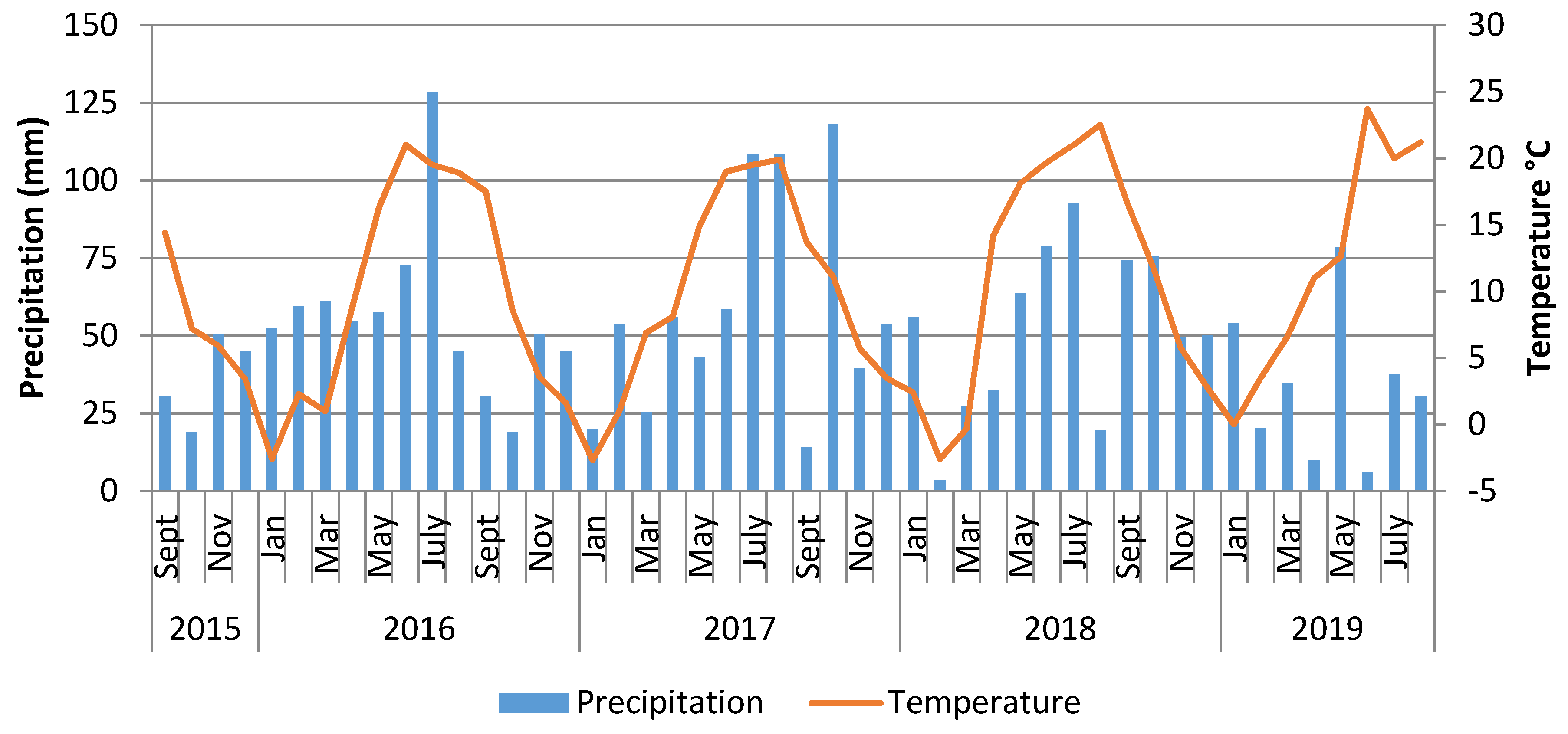
2.1. Weather Conditions
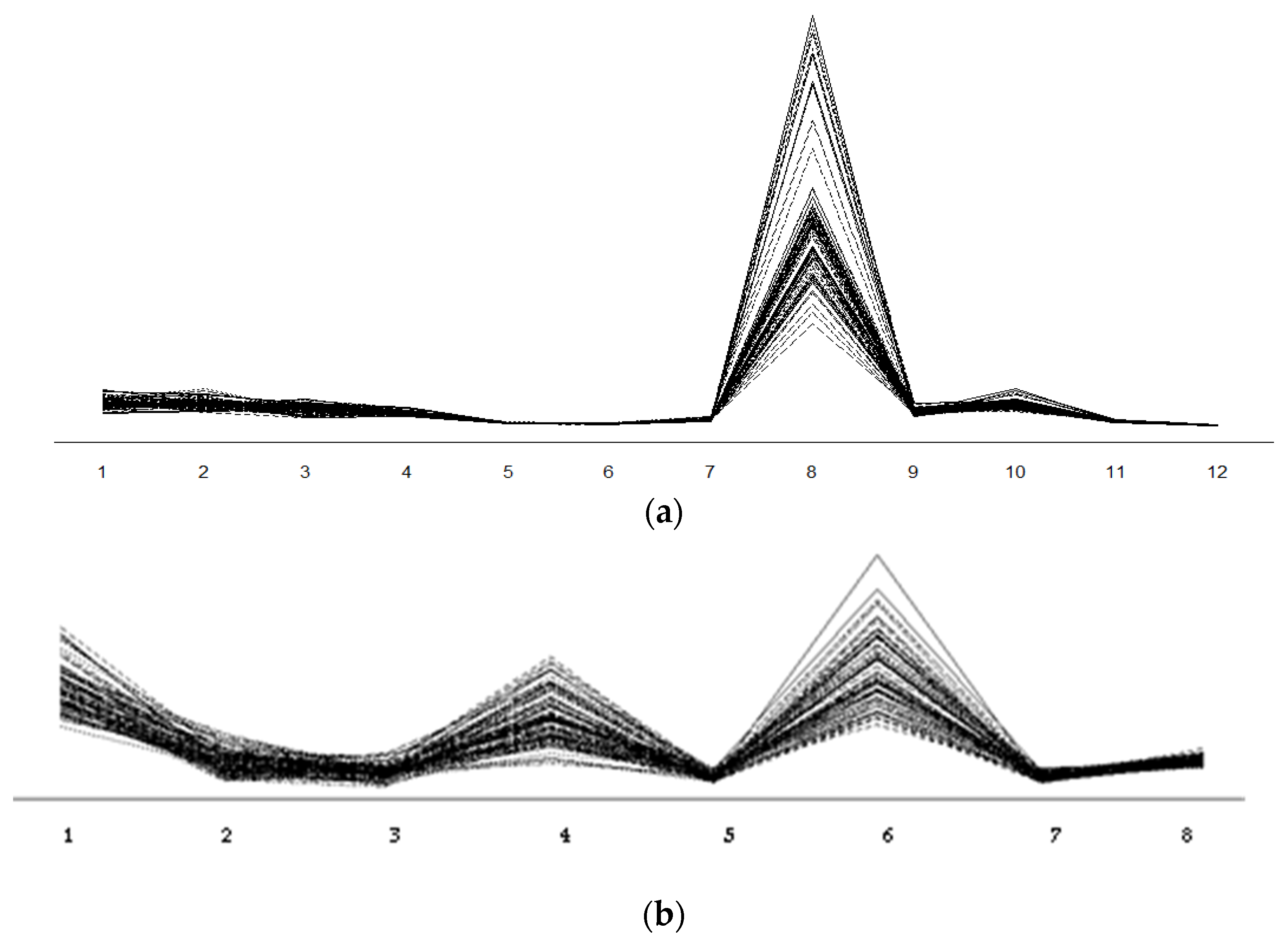
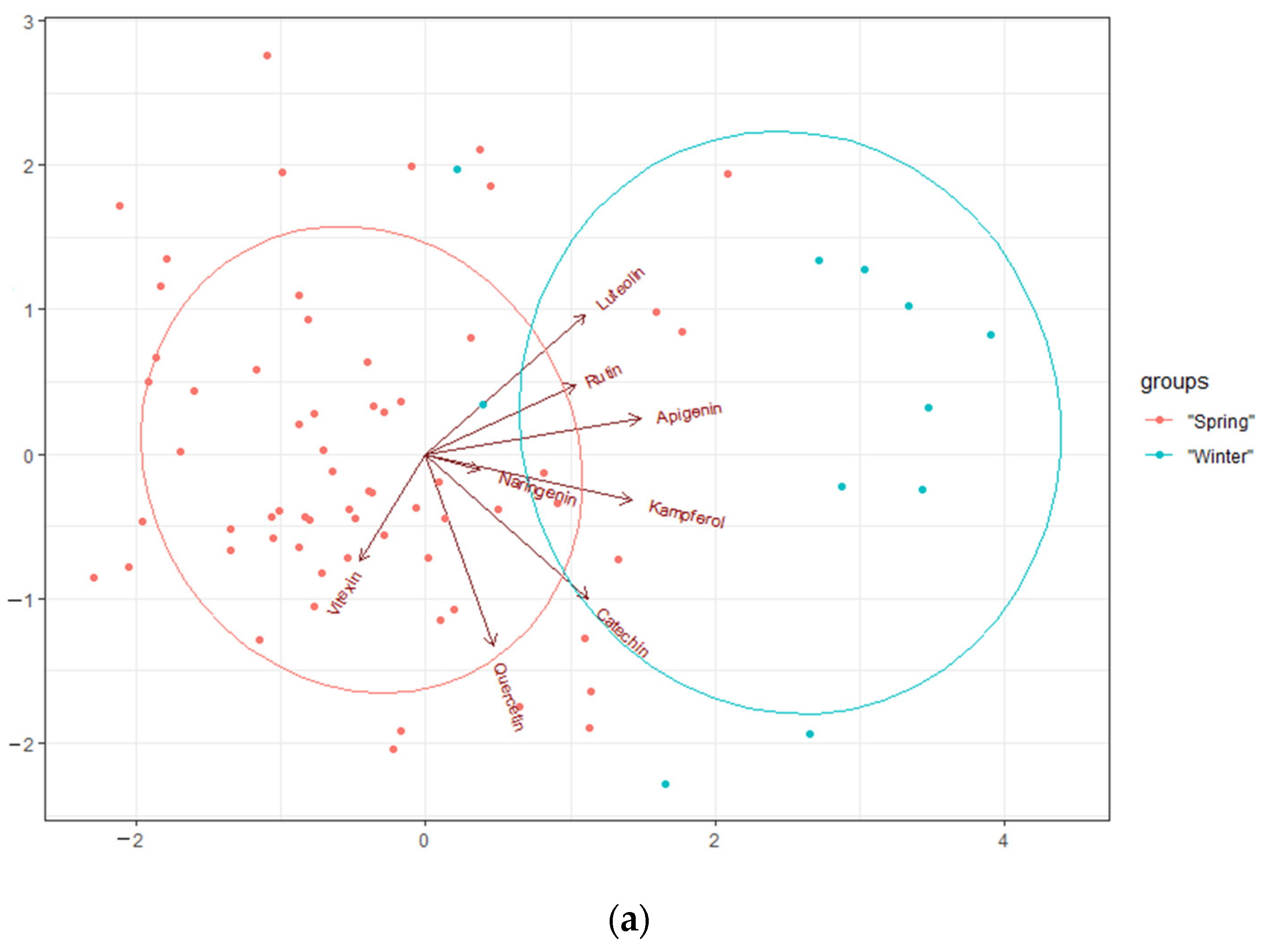
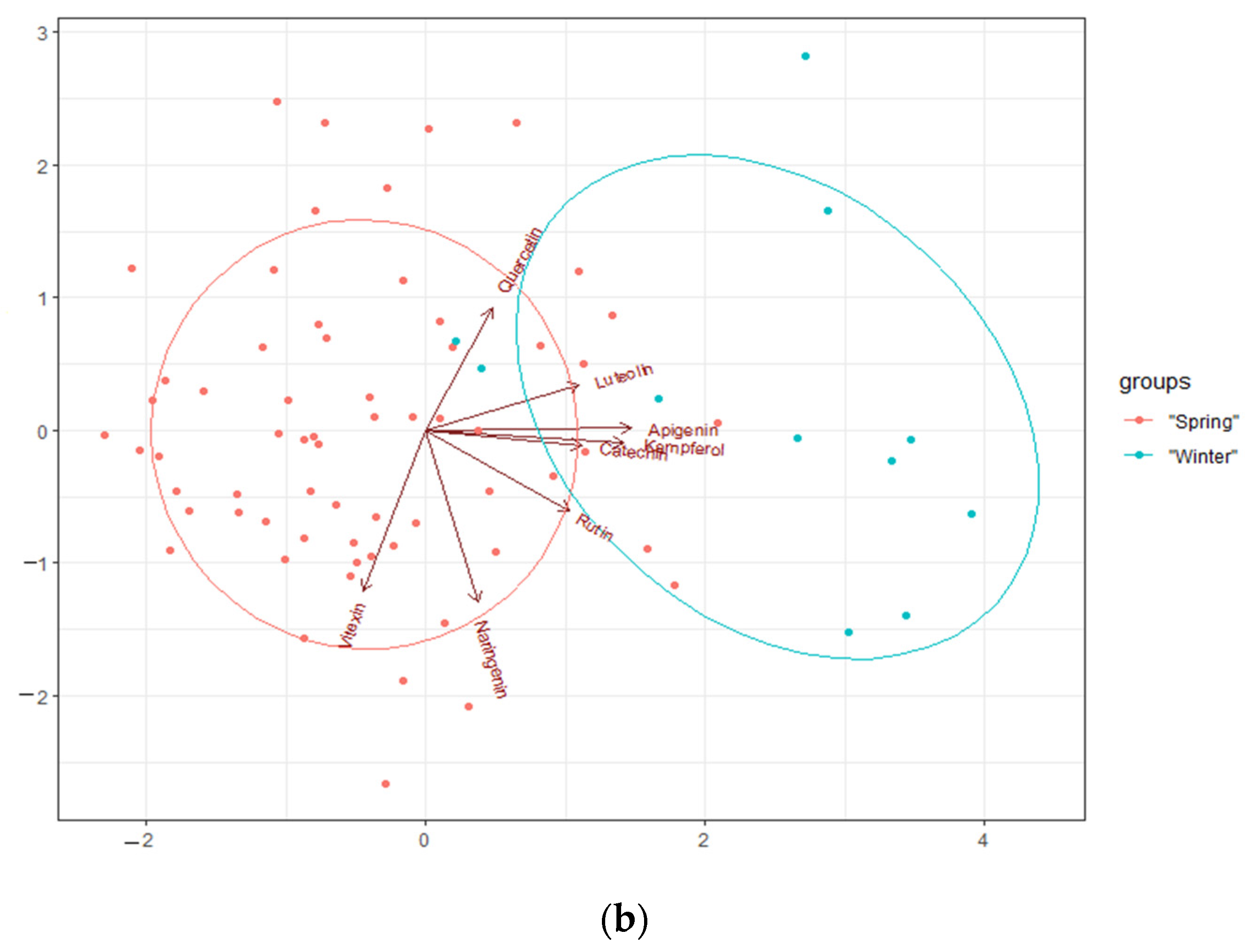
2.2. The Content of Polyphenols
3. Materials and Methods
3.1. Test Material
3.2. Field Experiments
3.3. Analysis of Phenolic Compounds
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Arab-Tehrany, E.; Jacquot, M.; Gaiani, C.; Imran, M.; Desobry, S.; Linder, M. Beneficial effects and oxidative stability of omega-3 long-chain polyunsaturated fatty acids. Trends Food Sci. Technol. 2012, 25, 24–33. [Google Scholar] [CrossRef]
- Grosshagauer, S.; Steinschaden, R.; Pignitter, M. Strategies to increase the oxidative stability of cold pressed oils. LWT 2019, 106, 72–77. [Google Scholar] [CrossRef]
- Choe, E.; Min, D.B. Mechanisms and Factors for Edible Oil Oxidation. Compr. Rev. Food Sci. Food Saf. 2006, 5, 169–186. [Google Scholar] [CrossRef]
- Ben, H.R.; Gargouri, B.; Bouaziz, M. Chemical Changes Occur in Extra-Virgin Olive Oil during Fruits Ripeness of Zalmati Cultivar Planted in Warm Desert Climate. J. Oleo Sci. 2022, 71, 469–479. [Google Scholar]
- Szukalska, E. Wybrane zagadnienia utleniania tłuszczów. Tłuszcze Jadalne 2003, 38, 42–61. [Google Scholar]
