
Identification of New Regulators of Pancreatic Cancer Cell Sensitivity to Oxaliplatin and Cisplatin



Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. sgRNA Libraries
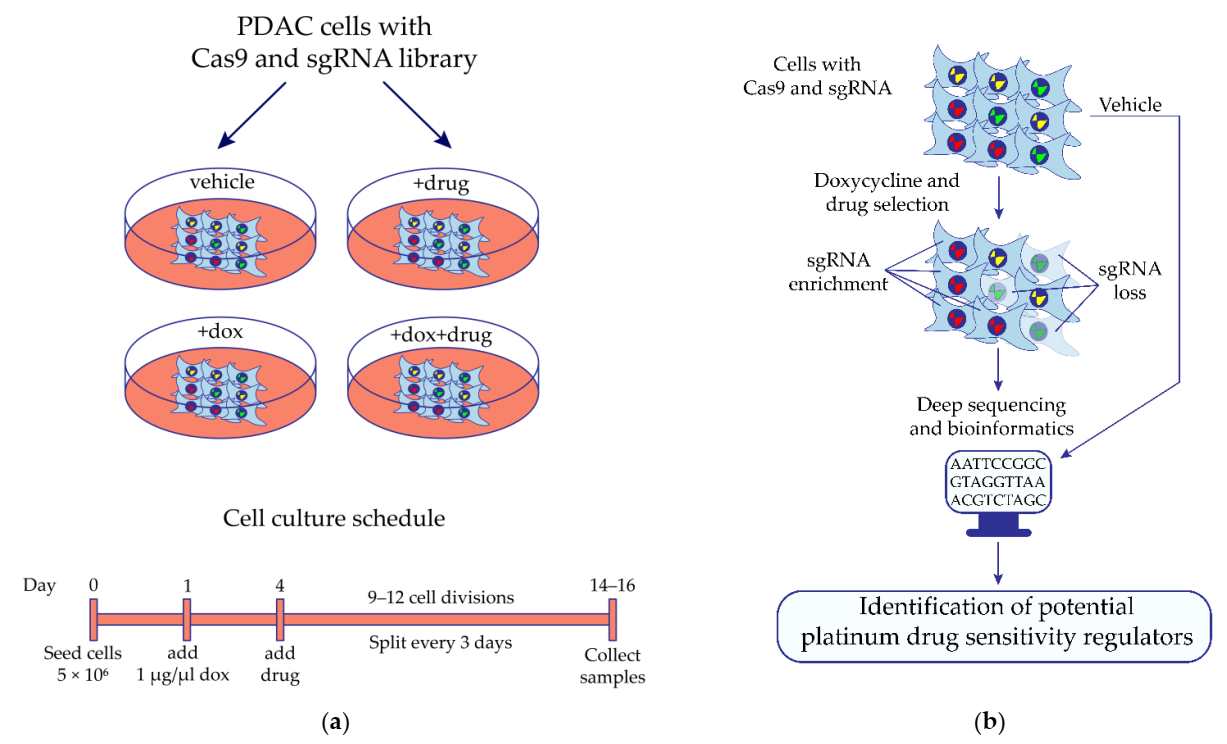
4.3. In Vitro CRISPR/Cas9 Screening
4.4. Gene Ontology Enrichment Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sarantis, P.; Koustas, E.; Papadimitropoulou, A.; Papavassiliou, A.G.; Karamouzis, M.V. Pancreatic Ductal Adenocarcinoma: Treatment Hurdles, Tumor Microenvironment and Immunotherapy. World J. Gastrointest. Oncol. 2020, 12, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer Treatment and Survivorship Statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting Cancer Incidence and Deaths to 2030: The Unexpected Burden of Thyroid, Liver, and Pancreas Cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducreux, M.; Seufferlein, T.; Van Laethem, J.-L.; Laurent-Puig, P.; Smolenschi, C.; Malka, D.; Boige, V.; Hollebecque, A.; Conroy, T. Systemic Treatment of Pancreatic Cancer Revisited. Semin. Oncol. 2019, 46, 28–38. [Google Scholar] [CrossRef]
- Ghaneh, P.; Costello, E.; Neoptolemos, J.P. Biology and Management of Pancreatic Cancer. Gut 2007, 56, 1134–1152. [Google Scholar] [CrossRef]
- Singh, R.R.; O’Reilly, E.M. New Treatment Strategies for Metastatic Pancreatic Ductal Adenocarcinoma. Drugs 2020, 80, 647–669. [Google Scholar] [CrossRef]
- Singh, R.R.; Goldberg, J.; Varghese, A.M.; Yu, K.H.; Park, W.; O’Reilly, E.M. Genomic Profiling in Pancreatic Ductal Adenocarcinoma and a Pathway towards Therapy Individualization: A Scoping Review. Cancer Treat. Rev. 2019, 75, 27–38. [Google Scholar] [CrossRef]
- Kamel, D.; Gray, C.; Walia, J.S.; Kumar, V. PARP Inhibitor Drugs in the Treatment of Breast, Ovarian, Prostate and Pancreatic Cancers: An Update of Clinical Trials. Curr. Drug. Targets 2018, 19, 21–37. [Google Scholar] [CrossRef]
