
Evaluating Phytochemical Profiles, Cytotoxicity, Antiviral Activity, Antioxidant Potential, and Enzyme Inhibition of Vepris boiviniana Extracts

 ,
,  ,
,  ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Characterization
2.2. Total Phenolic–Flavonoid Content
2.3. Antioxidant Properties
2.4. Enzymes Inhibition Properties
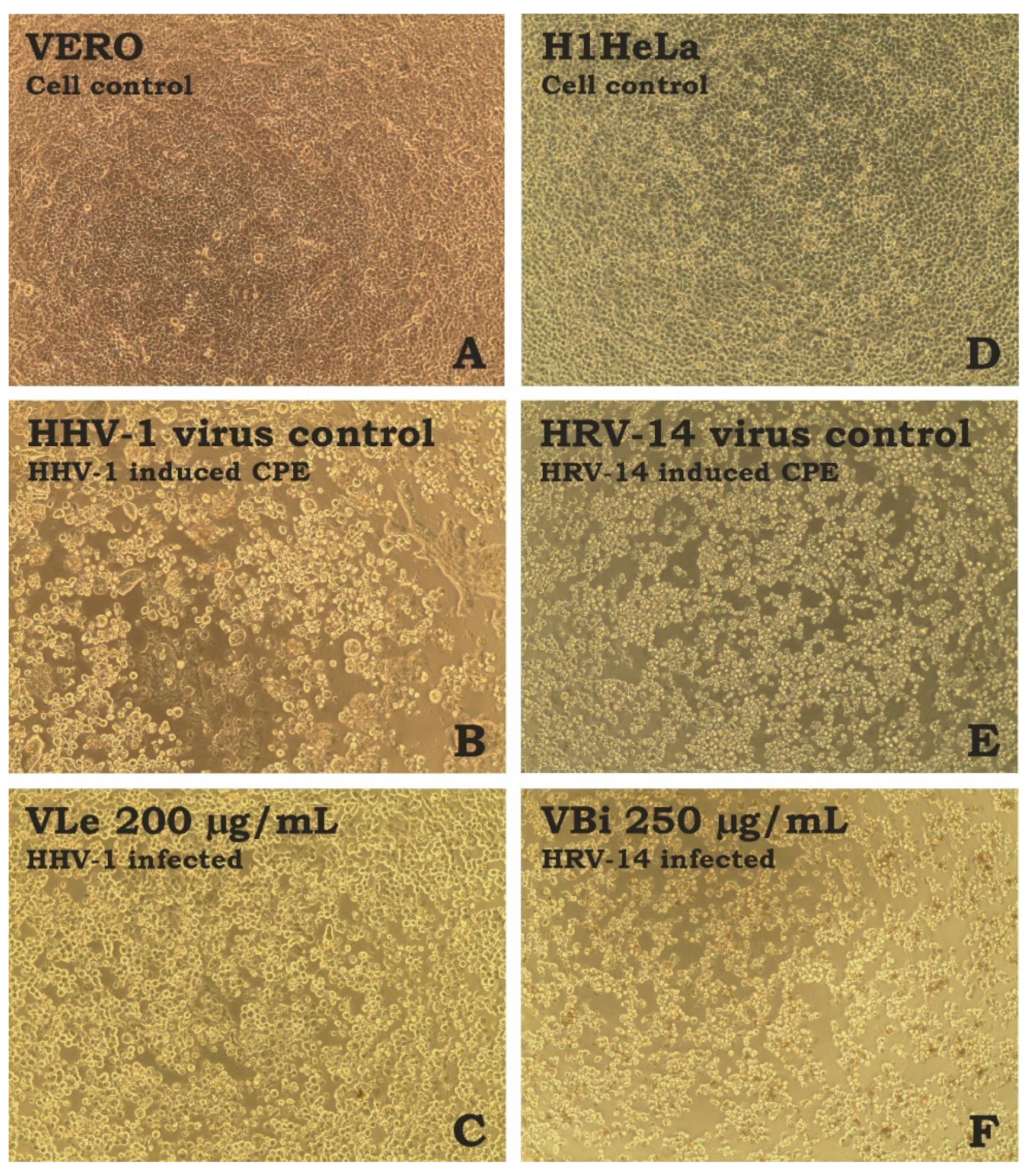
2.5. Cytotoxicity and Antiviral Properties
3. Materials and Methods
3.1. Plant Material
3.2. Sample Preparation
3.3. Total Phenolics and Flavonoids Content
3.4. Liquid Chromatography—Mass Spectrometry Analysis
3.5. In Vitro Antioxidant Assays
3.6. Enzyme Inhibitory Activity
3.7. Cytotoxicity and Antiviral Assays
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dincheva, I.; Badjakov, I.; Galunska, B. New Insights into the Research of Bioactive Compounds from Plant Origins with Nutraceutical and Pharmaceutical Potential. Plants 2023, 12, 258. [Google Scholar] [CrossRef] [PubMed]
- Nwozo, O.S.; Effiong, E.M.; Aja, P.M.; Awuchi, C.G. Antioxidant, phytochemical, and therapeutic properties of medicinal plants: A review. Int. J. Food Prop. 2023, 26, 359–388. [Google Scholar] [CrossRef]
- Buenz, E.J.; Verpoorte, R.; Bauer, B.A. The ethnopharmacologic contribution to bioprospecting natural products. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 509–530. [Google Scholar] [CrossRef]
- Mustafa, G.; Arif, R.; Atta, A.; Sharif, S.; Jamil, A. Bioactive compounds from medicinal plants and their importance in drug discovery in Pakistan. Matrix Sci. Pharma 2017, 1, 17–26. [Google Scholar] [CrossRef]
- Maggi, F.; Fortuné Randriana, R.; Rasoanaivo, P.; Nicoletti, M.; Quassinti, L.; Bramucci, M.; Lupidi, G.; Petrelli, D.; Vitali, L.A.; Papa, F. Chemical composition and in vitro biological activities of the essential oil of Vepris macrophylla (Baker) I. Verd. endemic to Madagascar. Chem. Biodivers. 2013, 10, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Koffi, Y.; Gleye, J.; Moulis, C.; Stanislas, E. Acridones from Vepris fitoravina and Vepris macrophylla. Planta Medica 1987, 53, 570–571. [Google Scholar] [CrossRef]
- Prakash Chaturvedula, V.; Schilling, J.K.; Miller, J.S.; Andriantsiferana, R.; Rasamison, V.E.; Kingston, D.G. New Cytotoxic Alkaloids from the Wood of Vepris punctata from the Madagascar Rainforest. J. Nat. Prod. 2003, 66, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Saive, M.; Frederich, M.; Fauconnier, M.-L. Plants used in traditional medicine and cosmetics in Mayotte Island (France): An ethnobotanical study. Indian J. Tradit. Knowl. 2018, 17, 645–653. [Google Scholar]
- Humbert, H.; Leroy, J.-F. Flore de Madagascar et des Comores; Biodiversity Heritage Library: Washington, DC, USA, 1960. [Google Scholar]
- Saive, M.; Frederich, M.; Fauconnier, M.-L. Plants used in traditional medicine in the Comoros archipelago: A review. Biotechnol. Agron. Soc. Environ. 2020, 24, 117–141. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MS n identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Clifford, M.N.; Wu, W.; Kuhnert, N. The chlorogenic acids of Hemerocallis. Food Chem. 2006, 95, 574–578. [Google Scholar] [CrossRef]
- Ramabulana, A.-T.; Steenkamp, P.; Madala, N.; Dubery, I.A. Profiling of chlorogenic acids from Bidens pilosa and differentiation of closely related positional isomers with the aid of UHPLC-QTOF-MS/MS-based in-source collision-induced dissociation. Metabolites 2020, 10, 178. [Google Scholar] [CrossRef]
- Yuzuak, S.; Ballington, J.; Xie, D.-Y. HPLC-qTOF-MS/MS-based profiling of flavan-3-ols and dimeric proanthocyanidins in berries of two muscadine grape hybrids FLH 13-11 and FLH 17-66. Metabolites 2018, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Rockenbach, I.I.; Jungfer, E.; Ritter, C.; Santiago-Schübel, B.; Thiele, B.; Fett, R.; Galensa, R. Characterization of flavan-3-ols in seeds of grape pomace by CE, HPLC-DAD-MSn and LC-ESI-FTICR-MS. Food Res. Int. 2012, 48, 848–855. [Google Scholar] [CrossRef]
- Hu, P.; Liang, Q.-L.; Luo, G.-A.; Zhao, Z.-Z.; Jiang, Z.-H. Multi-component HPLC fingerprinting of Radix Salviae Miltiorrhizae and its LC-MS-MS identification. Chem. Pharm. Bull. 2005, 53, 677–683. [Google Scholar] [CrossRef]