- Kurasiak-Popowska, D.; Stuper-Szablewska, K. The phytochemical quality of Camelina sativa seed and oil. Acta Agric. Scand. Sect. B Soil Plant Sci. 2019, 70, 39–47. [Google Scholar] [CrossRef]
- Pathak, R.; Mohsin, M.; Mehta, S.P.S. An Assessment of in vitro Antioxidant Potential of Camelina sativa L. Seed Oil and Estimation of Tocopherol Content using HPTLC Method. Int. J. Sci. Res. 2021, 13, 589–600. [Google Scholar] [CrossRef]
- Abramovi, H.; Butinar, B.; Nikolič, V. Changes occurring in phenolic content, tocopherol composition and oxidative stability of Camelina sativa oil during storage. Food Chem. 2007, 104, 903–909. [Google Scholar] [CrossRef]
- Tavarini, S.; De Leo, M.; Matteo, R.; Lazzeri, L.; Braca, A.; Angelini, L.G. Flaxseed and Camelina Meals as Potential Sources of Health-Beneficial Compounds. Plants 2021, 10, 156. [Google Scholar] [CrossRef]
- West, A.L.; Miles, E.A.; Lillycrop, K.A.; Han, L.; Napier, J.A.; Calder, P.C.; Burdge, G.C. Dietary supplementation with seed oil from transgenic Camelina sativa induces similar increments in plasma and erythrocyte DHA and EPA to fish oil in healthy humans. Br. J. Nutr. 2020, 124, 922–930. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Graczyk, M.; Przybylska-Balcerek, A.; Stuper-Szablewska, K. Influence of variety and weather conditions on fatty acid composition of winter and spring Camelina sativa varieties in Poland. Eur. Food Res. Technol. 2021, 247, 465–473. [Google Scholar] [CrossRef]
- Terpinc, P.; Polak, T.; Makuc, D.; Ulrih, N.P.; Abramovič, H. The occurrence and characterization of phenolic compounds in Camelina sativa seed, cake and oil. Food Chem. 2012, 131, 580–589. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic Compounds in Brassica Vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Cartea, E.; De Haro-Bailón, A.; Padilla, G.; Obregón-Cano, S.; del Rio-Celestino, M.; Ordás, A. Seed Oil Quality of Brassica napus and Brassica rapa Germplasm from Northwestern Spain. Foods 2019, 8, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pék, Z.; Daood, H.; Nagyné, M.; Neményi, A.; Helyes, L.M. Effect of environmental conditions and water status on the bioactive compounds of broccoli. Open Life Sci. 2013, 8, 777–787. [Google Scholar] [CrossRef]
- Vollmann, J.; Eynck, C. Camelina as a sustainable oilseed crop: Contributions of plant breeding and genetic engineering. Biotechnol. J. 2015, 10, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crops Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Henderson, A.E.; Hallett, R.H.; Soroka, J. Prefeeding Behavior of the Crucifer Flea Beetle, Phyllotreta cruciferae, on Host and Nonhost Crucifers. J. Insect. Behav. 2004, 17, 17–39. [Google Scholar] [CrossRef]