- Ortíz, R.; Quiñonero, F.; García-Pinel, B.; Fuel, M.; Mesas, C.; Cabeza, L.; Melguizo, C.; Prados, J. Nanomedicine to Overcome Multidrug Resistance Mechanisms in Colon and Pancreatic Cancer: Recent Progress. Cancers 2021, 13, 2058. [Google Scholar] [CrossRef]
- Friedenson, B. BRCA1 and BRCA2 Pathways and the Risk of Cancers Other than Breast or Ovarian. Medscape Gen. Med. 2005, 7, 60. [Google Scholar]
- Lynch, H.T.; Deters, C.A.; Snyder, C.L.; Lynch, J.F.; Villeneuve, P.; Silberstein, J.; Martin, H.; Narod, S.A.; Brand, R.E. BRCA1 and Pancreatic Cancer: Pedigree Findings and Their Causal Relationships. Cancer Genet. Cytogenet. 2005, 158, 119–125. [Google Scholar] [CrossRef]
- Liu, Q.; Liao, Q.; Zhao, Y. Chemotherapy and Tumor Microenvironment of Pancreatic Cancer. Cancer Cell Int. 2017, 17, 68. [Google Scholar] [CrossRef] [Green Version]
- Dauer, P.; Nomura, A.; Saluja, A.; Banerjee, S. Microenvironment in Determining Chemo-Resistance in Pancreatic Cancer: Neighborhood Matters. Pancreatology 2017, 17, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Quiñonero, F.; Mesas, C.; Doello, K.; Cabeza, L.; Perazzoli, G.; Jimenez-Luna, C.; Rama, A.R.; Melguizo, C.; Prados, J. The Challenge of Drug Resistance in Pancreatic Ductal Adenocarcinoma: A Current Overview. Cancer Biol. Med. 2019, 16, 688–699. [Google Scholar] [CrossRef]
- Moffat, G.T.; O’Reilly, E.M. The Role of PARP Inhibitors in Germline BRCA-Associated Pancreatic Ductal Adenocarcinoma. Clin. Adv. Hematol. Oncol. 2020, 18, 168–179. [Google Scholar]
- Rebelatto, T.F.; Falavigna, M.; Pozzari, M.; Spada, F.; Cella, C.A.; Laffi, A.; Pellicori, S.; Fazio, N. Should Platinum-Based Chemotherapy Be Preferred for Germline BReast CAncer Genes (BRCA) 1 and 2-Mutated Pancreatic Ductal Adenocarcinoma (PDAC) Patients? A Systematic Review and Meta-Analysis. Cancer Treat. Rev. 2019, 80, 101895. [Google Scholar] [CrossRef]
- Platinum Response Characteristics of Patients with Pancreatic Ductal Adenocarcinoma and a Germline BRCA1, BRCA2 or PALB2 Mutation. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7000723/ (accessed on 23 December 2021).