- Li, W.; Sun, Y.; Liang, W.; Fitzloff, J.F.; van Breemen, R.B. Identification of caffeic acid derivatives in Actea racemosa (Cimicifuga racemosa, black cohosh) by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Sojinu, O.S. Simultaneous determination of caffeic acid phenethyl ester and its metabolite caffeic acid in dog plasma using liquid chromatography tandem mass spectrometry. Talanta 2012, 94, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Prior, R.L. Screening of foods containing proanthocyanidins and their structural characterization using LC-MS/MS and thiolytic degradation. J. Agric. Food Chem. 2003, 51, 7513–7521. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-J.; Wang, Z.-B.; Mi, Y.-Y.; Gao, M.-J.; Lv, J.-N.; Meng, Y.-H.; Yang, B.-Y.; Kuang, H.-X. UHPLC-MS/MS determination, pharmacokinetic, and bioavailability study of taxifolin in rat plasma after oral administration of its nanodispersion. Molecules 2016, 21, 494. [Google Scholar] [CrossRef]
- Wolfender, J.-L.; Waridel, P.; Ndjoko, K.; Hobby, K.; Major, H.; Hostettmann, K. Evaluation of Q-TOF-MS/MS and multiple stage IT-MSn for the dereplication of flavonoids and related compounds in crude plant extracts. Analusis 2000, 28, 895–906. [Google Scholar] [CrossRef]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of extraction solvent/technique on the antioxidant activity of selected medicinal plant extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef] [PubMed]
- Waszkowiak, K.; Gliszczyńska-Świgło, A. Binary ethanol–water solvents affect phenolic profile and antioxidant capacity of flaxseed extracts. Eur. Food Res. Technol. 2016, 242, 777–786. [Google Scholar] [CrossRef]
- Chew, K.; Khoo, M.; Ng, S.; Thoo, Y.Y.; Aida, W.W.; Ho, C.W. Effect of ethanol concentration, extraction time and extraction temperature on the recovery of phenolic compounds and antioxidant capacity of Orthosiphon stamineus extracts. Int. Food Res. J. 2011, 18, 1427. [Google Scholar]
- Abdoulaye, M.; Clément, S.; Rodrigue, K.T.F.; Steve, N.S.F.; Adji, M.B.; Ahmed, A. Phytochemical screening, DPPH scavenging and antimicrobial activities of leaves of Eucalyptus camaldulensis, Cassia mimosoides and Vepris heterophylla from Northern Cameroon. J. Pharmacogn. Phytochem. 2019, 8, 1814–1819. [Google Scholar]
- Omujal, F.; Tenda, K.I.; Lutoti, S.; Kirabo, I.; Kasango, S.D.; Nambatya, K.G. Phytochemistry and anti-inflammatory activity of ethanolic root bark extract of Vepris nobilis Mziray (Rutaceae family). Sci. Afr. 2020, 9, e00484. [Google Scholar] [CrossRef]
- Acquaviva, A.; Di Simone, S.C.; Nilofar; Bouyahya, A.; Zengin, G.; Recinella, L.; Leone, S.; Brunetti, L.; Uba, A.I.; Guler, O. Screening for Chemical Characterization and Pharmacological Properties of Different Extracts from Nepeta italica. Plants 2023, 12, 2785. [Google Scholar] [CrossRef]
- Kainama, H.; Fatmawati, S.; Santoso, M.; Papilaya, P.M.; Ersam, T. The relationship of free radical scavenging and total phenolic and flavonoid contents of Garcinia lasoar PAM. Pharm. Chem. J. 2020, 53, 1151–1157. [Google Scholar] [CrossRef]
- Kitzberger, C.S.G.; Smânia Jr, A.; Pedrosa, R.C.; Ferreira, S.R.S. Antioxidant and antimicrobial activities of shiitake (Lentinula edodes) extracts obtained by organic solvents and supercritical fluids. J. Food Eng. 2007, 80, 631–638. [Google Scholar] [CrossRef]
- Ntchapda, F.; Bonabe, C.; Kemeta Azambou, D.R.; Talla, E.; Dimo, T. Diuretic and antioxidant activities of the aqueous extract of leaves of Vepris heterophylla (Engl.) R. Let (Rutaceae) in rats. BMC Complement. Altern. Med. 2016, 16, 516. [Google Scholar] [CrossRef]
- Ojuka, P.; Kimani, N.M.; Apollo, S.; Nyariki, J.; Ramos, R.S.; Santos, C.B. Phytochemistry of the Vepris genus plants: A review and in silico analysis of their ADMET properties. S. Afr. J. Bot. 2023, 157, 106–114. [Google Scholar] [CrossRef]
- Kiplimo, J.J.; Islam, M.S.; Koorbanally, N.A. A novel flavonoid and furoquinoline alkaloids from Vepris glomerata and their antioxidant activity. Nat. Prod. Commun. 2011, 6, 1934578X1100601215. [Google Scholar] [CrossRef]
- Imbenzi, P.S.; Osoro, E.K.; Aboud, N.S.; Ombito, J.O.; Cheplogoi, P.K. A review on chemistry of some species of genus Vepris (Rutaceae family). J. Sci. Innov. Res. 2014, 3, 357–362. [Google Scholar] [CrossRef]
- Acquaviva, A.; Nilofar; Bouyahya, A.; Zengin, G.; Di Simone, S.C.; Recinella, L.; Leone, S.; Brunetti, L.; Uba, A.I.; Cakilcioğlu, U. Chemical Characterization of Different Extracts from Artemisia annua and Their Antioxidant, Enzyme Inhibitory and Anti-Inflammatory Properties. Chem. Biodivers. 2023, 20, e202300547. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. Enzyme inhibitors: Strategies and challenges in drug design. Mol. Enzymol. Drug Targe 2023, 9, 1–4. [Google Scholar]
- de Oliveira, P.C.O.; Tinoco, L.W.; Cardoso, C.L.; Cass, Q.B.; de Moraes, M.C. Advances in screening assays for identifying cholinesterase ligands. TrAC Trends Anal. Chem. 2023, 168, 117362. [Google Scholar] [CrossRef]