- Séguin-Swartz, G.; Eynck, C.; Gugel, R.K.; Strelkov, S.E.; Olivier, C.Y.; Li, J.L.; Klein-Gebbinck, H.; Borhan, H.; Caldwell, C.D.; Falk, K.C. Diseases of Camelina sativa (false flax). Can. J. Plant Pathol. 2009, 31, 375–386. [Google Scholar] [CrossRef]
- Zanetti, F.; Gesch, R.W.; Walia, M.K.; Johnson, J.M.F.; Monti, A. Winter camelina root characteristics and yield performance under contrasting environmental conditions. Field Crops Res. 2020, 252, 107794. [Google Scholar] [CrossRef]
- Zanetti, F.; Eynck, C.; Christou, M.; Krzyżaniak, M.; Righini, D.; Alexopoulou, E.; Stolarski, M.J.; Van Loo, E.N.; Puttick, D.; Monti, A. Agronomic performance and seed quality attributes of Camelina (Camelina sativa L. crantz) in multi-environment trials across Europe and Canada. Ind. Crops Prod. 2017, 107, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.J.; de Camargo, A.C.; Shahidi, F. Phenolic profiles and antioxidant activity of defatted camelina and sophia seeds. Food Chem. 2018, 240, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Bilska, K.; Stuper-Szablewska, K.; Kulik, T.; Buśko, M.; Załuski, D.; Jurczak, S.; Perkowski, J. Changes