- Kiyamova, R.; Garifulin, O.; Gryshkova, V.; Kostianets, O.; Shyian, M.; Gout, I.; Filonenko, V. Preliminary Study of Thyroid and Colon Cancers-Associated Antigens and Their Cognate Autoantibodies as Potential Cancer Biomarkers. Biomarkers 2012, 17, 362–371. [Google Scholar] [CrossRef]
- Kostianets, O.; Shyian, M.; Sergiy, D.; Antoniuk, S.; Gout, I.; Filonenko, V.; Kiyamova, R. Serological Analysis of SEREX-Defined Medullary Breast Carcinoma-Associated Antigens. Cancer Investig. 2012, 30, 519–527. [Google Scholar] [CrossRef]
- Kostianets, O.; Shyyan, M.; Antoniuk, S.V.; Filonenko, V.; Kiyamova, R. Panel of SEREX-Defined Antigens for Breast Cancer Autoantibodies Profile Detection. Biomarkers 2017, 22, 149–156. [Google Scholar] [CrossRef]
- CRISPR/Cas9 Technique for Identification of Genes Regulating Oxaliplatin Resistance of Pancreatic Cancer Cell Line|SpringerLink. Available online: https://link.springer.com/article/10.1007/s12668-016-0272-3 (accessed on 23 December 2021).
- Joung, J.; Konermann, S.; Gootenberg, J.S.; Abudayyeh, O.O.; Platt, R.J.; Brigham, M.D.; Sanjana, N.E.; Zhang, F. Genome-Scale CRISPR-Cas9 Knockout and Transcriptional Activation Screening. Nat. Protoc. 2017, 12, 828–863. [Google Scholar] [CrossRef] [Green Version]
- Kasap, C.; Elemento, O.; Kapoor, T.M. DrugTargetSeqR: A Genomics- and CRISPR-Cas9-Based Method to Analyze Drug Targets. Nat. Chem. Biol. 2014, 10, 626–628. [Google Scholar] [CrossRef] [Green Version]
- Shalem, O.; Sanjana, N.E.; Hartenian, E.; Shi, X.; Scott, D.A.; Mikkelson, T.; Heckl, D.; Ebert, B.L.; Root, D.E.; Doench, J.G.; et al. Genome-Scale CRISPR-Cas9 Knockout Screening in Human Cells. Science 2014, 343, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Sarr, A.; Bré, J.; Um, I.H.; Chan, T.H.; Mullen, P.; Harrison, D.J.; Reynolds, P.A. Genome-Scale CRISPR/Cas9 Screen Determines Factors Modulating Sensitivity to ProTide NUC-1031. Sci. Rep. 2019, 9, 7643. [Google Scholar] [CrossRef]
- Wang, T.; Wei, J.J.; Sabatini, D.M.; Lander, E.S. Genetic Screens in Human Cells Using the CRISPR-Cas9 System. Science 2014, 343, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Nurgalieva, A.K.; Skripova, V.S.; Minigulova, L.F.; Kiyamova, R.G. Obtaining a Pancreatic Cancer Cell Line Stably Expressing Doxycycline-Dependent Endonuclease Cas9. Uchenye Zap. Kazan. Univ. Seriya Estestv. Nauk. 2018, 160, 386–394. [Google Scholar]
- Bruno, P.M.; Liu, Y.; Park, G.Y.; Murai, J.; Koch, C.E.; Eisen, T.J.; Pritchard, J.R.; Pommier, Y.; Lippard, S.J.; Hemann, M.T. A Subset of Platinum-Containing Chemotherapeutic Agents Kill Cells by Inducing Ribosome Biogenesis Stress Rather than by Engaging a DNA Damage Response. Nat. Med. 2017, 23, 461–471. [Google Scholar] [CrossRef]
- A Mammalian Functional-Genetic Approach to Characterizing Cancer Therapeutics|Nature Chemical Biology. Available online: https://www.nature.com/articles/nchembio.503 (accessed on 23 December 2021).