- Mata, R.; Flores-Bocanegra, L.; Ovalle-Magallanes, B.; Figueroa, M. Natural products from plants targeting key enzymes for the future development of antidiabetic agents. Nat. Prod. Rep. 2023, 40, 1198–1249. [Google Scholar] [CrossRef]
- Vaezi, M. Structure and inhibition mechanism of some synthetic compounds and phenolic derivatives as tyrosinase inhibitors: Review and new insight. J. Biomol. Struct. Dyn. 2023, 41, 4798–4810. [Google Scholar] [CrossRef] [PubMed]
- Temel, H.; Atlan, M.; Ertas, A.; Yener, I.; Akdeniz, M.; Yazan, Z.; Yilmaz, M.A.; Doganyigit, Z.; Okan, A.; Akyuz, E. Cream production and biological in vivo/in vitro activity assessment of a novel boron-based compound derived from quercetin and phenyl boronic acid. J. Trace Elem. Med. Biol. 2022, 74, 127073. [Google Scholar] [CrossRef]
- Yu, Q.; Fan, L.; Duan, Z. Five individual polyphenols as tyrosinase inhibitors: Inhibitory activity, synergistic effect, action mechanism, and molecular docking. Food Chem. 2019, 297, 124910. [Google Scholar] [CrossRef]
- Si, Y.-X.; Wang, Z.-J.; Park, D.; Jeong, H.O.; Ye, S.; Chung, H.Y.; Yang, J.-M.; Yin, S.-J.; Qian, G.-Y. Effects of isorhamnetin on tyrosinase: Inhibition kinetics and computational simulation. Biosci. Biotechnol. Biochem. 2012, 76, 1091–1097. [Google Scholar] [CrossRef]
- Geran, R.; Greenberg, N.; Macdonald, M.; Schumacher, A. Tumors and Other Biological Systems. Cancer Chemother. Rep. 1972, 13, 1–87. [Google Scholar]
- Sulik, M.; Maj, E.; Wietrzyk, J.; Huczyński, A.; Antoszczak, M. Synthesis and anticancer activity of dimeric polyether ionophores. Biomolecules 2020, 10, 1039. [Google Scholar] [CrossRef] [PubMed]
- Tram, N.T.T.; Anh, D.H.; Thuc, H.H.; Tuan, N.T. Investigation of chemical constituents and cytotoxic activity of the lichen Usnea undulata. Vietnam J. Chem. 2020, 58, 63–66. [Google Scholar] [CrossRef]
- Ombito, J.O.; Chi, G.F.; Wansi, J.D. Ethnomedicinal uses, phytochemistry, and pharmacology of the genus Vepris (Rutaceae): A review. J. Ethnopharmacol. 2021, 267, 113622. [Google Scholar] [CrossRef]
- Popp, F.D.; Chakraborty, D.P. Alkaloids of the Bark of Teclea grandifolia. J. Pharm. Sci. 1964, 53, 968. [Google Scholar] [CrossRef] [PubMed]
- Gessler, M.C.; Tanner, M.; Chollet, J.; Nkunya, M.H.H.; Heinrich, M. Tanzanian medicinal plants used traditionally for the treatment of malaria: In vivo antimalarial and in vitro cytotoxic activities. Phytother. Res. 1995, 9, 504–508. [Google Scholar] [CrossRef]
- Moshi, M.J.; Kamuhabwa, A.; Mbwambo, Z.; De Witte, P. Cytotoxic Screening of Some Tanzania Medicinal Plants. East Cent. Afr. J. Pharm. Sci. 2005, 6, 52–56. [Google Scholar] [CrossRef]
- Kuete, V.; Fankam, A.G.; Wiench, B.; Efferth, T. Cytotoxicity and Modes of Action of the Methanol Extracts of Six Cameroonian Medicinal Plants against Multidrug-Resistant Tumor Cells. Evid.-Based Complement. Altern. Med. 2013, 2013, 285903. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Y.; Jiang, X.; Zhang, H.; Gao, Z.; Li, Y.; Fu, R.; Li, L.; Li, J.; Cui, H. ROS-mediated activation and mitochondrial translocation of CaMKII contributes to Drp1-dependent mitochondrial fission and apoptosis in triple-negative breast cancer cells by isorhamnetin and chloroquine. J. Exp. Clin. Cancer Res. 2019, 38, 225. [Google Scholar] [CrossRef]
- Li, C.; Yang, X.; Chen, C.; Cai, S.; Hu, J. Isorhamnetin suppresses colon cancer cell growth through the PI3K-Akt-mTOR pathway. Mol. Med. Rep. 2014, 9, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Zhai, T.; Zhang, X.; Hei, Z.; Jin, L.; Han, C.; Ko, A.T.; Yu, X.; Wang, J. Isorhamnetin inhibits human gallbladder cancer cell proliferation and metastasis via PI3K/AKT signaling pathway inactivation. Front. Pharmacol. 2021, 12, 628621. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, B.; Pu, N.; Ran, X.; Lian, T.; Cai, Y.; Xing, W.; Sun, K. Isorhamnetin suppresses human gastric cancer cell proliferation through mitochondria-dependent apoptosis. Molecules 2022, 27, 5191. [Google Scholar] [CrossRef]
- Duan, R.; Liang, X.; Chai, B.; Zhou, Y.; Du, H.; Suo, Y.; Chen, Z.; Li, Q.; Huang, X. Isorhamnetin induces melanoma cell apoptosis via the PI3K/Akt and NF-κB pathways. BioMed Res. Int. 2020, 2020, 1057943. [Google Scholar] [CrossRef]
