
Kinase Signaling in Apoptosis Induced by Saturated Fatty Acids in Pancreatic β-Cells


Abstract
:
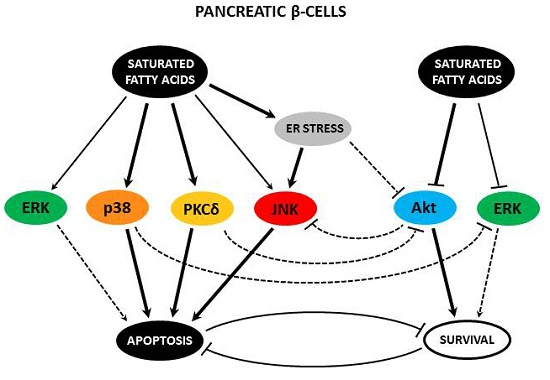{kind=link}
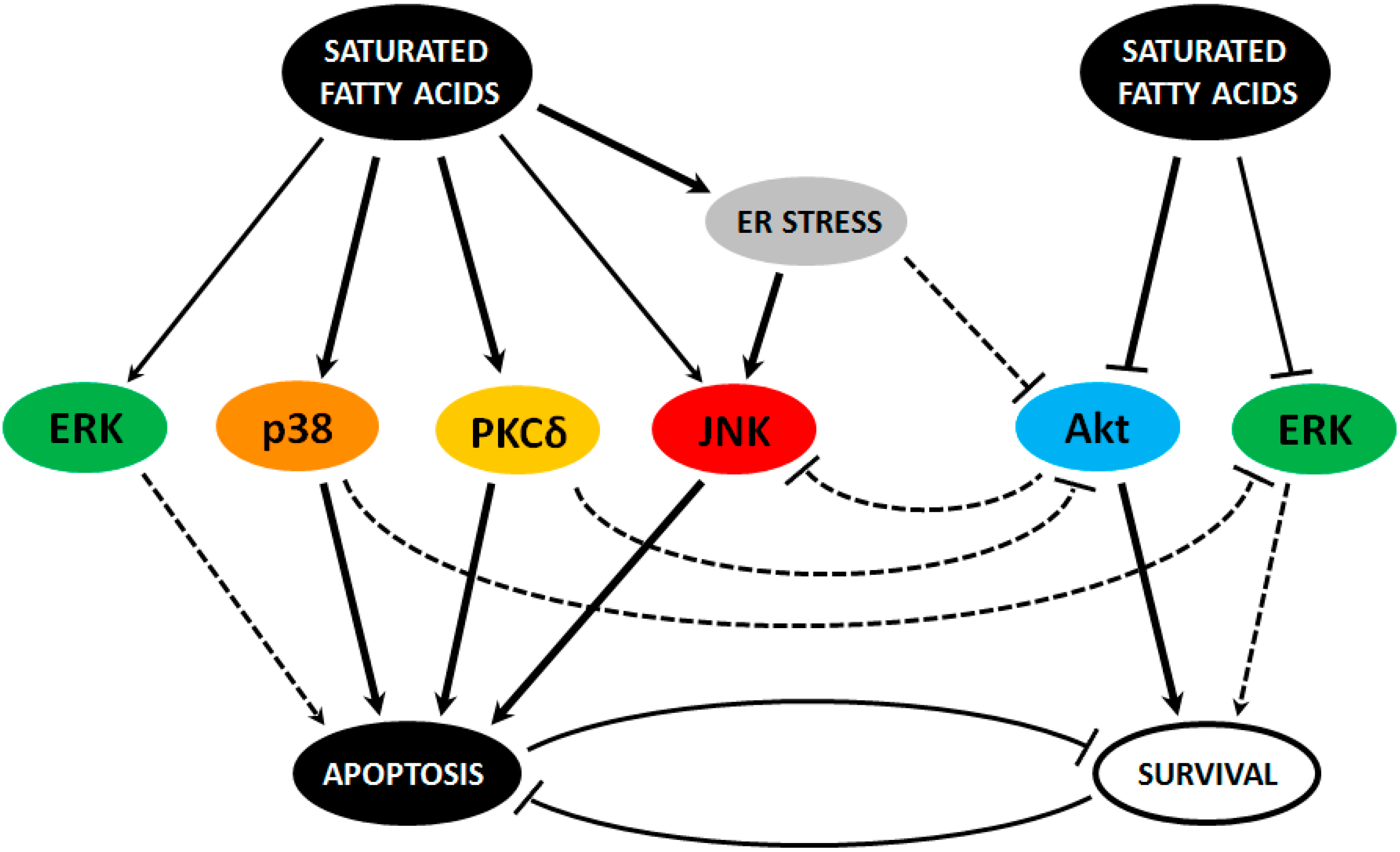{kind=link}
1. Introduction
2. c-Jun N-Terminal Kinase (JNK)
2.1. JNK and Its Role in Cell Signaling
2.2. JNK in Apoptosis Induced by Saturated Fatty Acids in β-Cells
3. Protein Kinase C (PKC)
3.1. PKC and Its Role in Cell Signaling
3.2. PKC in Apoptosis Induced by Saturated Fatty Acids in β-Cells
4. p38 Mitogen-Activated Protein Kinase (p38 MAPK)
4.1. p38 MAPK and Its Role in Cell Signaling
4.2. p38 MAPK in Apoptosis Induced by Saturated Fatty Acids in β-Cells
5. Extracellular Signal-Regulated Kinase 1/2 (ERK1/2)
5.1. ERK1/2 and Its Role in Cell Signaling
5.2. ERK1/2 in Apoptosis Induced by Saturated Fatty Acids in β-Cells
6. Akt (PKB, Protein Kinase B)
6.1. Akt and Its Role in Cell Signaling
6.2. Akt in Apoptosis Induced by Saturated Fatty Acids in β-Cells
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ASK1 | apoptosis signal-regulating kinase 1 |
| ATF2 | activating transcription factor 2 |
| ATF6 | activating transcription factor 6 |
| BAD | Bcl-2-associated death promoter |
| Bcl-2 | B-cell lymphoma 2 |
| BiP | immunoglobulin heavy chain-binding protein |
| Cdc42 | cell division control protein 42 |
| CHOP | CCAAT-enhancer-binding protein C/EBP |
| CSF-1 | colony stimulating factor 1 |
| DAG | diacylglycerol |
| eIF-4E | eukaryotic translation initiation factor 4E |
| eNOS | endothelial nitric oxide synthase |
| ER | endoplasmic reticulum |
| ERK1/2 | extracellular signal-regulated kinase 1/2 |
| FA | fatty acid |
| Foxo | forkhead box |
| IL-1β | interleukin 1β |
| IRE1α | inositol-requiring protein 1α |
| JNK | c-Jun N-terminal kinase |
| MAPK | mitogen-activated protein kinase |
| MAPKAPK | MAP kinase-activated protein kinase |
| MEK1/2 | mitogen-activated protein kinase/ERK kinase |
| MKK | mitogen-activated protein kinase kinase |
| MLKs | mixed-lineage kinases |
| MNKs | MAP kinase signal-integrating kinases |
| MSK1/2 | mitogen- and stress-activated protein kinase 1/2 |
| mTOR | mammalian target of rapamycin |
| NF-κB | nuclear factor κB |
| PAK | p21-activated protein kinase |
| PERK | protein kinase RNA (PKR)-like ER kinase |
| PIP2 | phosphatidylinositol 4,5-bisphosphate |
| PI3K | phosphoinositide 3-kinase |
| PDK1 | phosphoinositide-dependent protein kinase 1 |