- Riddell, I.A. Cisplatin and Oxaliplatin: Our Current Understanding of Their Actions. Met. Ions. Life Sci. 2018, 18, 199–218. [Google Scholar] [CrossRef]
- Poirier, J.T. CRISPR Libraries and Screening. Prog. Mol. Biol. Transl. Sci. 2017, 152, 69–82. [Google Scholar] [CrossRef]
- Parnas, O.; Jovanovic, M.; Eisenhaure, T.M.; Herbst, R.H.; Dixit, A.; Ye, C.J.; Przybylski, D.; Platt, R.J.; Tirosh, I.; Sanjana, N.E.; et al. A Genome-Wide CRISPR Screen in Primary Immune Cells to Dissect Regulatory Networks. Cell 2015, 162, 675–686. [Google Scholar] [CrossRef] [Green Version]
- A Highly Annotated Database of Genes Associated with Platinum Resistance in Cancer|Oncogene. Available online: https://www.nature.com/articles/s41388-021-02055-2 (accessed on 23 December 2021).
- Du, J.; Gu, J.; Li, J. Mechanisms of Drug Resistance of Pancreatic Ductal Adenocarcinoma at Different Levels. Biosci. Rep. 2020, 40, BSR20200401. [Google Scholar] [CrossRef]
- Zeng, S.; Pöttler, M.; Lan, B.; Grützmann, R.; Pilarsky, C.; Yang, H. Chemoresistance in Pancreatic Cancer. Int. J. Mol. Sci. 2019, 20, 4504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constam, D.B.; Tobler, A.R.; Rensing-Ehl, A.; Kemler, I.; Hersh, L.B.; Fontana, A. Puromycin-Sensitive Aminopeptidase: Sequence analysis, expression, and functional characterization. J. Biol. Chem. 1995, 270, 26931–26939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Somyajit, K.; Narita, T.; Maskey, E.; Stanlie, A.; Kremer, M.; Typas, D.; Lammers, M.; Mailand, N.; Nussenzweig, A.; et al. DNA Repair Network Analysis Reveals Shieldin as a Key Regulator of NHEJ and PARP Inhibitor Sensitivity. Cell 2018, 173, 972–988.e23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, K.; Yang, Y.-G.; Pierce, A.J.; Taniguchi, T.; Digweed, M.; D’Andrea, A.D.; Wang, Z.-Q.; Jasin, M. Human Fanconi Anemia Monoubiquitination Pathway Promotes Homologous DNA Repair. Proc. Natl. Acad. Sci. USA 2005, 102, 1110–1115. [Google Scholar] [CrossRef] [Green Version]
- Santidrian, A.F.; Matsuno-Yagi, A.; Ritland, M.; Seo, B.B.; LeBoeuf, S.E.; Gay, L.J.; Yagi, T.; Felding-Habermann, B. Mitochondrial Complex I Activity and NAD+/NADH Balance Regulate Breast Cancer Progression. J. Clin. Investig. 2013, 123, 1068–1081. [Google Scholar] [CrossRef] [Green Version]
- Wangpaichitr, M.; Theodoropoulos, G.; Nguyen, D.J.M.; Wu, C.; Spector, S.A.; Feun, L.G.; Savaraj, N. Cisplatin Resistance and Redox-Metabolic Vulnerability: A Second Alteration. Int. J. Mol. Sci. 2021, 22, 7379. [Google Scholar] [CrossRef]
- Yu, W.; Chen, Y.; Dubrulle, J.; Stossi, F.; Putluri, V.; Sreekumar, A.; Putluri, N.; Baluya, D.; Lai, S.Y.; Sandulache, V.C. Cisplatin Generates Oxidative Stress Which Is Accompanied by Rapid Shifts in Central Carbon Metabolism. Sci. Rep. 2018, 8, 4306. [Google Scholar] [CrossRef] [Green Version]
- ClusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters|OMICS: A Journal of Integrative Biology. Available online: https://www.liebertpub.com/doi/10.1089/omi.2011.0118 (accessed on 23 December 2021).