- Wang, M.; Xu, Z.; Cai, Q.; Deng, Y.; Shi, W.; Zhou, H.; Wang, D.; Li, J. Isorhamnetin inhibits progression of ovarian cancer by targeting ESR1. Ann. Transl. Med. 2022, 10, 1216. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-L.; Quan, Q.; Ji, R.; Guo, X.-Y.; Zhang, J.-M.; Li, X.; Liu, Y.-G. Isorhamnetin suppresses PANC-1 pancreatic cancer cell proliferation through S phase arrest. Biomed. Pharmacother. 2018, 108, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Zhang, Y.; Li, J.; Huang, S.; Gao, R. Isorhamnetin inhibited the proliferation and metastasis of androgen-independent prostate cancer cells by targeting the mitochondrion-dependent intrinsic apoptotic and PI3K/Akt/mTOR pathway. Biosci. Rep. 2020, 40, BSR20192826. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Lee, D.-E.; Lee, K.W.; Son, J.E.; Seo, S.K.; Li, J.; Jung, S.K.; Heo, Y.-S.; Mottamal, M.; Bode, A.M. Isorhamnetin suppresses skin cancer through direct inhibition of MEK1 and PI3-K. Cancer Prev. Res. 2011, 4, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Xiao, Y.; Liu, S.; Luo, F.; Tang, D.; Yu, Y.; Xie, Y. Isorhamnetin induces cell cycle arrest and apoptosis by triggering DNA damage and regulating the AMPK/mTOR/p70S6K signaling pathway in doxorubicin-resistant breast cancer. Phytomedicine 2023, 114, 154780. [Google Scholar] [CrossRef]
- Sarkar, S.; Das, A.K.; Bhattacharya, S.; Gachhui, R.; Sil, P.C. Isorhamnetin exerts anti-tumor activity in DEN+ CCl4-induced HCC mice. Med. Oncol. 2023, 40, 188. [Google Scholar] [CrossRef] [PubMed]
- Llorent-Martínez, E.J.; Ruiz-Medina, A.; Zengin, G.; Ak, G.; Jugreet, S.; Mahomoodally, M.F.; Emre, G.; Orlando, G.; Libero, M.L.; Nilofar. New Biological and Chemical Evidences of Two Lamiaceae Species (Thymbra capitata and Thymus sipyleus subsp. rosulans): In Vitro, In Silico and Ex Vivo Approaches. Molecules 2022, 27, 9029. [Google Scholar]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G. A study on in vitro enzyme inhibitory properties of Asphodeline anatolica: New sources of natural inhibitors for public health problems. Ind. Crops Prod. 2016, 83, 39–43. [Google Scholar] [CrossRef]
- Świątek, Ł.; Wasilewska, I.; Boguszewska, A.; Grzegorczyk, A.; Rezmer, J.; Rajtar, B.; Polz-Dacewicz, M.; Sieniawska, E. Herb Robert’s Gift against Human Diseases: Anticancer and Antimicrobial Activity of Geranium robertianum L. Pharmaceutics 2023, 15, 1561. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| No | tR | Molecular Formula | Negative Ion Mode | Positive Ion Mode | Tentative Idenificaction | Extracts | References | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Precursor Ion Measured (m/z) (Δ, ppm) | Predicted (m/z) | Fragment Ions (m/z) | Precursor Ion (m/z) Measured (Δ, ppm) | Predicted (m/z) | Fragment Ions (m/z) | ||||||
| 1 | 1.58 | C6H14O6 | [M − H]− 181.0718 (−0.21) | 181.0718 | 59.0140; 89.0241; 101.0242; 71.0137; 163.0606; 73.0291 | [M + Na]+ 205.0679 (1.97) | 205.0683 | 82.9994; 55.0567; 82.0674; 84.0021; 97.0191; 56.0512 | Hexitol | VLe, VLw, VLi, VBh, VBe, VBw, VBi | PubChem |
| 2 | 1.75 | C5H10O6 | [M − H]− 165.0399 (3.38) | 165.0405 | 75.0089; 129.0188; 59.0142; 147.0300; 55.0201; 87.0093 | - | - | - | Tetrahydroxypentanoic acid (Arabinoic acid) | VLe, VLw, VLi, VBe, VBw, VBi | PubChem |
| 3 | 1.84 | C6H12O7 | [M − H]− 195.0512 (−0.89) | 195.0510 | 75.0099; 129.0195; 99.0091; 59.0151; 87.0089; 177.0406 | - | - | - | Gluconic acid | VLe, VLw, VLi, VBe, VBw, VBi | PubChem |
| 4 | 1.91 | C7H12O6 | [M − H]− 191.0559 (1.10) | 191.0561 | 85.0295; 59.0139; 93.0342; 127.0389; 87.0089; 75.0088 | [M + Na]+ 215.0524 (1.09) | 215.0526 | 172.0942; 154.0841; 85.0287; 99.0435; 111.0217; 197.0415 | Quinic acid | VLe, VLw, VLi, VBh, VBe, VBw, VBi | PubChem |
| 5 | 2.18 | C4H6O5 | [M − H]− 133.0138 (3.33) | 133.0142 | 115.0028; 71.0139; 72.9934; 89.0234; 116.0060 | - | - | - | Malic acid | VLe, VLw, VLi, VBh, VBe, VBw, VBi | PubChem |
| 6 | 2.26 | C6H8O7 | [M − H]− 191.0196 (0.66) | 191.0197 | 111.0081; 173.0082; 85.0294; 154.9977; 117.0186; 72.9933 | [M + Na]+ 215.0163 (−0.40) | 215.0162 | 172.0999; 101.0275; 154.0913; 83.0177; 129.0218; 197.0082 | Citric acid | VLe, VLw, VLi, VBh, VBe, VBw, VBi | PubChem |
| 7 | 4.92 | C14H26O10 | [M − COOH]− 399.1502 (1.69) | 399.1508 | 353.1451; 207.0873; 354.1470; 101.0258; 161.0457; 59.0139 | [M + Na]+ 377.1420 (−0.51) | 377.1418 | 71.0482; 85.0289; 247.0460; 101.9770; 155.9456; 170.9016 | Carbohydrate | VBi, VBw, VBe | PubChem |
| 8 | 10.59 | C8H10O3 | [M − H]− 153.0560 (−1.83) | 153.0557 | 123.0454; 124.0489; 110.9839; 122.0364; 95.0486 | - | - | - | Hydroxytyrosol | VLw | PubChem |
| 9 | 10.76 | C16H30O11 | [M − COOH]− 443.1774 (−0.97) | 443.1770 | 397.1731; 251.1137; 101.0252; 398.1769; 161.0423; 59.0151 | [M + Na]+ 421.1682 (−0.42) | 421.1680 | 275.1098; 177.0557; 276.1119; 197.0794; 165.0535; 121.0648 | Carbohydrate | VBi, VBw, VBe | PubChem |