in Phenylpropanoid and Trichothecene Production by Fusarium culmorum and F. graminearum Sensu Stricto via Exposure to Flavonoids. Toxins 2018, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybylska-Balcerek, A.; Szablewski, T.; Szwajkowska-Michałek, L.; Świerk, D.; Cegielska-Radziejewska, R.; Krejpcio, Z.; Suchowilska, E.; Tomczyk, Ł.; Stuper-Szablewska, K. Sambucus Nigra Extracts–Natural Antioxidants and Antimicrobial Compounds. Molecules 2021, 26, 2910. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lambda-Wilks | R2 | |
|---|---|---|
| phenols | ||
| Sinapic acid | 0.370077 | 0.136062 |
| Protocatechuic acid | 0.144588 | 0.201786 |
| Syringic acid | 0.127974 | 0.070381 |
| Caffeic acid | 0.130240 | 0.070799 |
| 9-hydroxybenzoic acid | 0.128604 | 0.101760 |
| Chlorogenic acid | 0.126685 | 0.311918 |
| Ferulic acid | 0.123810 | 0.159464 |
| flavonoids | ||
| Apigenin | 0.539560 | 0.031118 |
| Kaempferol | 0.572799 | 0.029863 |
| Vitexin | 0.467732 | 0.026047 |
| Rutin | 0.438533 | 0.0211658 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurasiak-Popowska, D.; Graczyk, M.; Przybylska-Balcerek, A.; Stuper-Szablewska, K.; Szwajkowska-Michałek, L. An Analysis of Variability