{kind=link}
{kind=link}
| Name of sgRNA Library | Number of sgRNA 1 | sgRNA Per Gene | Number of Target Genes |
|---|---|---|---|
| Cell cycle/ nuclear proteins genes (“CCN”) | 50,000 | up to 52 | 4799 |
| Genome-Wide (“GW”) | 90,000 | 4–5 | 18,166 |
| Candidate genes changing sensitivity of MIA PaCa-2 pancreatic cancer cells to oxaliplatin, n = 146 | |
|---|---|
| CRISPR/Cas9 screening with cell cycle/nuclear proteins sgRNA library | |
| knockout led to increase in sensitivity to oxaliplatin (n = 7): | |
| BRIP1, ERCC4, FANCD2, FANCG, FANCI, MAD2L2, NPEPPS | |
| knockout led to decrease in sensitivity to oxaliplatin (n = 1): | |
| PPP2R2A | |
| CRISPR/Cas9 screening with genome-wide sgRNA library | |
| knockout led to increase in sensitivity to oxaliplatin (n = 96): | |
| ACOX1, AP2M1, ATF6B, ATF7IP2, BPI, BTK, C14orf93, C17orf70, CDC42EP3, CDKN1B, CERS3, CERS6, COX7A2L, CRYBB1, DDAH1, DDX27, DEFA6, EMC2, EPB41L3, ESCO1, FAM160A2, FAM209A, FANCD2, FCHO2, FGFRL1, FOCAD, GCLM, GFOD2, GJB6, GNAQ, GNGT1, H2BFWT, HERC6, HEYL, HIST1H1T, HMG20B, HMGCS2, HSD3B2, KDM2B, KRTAP10-8, LEFTY1, LGI2, LMAN1, LPO, LRP1, LRRC26, MAD2L2, MAN2B1, MAP7D1, MAPK15, MARCKSL1, MCM9, MDN1, MEF2A, METRN, MPLKIP, MYO1G, NGLY1, NIPSNAP3B, NPEPPS, NPRL2, ORC6, PAX7, PCGF1, PDIA2, PEF1, PIEZO1, PKD2L2, PLAGL2, PPY, PRF1, PRRC2A, PTBP2, PVRL4, RASSF5, RGL2, RPA4, RSPH10B, SEMA3G, SMC5, SMOX, SMR3B, SYNGR2, TEKT2, TMEM185B, TPM3, TRIM4, UBE2N, UBE2T, UBE2V2, UNC5B, VCY, VCY1B, WBP11, ZNF474, ZNF804A | |
| knockout led to decrease in sensitivity to oxaliplatin (n = 45): | |
| AGO1, AGO2, ALDH1A1, ANKHD1, ATP5O, C14orf105, CAMTA2, CCDC102B, CLDN8, CREBL2, DDX26B, DYRK1A, EYA3, GAS7, GATS, GDAP1L1, GPX8, LCA5L, LGSN, LOC100127983, MAPK14, MED29, MIS18A, MRPL51, MRPS26, MYOZ2, NDUFB10, NDUFC2, NDUFC2-KCTD14, NDUFS8, NUDCD2, PPAPDC2, RBBP7, RECK, RIPPLY3, SLC38A2, SMAD2, STK11, TEAD2, TLL1, TMC5, TNFRSF12A, ZBTB18, ZNF333, ZNF536 | |
| Candidate genes changing sensitivity of MIA PaCa-2 pancreatic cancer cells to cisplatin, n = 34 | |
| CRISPR/Cas9 screening with cell cycle/nuclear proteins sgRNA library | |
| knockout led to increase in sensitivity to cisplatin (n = 32): | |
| AUNIP, BRCA1, BRIP1, CAB39, CDCA5, DBF4, DDX11, ERCC4, ESCO2, EXO1, FANCD2, FANCG, FANCI, FBXW7, INCA1, KPNA2, MAD2L2, MND1, NBN, NCAPG2, NEUROD6, NPEPPS, PLEKHA7, PPP1R12A, PSMC3IP, PSME3, RAD51B, RAD9A, RHNO1, STRA13, XRCC2, ZNF318 | |
| knockout led to decrease in sensitivity to cisplatin (n = 2): | |
| PMS2, TXNRD1 | |
| Total list of candidate genes, n = 173 | |
|---|---|
| First time associated with cancer platinum sensitivity/resistance, n = 148 | |
| knockout led to increase in sensitivity to platinum drugs (n = 109): | |