| 10 | 10.76 | C16H30O9 | [M − COOH]− 411.1878 (−1.68) | 411.1872 | 365.1816; 89.0246; 366.1882; 119.0359; 203.1278; 59.0145 | [M + Na]+ 389.1773 (2.47) | 389.1782 | 203.0502; 209.1096; 371.1648; 227.1205; 163.0362; 230.1345 | Terpene glycoside | VLe, VLw, VLi | PubChem |
| 11 | 11.43 | C14H20O7 | [M − COOH]− 345.1190 (0.35) | 345.1191 | 137.0598; 299.1120; 161.0444; 101.0229; 71.0128; 300.1167 | [M + Na]+ 323.1106 (−1.59) | 323.1101 | 85.0298; 69.0340; 167.0163; 121.0637; 85.0633; 203.0536 | Salidroside isomer | VLe, VLw, VLi, VBe, VBi | fragmentation PubChem |
| 12 | 11.51 | C16H30O11 | [M − COOH]− 443.1779 (−2.22) | 443.1770 | 397.1708; 251.1151; 398.1752; 101.0257; 59.0149; 71.0133 | [M + Na]+ 421.1684 (−0.92) | 421.1680 | 275.1136; 276.1106; 121.0518; 117.9783; 167.0628; 205.0273 | Carbohydrate | VBi, VBw | PubChem |
| 13 | 11.84 | C7H6O4 | [M − H]− 153.0194 (−0.44) | 153.0193 | 109.0287; 108.0222; 66.9944; 81.0323; 53.0410; 91.0199 | - | - | - | Dihydroxybenzoic acid | VLw, VLi, VBw, VBi | PubChem |
| 14 | 12.18 | C14H20O8 | [M − H]− 315.1086 (−0.19) | 315.1085 | 123.0446; 153.0552; 124.0473; 154.0587; 122.0364; 109.0279 | - | - | - | Hydroxythyrosol glucoside | VLe, VLw, VLi, VBe, VBi | fragmentation, PubChem |
| 15 | 12.93 | C19H34O9 | [M − COOH]− 451.2188 (−0.77) | 451.2185 | 405.2145; 89.0246; 179.0546; 119.0338; 59.0150; 71.0129 | [M + Na]+ 429.2080 (3.70) | 429.2095 | 268.0942; 249.1412; 201.0003; 203.0486; 158.9949; 85.0250 | Norisoprenoid glucoside derivative | VLe, VLw, VLi | fragmentation, PubChem |
| 16 | 13.35 | C16H30O9 | [M − COOH]− 411.1875 (−0.86) | 411.1872 | 89.0239; 179.0565; 59.0143; 365.1825; 119.0342; 71.0139 | [M + Na]+ 389.1772 (2.74) | 389.1782 | 135.0390; 227.1181; 359.1620; 147.0376; 203.0461; 85.0241 | Terpene glycoside | VLe, VLw, VLi | PubChem |
| 17 | 14.02 | C7H6O3 | [M − H]− 137.0244 (0.13) | 137.0244 | 108.0212; 136.0161; 92.0266; 91.0188; 53.0407; 109.0270 | - | - | - | Hydroxybenzoic acid isomer | VLe, VLw, VLi, VBe, VBw, VBi | PubChem |
| 18 | 14.35 | C19H34O9 | [M − COOH]− 451.2181 (0.95) | 451.2185 | 405.2111; 89.0240; 59.0143; 406.2163; 101.0232; 71.0146 | [M + Na]+ 429.2073 (5.43) | 429.2095 | 98.9728; 201.0012; 158.9929; 411.1956; 85.0265; 203.0539 | Norisoprenoid glucoside derivative | VLe, VLw, VLi | fragmentation, PubChem |
| 19 | 14.86 | C18H19NO4 | [M − H]− 312.1245 (−1.18) | 312.1241 | 282.0769; 297.1001; 254.0812; 239.0699; 283.0788; 224.0481 | [M + H]+ 314.1384 (0.91) | 314.1387 | 222.0662; 191.0842; 282.0879; 265.0848; 237.0896; 219.0792 | Alkaloid | VLe, VLi, VBh, VBe, VBw, VBi | PubChem |
| 20 | 14.93 | C16H18O9 | [M − H]− 353.0878 (0.02) | 353.0878 | 191.0548; 179.0343; 135.0447; 192.0587; 85.0297; 209.0292 | [M + H]+ 355.1025 (−0.40) | 355.1024 | - | Caffeoylquinic acid (3-O-caffeoylquinic acid) | VLi, VBe, VBi | PubChem [11,12,13] |
| 21 | 15.35 | C7H6O3 | [M − H]− 137.0244 (0.13) | 137.0244 | 93.0344; 65.0402; 94.0387; 66.0429; 75.0254 | [M + H]+ 139.0391 (−0.94) | 139.0390 | - | Hydroxybenzoic acid isomer | VLw, VBw | PubChem |
| 22 | 15.36 | C19H21NO4 | [M − H]− 326.1400 (−0.67) | 326.1398 | 311.1156; 268.0850; 296.0925; 239.0699; 281.0683; 224.0475 | [M + H]+ 328.1545 (−0.51) | 328.1543 | 297.1113; 265.0856; 298.1150; 266.0887; 282.0879; 237.0903 | Alkaloid | VLe, VLi, VBh, VBe, VBw, VBi | PubChem |
| 23 | 15.52 | C19H34O9 | [M − COOH]− 451.2194 (−2.25) | 451.2185 | 405.2152; 89.0239; 59.0138; 119.0331; 179.0534; 406.2163 | [M + Na]+ 429.2083 (2.96) | 429.2095 | 249.1440; 209.1512; 147.0774; 99.0438; 85.0273; 152.0693 | Norisoprenoid glucoside derivative | VLe, VLw, VLi | fragmentation, PubChem |
| 24 | 16.52 | C30H26O12 | [M − H]− 577.1350 (0.26) | 577.1351 | 125.0252; 289.0689; 407.0742; 245.0817; 151.0402; 161.0276 | [M + H]+ 579.1486 (1.91) | 579.1497 | 289.0668; 291.0834; 427.1004; 127.0366; 409.0903; 247.0570 | Procyanidin dimer type B | VBh, VBe, VBi | fragmentation, PubChem, [14] |
| 25 | 17.43 | C15H14O6 | [M − H]− 289.0717 (0.21) | 289.0718 | 245.0821; 109.0287; 203.0711; 125.0234; 205.0503; 151.0398 | [M + H]+ 291.0850 (4.53) | 291.0863 | 139.0366; 123.0418; 165.0522; 147.0417; 140.0398; 207.0619 | Epicatechin | VBh, VBe, VBi | fragmentation, PubChem, [14,15] |
| 26 | 17.77 | C16H18O9 | [M − H]− 353.0879 (−0.27) | 353.0878 | 191.0558; 192.0589; 85.0295; 193.0594; 209.0293; 161.0235 | [M + H]+ 355.1020 (1.01) | 355.1024 | - | Caffeoylquinic acid (5-O-caffeoylquinic acid) | VLe, VLw, VLi, VBh, VBe, VBi | PubChem, [11,12,13] |