in the Content of Phenolic Acids and Flavonoids in Camelina Seeds Depending on Weather Conditions, Functional Form, and Genotypes. Molecules 2022, 27, 3364. https://doi.org/10.3390/molecules27113364
Kurasiak-Popowska D, Graczyk M, Przybylska-Balcerek A, Stuper-Szablewska K, Szwajkowska-Michałek L. An Analysis of Variability in the Content of Phenolic Acids and Flavonoids in Camelina Seeds Depending on Weather Conditions, Functional Form, and Genotypes. Molecules. 2022; 27(11):3364. https://doi.org/10.3390/molecules27113364
Chicago/Turabian StyleKurasiak-Popowska, Danuta, Małgorzata Graczyk, Anna Przybylska-Balcerek, Kinga Stuper-Szablewska, and Lidia Szwajkowska-Michałek. 2022. "An Analysis of Variability in the Content of Phenolic Acids and Flavonoids in Camelina Seeds Depending on Weather Conditions, Functional Form, and Genotypes" Molecules 27, no. 11: 3364. https://doi.org/10.3390/molecules27113364
APA StyleKurasiak-Popowska, D., Graczyk, M., Przybylska-Balcerek, A., Stuper-Szablewska, K., & Szwajkowska-Michałek, L. (2022). An Analysis of Variability in the Content of Phenolic Acids and Flavonoids in Camelina Seeds Depending on Weather Conditions, Functional Form, and Genotypes. Molecules, 27(11), 3364. https://doi.org/10.3390/molecules27113364