| ACOX1, AP2M1, ATF6B, ATF7IP2, AUNIP, BPI, BTK, C14orf93, C17orf70, CAB39, CDC42EP3, CDCA5, CERS3, CERS6, COX7A2L, CRYBB1, DBF4, DDAH1, DDX11, DDX27, DEFA6, EMC2, EPB41L3, ESCO1, ESCO2, FAM160A2, FAM209A, FBXW7, FCHO2, FGFRL1, FOCAD, GFOD2, GJB6, GNAQ, GNGT1, H2BFWT, HERC6, HEYL, HIST1H1T, HMG20B, HMGCS2, HSD3B2, INCA1, KDM2B, KPNA2, KRTAP10-8, LEFTY1, LGI2, LMAN1, LPO, LRRC26, MAN2B1, MAP7D1, MAPK15, MARCKSL1, MDN1, MEF2A, METRN, MND1, MPLKIP, MYO1G, NCAPG2, NEUROD6, NGLY1, NIPSNAP3B, NPEPPS, NPRL2, ORC6, PAX7, PCGF1, PDIA2, PEF1, PIEZO1, PKD2L2, PLAGL2, PLEKHA7, PPP1R12A, PPY, PRF1, PRRC2A, PSMC3IP, PSME3, PTBP2, PVRL4, RAD9A, RASSF5, RGL2, RHNO1, RPA4, RSPH10B, SEMA3G, SMC5, SMOX, SMR3B, STRA13, SYNGR2, TEKT2, TMEM185B, TPM3, TRIM4, UBE2N, UBE2V2, UNC5B, VCY, VCY1B, WBP11, ZNF318, ZNF474, ZNF804A | |
| knockout led to decrease in sensitivity to platinum drugs (n = 39): | |
| AGO1, AGO2, ANKHD1, ATP5O, C14orf105, CAMTA2, CCDC102B, CLDN8, CREBL2, DDX26B, DYRK1A, EYA3, GATS, GDAP1L1, GPX8, LCA5L, LGSN, LOC100127983, MED29, MIS18A, MRPL51, MRPS26, MYOZ2, NDUFB10, NDUFC2, NDUFC2-KCTD14, NUDCD2, PPAPDC2, RBBP7, RECK, RIPPLY3, SLC38A2, SMAD2, TEAD2, TLL1, TMC5, ZBTB18, ZNF333, ZNF536 | |
| Previously associated with platinum sensitivity/resistance, n = 25 | |
| knockout led to increase in sensitivity to platinum drugs (n = 16): | |
| BRCA1, BRIP1, CDKN1B, ERCC4, EXO1, FANCD2, FANCG, FANCI, GCLM, LRP1, MAD2L2, MCM9, NBN, RAD51B, UBE2T, XRCC2 | |
| knockout led to decrease in sensitivity to platinum drugs (n = 9): | |
| ALDH1A1, GAS7, MAPK14, NDUFS8, PMS2, PPP2R2A, STK11, TNFRSF12A, TXNRD1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skripova, V.; Vlasenkova, R.; Zhou, Y.; Astsaturov, I.; Kiyamova, R. Identification of New Regulators of Pancreatic Cancer Cell Sensitivity to Oxaliplatin and Cisplatin. Molecules 2022, 27, 1289. https://doi.org/10.3390/molecules27041289
Skripova V, Vlasenkova R, Zhou Y, Astsaturov I, Kiyamova R. Identification of New Regulators of Pancreatic Cancer Cell Sensitivity to Oxaliplatin and Cisplatin. Molecules. 2022; 27(4):1289. https://doi.org/10.3390/molecules27041289
Chicago/Turabian StyleSkripova, Vera, Ramilia Vlasenkova, Yan Zhou, Igor Astsaturov, and Ramziya Kiyamova. 2022. "Identification of New Regulators of Pancreatic Cancer Cell Sensitivity to Oxaliplatin and Cisplatin" Molecules 27, no. 4: 1289. https://doi.org/10.3390/molecules27041289
APA StyleSkripova, V., Vlasenkova, R., Zhou, Y., Astsaturov, I., & Kiyamova, R. (2022). Identification of New Regulators of Pancreatic Cancer Cell Sensitivity to Oxaliplatin and Cisplatin. Molecules, 27(4), 1289. https://doi.org/10.3390/molecules27041289