| 27 | 18.27 | C16H18O9 | [M − H]− 353.0879 (−0.27) | 353.0878 | 191.0550; 173.0449; 179.0340; 135.0440; 85.0296; 192.0578 | - | - | - | Caffeoylquinic acid (4-O-caffeoylquinic acid) | VLi, VBe, VBi | PubChem, [11,12,13] |
| 28 | 18.35 | C9H8O4 | [M − H]− 179.0355 (−2.88) | 179.0350 | 135.0448; 134.0372; 89.0403; 107.0504; 79.0546; 136.0468 | - | - | - | Caffeic acid | VLw, VBi | [16,17,18] |
| 29 | 18.44 | C33H40O21 | [M − H]− 771.1985 (0.56) | 771.1989 | 299.0210; 301.0362; 300.0278; 609.1506; 462.0849; 463.0881 | [M + H]+ 773.2124 (1.40) | 773.2135 | 465.1007; 627.1523; 303.0478; 466.1045; 611.1585; 628.1545 | Pentahydroxyflavone O-rutinoside O-hexoside (Quercetin 3-O-rutinoside-7-O-glucoside) | VLe, VLi | fragmentation, PubChem |
| 30 | 18.77 | C17H32O10 | [M − COOH]− 441.1982 (−1.14) | 441.1978 | 395.1916; 249.1333; 396.1962; 101.0240; 71.0145; 59.0136 | [M + Na]+ 419.1884 (0.93) | 419.1888 | - | Carbohydrate | VBi, VBw, VBe, VLi, VLw, VLe | PubChem |
| 31 | 18.95 | C30H26O11 | [M − H]− 561.1423 (−3.67) | 561.1402 | 289.0682; 165.0551; 125.0281; 435.1033; 407.0783; | - | - | - | Procyanidin dimer ((epi)Catechin-(epi)Afzelechin) | VBe | PubChem, [19] |
| 32 | 19.53 | C33H40O20 | [M − H]− 755.2034 (0.82) | 755.2040 | 285.0402; 593.1524; 283.0235; 284.0314; 594.1574; 255.0276 | [M + H]+ 757.2195 (0.22) | 757.2197 | - | Tetrahydroxyflavone O-rutinoside O-hexoside (Kaempferol 3-O-rutinoside-7-O-glucoside) | VLe, VLi | Fragmentation, PubChem |
| 33 | 19.86 | C34H42O21 | [M − H]− 785.2160 (−1.80) | 785.2146 | 315.0504; 623.1632; 314.0402; 300.0305; 299.0178; 271.0259 | [M + H]+ 787.2292 (−0.08) | 787.2291 | - | Tetrahydroxymethoxyflavone O-rutinoside O-hexoside (Isorhamnetin 3-O-rutinoside-glucoside) | VLe, VLi | Fragmentation PubChem |
| 34 | 20.19 | C45H36O18 | [M − H]− 863.1821 (0.91) | 863.1829 | 411.0743; 289.0724; 285.0425; 711.1369; 451.1040; 412.0785 | [M + H]+ 865.1972 (0.28) | 865.1974 | 533.1065; 713.1485; 287.0526; 695.1387; 575.1167; 739.1642 | Procyanidin trimer | VLe, VLi, VBh, VBe, VBi | Fragmentation, PubChem |
| 35 | 20.22 | C16H18O8 | [M − H]− 337.0934 (−1.51) | 337.0929 | 191.0563; 93.0353; 119.0508; 87.0092; 85.0299; 163.0404 | - | - | - | Coumaroylquinic acid isomer | VLe, VLi, VBe, VBi | Fragmentation, PubChem, [11,12,13] |
| 36 | 20.95 | C39H32O15 | [M − H]− 739.1657 (1.55) | 739.1668 | 289.0721; 177.0197; 339.0505; 587.1186; 449.0865; 290.0707 | [M + H]+ 741.1817 (−0.41) | 741.1814 | - | Tannin | VBe, VBi | Fragmentation, PubChem |
| 37 | 21.19 | C16H18O8 | [M − H]− 337.0925 (1.16) | 337.0929 | 191.0547; 85.0299; 192.0574; 93.0342; 127.0398; 59.0149 | - | - | - | Coumaroylquinic acid isomer | VLe, VLi | Fragmentation, PubChem, [11,12,13] |
| 38 | 22.28 | C32H38O20 | [M − H]− 741.1896 (−1.66) | 741.1884 | 300.0286; 301.0308; 271.0255; 742.1901; 255.0309; 178.9962 | [M + H]+ 743.2028 (1.64) | 743.2040 | 303.0475; 304.0508; 465.1008; 611.1563; 743.2041; 85.0275 | Pentahydroxyflavone O-rutinoside-pentoside (Quercetin 3-O-rutinoside-pentoside) | VLe, VLw, VLi | Fragmentation, PubChem |
| 39 | 22.52 | C10H10O4 | [M − H]− 193.0509 (−1.38) | 193.0506 | 134.0375; 178.0270; 149.0610; 93.0348; 137.0228; 135.0433 | - | - | - | Isoferulic acid | VLw | PubChem |
| 40 | 22.70 | C18H34O10 | [M − COOH]− 455.2145 (−2.68) | 455.2134 | 409.2081; 263.1491; 410.2107; 101.0243; 264.1529; 411.2163 | [M + Na]+ 433.2047 (−0.69) | 433.2044 | - | Hexyl 2-O-t(6-Deoxy-Alpha-L-Galactopyranosyl)-Beta-D-Galactopyranoside | VLe, VLw, VLi, VBe, VBw, VBi | PubChem |
| 41 | 23.19 | C26H28O16 | [M − H]− 595.1312 (−1.24) | 595.1305 | 300.0288; 271.0236; 301.0319; 255.0282; 272.0292; 302.0331 | [M + H]+ 597.1458 (−1.32) | 597.1450 | - | Pentahydroxyflavone O-hexoside O-pentoside (Quercetin O-glucoside O-pentoside) | VLe, VLw, VLi | Fragmentation, PubChem |
| 42 | 23.45 | C27H30O16 | [M − H]− 609.1453 (1.33) | 609.1461 | 300.0265; 301.0334; 271.0239; 255.0279; 151.0025; 243.0297 | [M + H]+ 611.1608 (−0.23) | 611.1607 | 303.0487; 465.1022; 85.0289; 129.0542; 147.0646 | Rutin | VLe, VLw, VLi, VBe, VBi | Fragmentation, PubChem |
| 43 | 23.70 | C15H12O7 | [M − H]− 303.0513 (−0.90) | 303.0510 | 285.0401; 125.0238; 177.0200; 286.0451; 175.0402; 151.0032 | - | - | - | Pentahydroxydihydroflavone (Taxifolin) | VBe, VBi | Fragmentation, PubChem, [20] |
| 44 | 24.19 | C21H20O12 | [M − H]− 463.0889 (−1.51) | 463.0882 | 300.0261; 271.0239; 301.0323; 255.0288; 243.0277; 151.0023 | [M + H]+ 465.1023 (0.98) | 465.1028 | 303.0491; 85.0290; 61.0298; 97.0284; 91.0394; 73.0287 | Pentahydroxyflavone O-hexoside (Quercetin 3-O-glucoside) | VLe, VLw, VLi, VBe, VBi | Fragmentation, PubChem |
| 45 | 24.78 | C9H16O4 | [M − H]− 187.0978 (−1.16) | 187.0976 | 125.0971; 97.0663; 126.0998; 169.0864; 123.0799; 143.1070 | - | - | - | Azelaic acid | VLe, VLw, VLi, VBh, VBe, VBw, VBi | PubChem |
| 46 | 24.95 | C27H30O15 | [M − H]− 593.1532 (−3.38) | 593.1512 | 285.0401; 284.0329; 255.0302; 227.0356; 286.0441; 229.0504 | [M + H]+ 595.1659 (−0.26) | 595.1657 | 287.0534; 449.1074; 288.0573; 450.1108; 85.0277; 129.0532 | Tetrahydroxyflavone O-rutinoside (Kaempferol 3-O-rutinoside = Nicotiflorin) | VLe, VLw, VLi | Fragmentation, PubChem |
| 47 | 25.11 | C28H32O16 | [M − H]− 623.1636 (−2.95) | 623.1618 | 315.0483; 300.0240; 314.0411; 299.0182; 271.0225; 243.0284 | [M + H]+ 625.1754 (1.46) | 625.1763 | 317.0601; 318.0645; 479.1136; 480.1169; 85.0259; 129.0510 | Tetrahydroxymethoxyflavone O-rutinoside (Isorhamnetin 3-O-rutinoside) | VLe, VLw, VLi | Fragmentation, PubChem |
| 48 | 25.28 | C20H18O11 | [M − H]− 433.0787 (−2.45) | 433.0776 | 300.0258; 271.0255; 255.0293; 301.0340; 243.0294; 151.0036 | [M + H]+ 435.0929 (−1.64) | 435.0922 | 303.0465; 304.0503; 73.0267; 61.0270; 229.0453; 165.0153 | Pentahydroxyflavone O- pentoside (Quercetin O-pentoside) | VLe, VLw, VLi, VBi | Fragmentation |
| 49 | 25.61 | C21H18O13 | [M − H]− 477.0685 (−2.17) | 477.0675 | 301.0362; 151.0033; 178.9972; 121.0292; 107.0118; 255.0337 | - | - | - | Pentahydroxyflavone O- glucuronide (Quercetin 3-O-glucuronide) | VBe, VBi | Fragmentation, PubChem |
| 50 | 25.78 | C21H20O11 | [M − H]− 447.0941 (−1.82) | 447.0933 | 300.0270; 271.0244; 301.0337; 255.0298; 151.0033; 243.0307 | [M + H]+ 449.1073 (1.20) | 449.1078 | 303.0483; 287.0533; 85.0284; 71.0493; 57.0337; 129.0531 | Pentahydroxyflavone O-rhamnoside (Quercetin 3-O-rhamnoside) | VLe, VLw, VLi, VBh, VBe, VBi | Fragmentation, PubChem |
| 51 | 25.94 | C22H22O12 | [M − H]− 477.1030 (1.78) | 477.1038 | 314.0425; 243.0285; 271.0235; 285.0397; 257.0432; 299.0185 | [M + H]+ 479.1174 (2.10) | 479.1184 | 317.0634; 318.0662; 85.0262; 302.0393; 97.0263; 285.0365 | Tetrahydroxymethoxyflavone O-hexoside (Isorhamnetin 3-O-glucoside) | VLe, VLw, VLi | Fragmentation, PubChem |
| 52 | 26.04 | C25H24O12 | [M − H]− 515.1207 (−2.33) | 515.1195 | 353.0877; 191.0570; 179.0355; 354.0913; 135.0454; 173.0442 | - | - | - | di-Caffeoylquinic acid isomer | VLi, VBh, VBe, VBi | PubChem, [11,13] |
| 53 | 26.37 | C21H36O10 | [M − COOH]− 493.2304 (−3.01) | 493.2291 | 447.2228; 448.2254; 315.1811; 101.0241; 71.0134; 161.0448 | [M + Na]+ 471.2198 (0.60) | 471.2201 | 335.0914; 471.2188; 336.0947; 472.2213; 337.0962; 275.0689 | Terpene glycoside | VLe, VLw, VLi, VBh, VBe, VBw, VBi | PubChem |
| 54 | 27.04 | C25H24O12 | [M − H]− 515.1202 (−1.36) | 515.1195 | 353.0872; 173.0445; 179.0349; 191.0551; 354.0892; 135.0442 | - | - | - | di-Caffeoylquinic acid isomer | VLi, VBh, VBe, VBi | PubChem, [11,13] |
| 55 | 27.37 | C30H26O15 | [M − H]− 625.1199 (−0.01) | 625.1199 | 300.0268; 301.0320; 463.0909; 271.0202; 464.0909; 255.0309 | - | - | - | Pentahydroxyflavone O-Caffeoyl-hexoside (Quercetin 3-O-caffeoyl-glucoside) | VBe | fragmentation, PubChem |
| 56 | 27.70 | C21H20O10 | [M − H]− 431.0993 (−2.15) | 431.0984 | 255.0294; 227.0342; 284.0315; 285.0396; 256.0341; 228.0382 | [M + H]+ 433.1122 (1.67) | 433.1129 | 287.0526; 71.0476; 288.0560; 57.0323; 72.0504; 153.0142 | Tetrahydroxyflavone O-rhamnoside (Kaempferol 3-O-rhamnoside) | VLe, VLi, VBe, VBi | fragmentation, PubChem |
| 57 | 33.71 | C18H34O5 | [M − H]− 329.2332 (0.45) | 329.2333 | 171.1027; 211.1340; 229.1466; 139.1130; 212.1359; 99.0818 | - | - | - | Fatty acid | VLe, VLw, VLi, VBh, VBe, VBw, VBi | PubChem |
| 58 | 33.96 | C15H10O5 | [M − H]− 269.0455 (0.17) | 269.0455 | 117.0347; 65.0043; 151.0018; 107.0139; 118.0371; 149.0254 | - | - | - | Trihydroxyflavone (Apigenin) | VBe | fragmentation, PubChem, [21] |
| 59 | 34.46 | C16H12O7 | [M − H]− 315.0511 (−0.23) | 315.0510 | 300.0274; 151.0027; 63.0245; 107.0138; 108.0219; 83.0137 | - | - | - | Tetrahydroxymethoxyflavone (Isorhamnetin) | VLe | fragmentation, PubChem |
| 60 | 35.63 | C18H34O5 | [M − H]− 329.2341 (−2.28) | 329.2333 | 201.1164; 199.1327; 171.1060; 211.1342; 202.1133; 59.0147 | - | - | - | Fatty acid | VLe, VLw, VLi, VBe, VBw, VBi | PubChem |
| Part | Extracts | Total Phenolic Content (mg GAE/g) | Total Flavonoid Content (mg RE/g) |
|---|---|---|---|
| Leaves | n-hexane | 29.43 ± 3.27 g | 20.73 ± 0.58 b |
| Ethanol (80%) | 70.61 ± 1.42 c | 25.39 ± 0.68 a | |
| Water | 34.98 ± 0.25 f | 13.12 ± 0.27 c | |
| Infusion | 46.44 ± 0.62 d | 25.72 ± 0.42 a | |
| Stem barks | n-hexane | 27.49 ± 0.55 g | 3.42 ± 0.08 e |
| Ethanol (80%) | 108.19 ± 0.98 a | 6.13 ± 0.27 d | |
| Water | 42.27 ± 0.68 e | 1.17 ± 0.38 f | |
| Infusion | 75.85 ± 0.35 b | 4.11 ± 0.18 e |
| Part | Extracts | DPPH (mg TE/g) | ABTS (mg TE/g) | CUPRAC (mg TE/g) | FRAP (mg TE/g) | PBD (mmol TE/g) | MCA (mg EDTAE/g) |
|---|---|---|---|---|---|---|---|
| Leaves | n-hexane | 10.95 ± 0.34 e | 23.91 ± 0.60 g | 62.98 ± 3.01 g | 36.48 ± 0.53 f | 2.32 ± 0.06 a | 11.33 ± 0.64 d |
| Ethanol (80%) | 132.61 ± 6.07 b | 203.34 ± 5.21 c | 200.09 ± 1.15 c | 122.57 ± 0.76 c | 1.93 ± 0.01 b | 9.92 ± 0.80 e | |
| Water | 39.33 ± 0.26 d | 85.64 ± 1.71 e | 83.99 ± 0.98 f | 65.72 ± 0.44 e | 0.88 ± 0.02 g | 25.37 ± 0.19 b | |
| Infusion | 59.06 ± 1.34 c | 133.29 ± 5.21 d | 115.92 ± 1.00 d | 76.42 ± 1.17 d | 1.08 ± 0.02 f | 28.43 ± 0.04 a | |
| Stem barks | n-hexane | 30.21 ± 1.15 d | 51.26 ± 3.59 f | 65.47 ± 1.03 g | 36.78 ± 0.55 f | 1.49 ± 0.09 d | 18.31 ± 0.65 c |
| Ethanol (80%) | 388.42 ± 9.96 a | 551.82 ± 9.20 a | 512.03 ± 2.39 a | 320.28 ± 7.02 a | 2.41 ± 0.09 a | 25.47 ± 0.26 b | |
| Water | 32.26 ± 1.89 d | 88.25 ± 0.17 e | 106.51 ± 0.25 e | 80.20 ± 1.17 d | 1.27 ± 0.05 e | 28.86 ± 0.54 a | |
| Infusion | 137.39 ± 4.20 b | 216.38 ± 3.85 b | 218.46 ± 0.70 b | 138.28 ± 2.22 b | 1.70 ± 0.07 c | 29.54 ± 0.12 a |
| Part | Extracts | AChE (mg GALAE/g) | BChE (mg GALAE/g) | Tyrosinase (mg KAE/g) | Amylase (mmol ACAE/g) | Glucosidase (mmol ACAE/g) |
|---|---|---|---|---|---|---|
| Leaves | n-hexane | 1.69 ± 0.07 d | 1.62 ± 0.03 c d | 43.43 ± 0.82 d | 0.71 ± 0.03 a | 0.22 ± 0.06 e |
| Ethanol (80%) | 1.67 ± 0.08 d | 3.08 ± 0.14 a | 59.89 ± 0.45 b | 0.40 ± 0.01 c | 1.19 ± 0.01 a | |
| Water | 2.25 ± 0.03 b | 1.32 ± 0.22 d | na | 0.06 ± 0.02 f | 0.37 ± 0.05 d | |
| Infusion | 0.70 ± 0.01 e | 1.75 ± 0.14 c | na | 0.05 ± 0.01 f | 0.04 ± 0.01 f | |
| Stem barks | n-hexane | 0.66 ± 0.07 e | 2.31 ± 0.21 b | 47.01 ± 0.32 c | 0.49 ± 0.01 b | 1.08 ± 0.03 b |
| Ethanol (80%) | 2.03 ± 0.17 c | 3.08 ± 0.06 a | 64.89 ± 0.40 a | 0.47 ± 0.01 b | 1.26 ± 0.01 a | |
| Water | 2.41 ± 0.01 a | 0.23 ± 0.01 e | na | 0.13 ± 0.01 d | 0.90 ± 0.01 c | |
| Infusion | 0.33 ± 0.02 f | 1.82 ± 0.08 c | na | 0.09 ± 0.01 e | 0.97 ± 0.01 c |
| Vepris boiviniania | VERO | FaDu | H1HeLa | RKO | |||
|---|---|---|---|---|---|---|---|
| CC50 | CC50 | SI | CC50 | SI | CC50 | SI | |
| Leaves—hexane (VLh) | 63.06 ± 5.30 | 82.82 ± 9.53 | 0.76 | 26.20 ± 0.59 * | 2.41 | 22.04 ± 3.24 * | 2.86 |
| Leaves—ethanol (VLe) | 613.27 ± 62.25 | 114.90 ± 11.77 | 5.34 ** | 63.46 ± 2.03 ** | 9.66 | 119.33 ± 10.21 ** | 5.14 |
| Leaves—aqueous (VLa) | 251.4 ± 26.98 | 254.4 ± 20.52 | 0.99 | >500 | <1 | >500 | <1 |
| Leaves—infusion (VLi) | 332.8 ± 25.44 | 125.5 ± 13.33 ** | 2.65 | >500 | <1 | >500 | <1 |
| Bark—hexane (VBh) | 81.01 ± 11.24 | 56.50 ± 3.52 | 1.43 | 43.29 ± 4.06 * | 1.87 | 49.55 ± 5.32 * | 1.63 |
| Bark—ethanol (VBe) | 206.63 ± 7.71 | 118.47 ± 8.72 ** | 1.74 | 80.03 ± 11.59 ** | 2.58 | 102.45 ± 5.75 ** | 2.02 |
| Bark—infusion (VBi) | >500 | 296.85 ± 21.71 | >1.68 | >500 | na | 306.20 ± 9.19 | >1.63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakar, K.; Nilofar; Mohamed, A.; Świątek, Ł.; Hryć, B.; Sieniawska, E.; Rajtar, B.; Ferrante, C.; Menghini, L.; Zengin, G.; et al. Evaluating Phytochemical Profiles, Cytotoxicity, Antiviral Activity, Antioxidant Potential, and Enzyme Inhibition of Vepris boiviniana Extracts. Molecules 2023, 28, 7531. https://doi.org/10.3390/molecules28227531
Bakar K, Nilofar, Mohamed A, Świątek Ł, Hryć B, Sieniawska E, Rajtar B, Ferrante C, Menghini L, Zengin G, et al. Evaluating Phytochemical Profiles, Cytotoxicity, Antiviral Activity, Antioxidant Potential, and Enzyme Inhibition of Vepris boiviniana Extracts. Molecules. 2023; 28(22):7531. https://doi.org/10.3390/molecules28227531
Chicago/Turabian StyleBakar, Kassim, Nilofar, Andilyat Mohamed, Łukasz Świątek, Benita Hryć, Elwira Sieniawska, Barbara Rajtar, Claudio Ferrante, Luigi Menghini, Gokhan Zengin, and et al. 2023. "Evaluating Phytochemical Profiles, Cytotoxicity, Antiviral Activity, Antioxidant Potential, and Enzyme Inhibition of Vepris boiviniana Extracts" Molecules 28, no. 22: 7531. https://doi.org/10.3390/molecules28227531
APA StyleBakar, K., Nilofar, Mohamed, A., Świątek, Ł., Hryć, B., Sieniawska, E., Rajtar, B., Ferrante, C., Menghini, L., Zengin, G., & Polz-Dacewicz, M. (2023). Evaluating Phytochemical Profiles, Cytotoxicity, Antiviral Activity, Antioxidant Potential, and Enzyme Inhibition of Vepris boiviniana Extracts. Molecules, 28(22), 7531. https://doi.org/10.3390/molecules28227531