| PKB | protein kinase B (also known as Akt kinase) |
| PKC | protein kinase C |
| p90RSK | p90 ribosomal S6 kinase |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| Raf | rapidly accelerated fibrosarcoma |
| Ras | rat sarcoma |
| RTKs | receptor tyrosine kinases |
| SAPK | stress-activated protein kinase |
| STAT | signal transducers and activators of transcription |
| Syk | spleen tyrosine kinase |
| TAK1 | transforming growth factor-β-activated kinase 1 |
| TCDD | 2,3,7,8-tetrachlorodibenzo-p-dioxin |
| TNF-α | tumor necrosis factor α |
| TPL2 | tumor progression locus 2 |
References
- Lupi, R.; Dotta, F.; Marselli, L.; del Guerra, S.; Masini, M.; Santangelo, C.; Patane, G.; Boggi, U.; Piro, S.; Anello, M.; et al. Prolonged exposure to free fatty acids has cytostatic and pro-apoptotic effects on human pancreatic islets: Evidence that β-cell death is caspase mediated, partially dependent on ceramide pathway, and Bcl-2 regulated. Diabetes 2002, 51, 1437–1442. [Google Scholar] [CrossRef] [PubMed]
- Maedler, K.; Oberholzer, J.; Bucher, P.; Spinas, G.A.; Donath, M.Y. Monounsaturated fatty acids prevent the deleterious effects of palmitate and high glucose on human pancreatic β-cell turnover and function. Diabetes 2003, 52, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Azevedo-Martins, A.K.; Monteiro, A.P.; Lima, C.L.; Lenzen, S.; Curi, R. Fatty acid-induced toxicity and neutral lipid accumulation in insulin-producing RINm5F cells. Toxicol. In Vitro 2006, 20, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Welters, H.J.; Diakogiannaki, E.; Mordue, J.M.; Tadayyon, M.; Smith, S.A.; Morgan, N.G. Differential protective effects of palmitoleic acid and cAMP on caspase activation and cell viability in pancreatic β-cells exposed to palmitate. Apoptosis 2006, 11, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Fürstova, V.; Kopska, T.; James, R.F.; Kovar, J. Comparison of the effect of individual saturated and unsaturated fatty acids on cell growth and death induction in the human pancreatic β-cell line NES2Y. Life Sci. 2008, 82, 684–691. [Google Scholar] [CrossRef]
- Maedler, K.; Spinas, G.A.; Dyntar, D.; Moritz, W.; Kaiser, N.; Donath, M.Y. Distinct effects of saturated and monounsaturated fatty acids on β-cell turnover and function. Diabetes 2001, 50, 69–76. [Google Scholar] [CrossRef] [PubMed]
- El-Assaad, W.; Buteau, J.; Peyot, M.L.; Nolan, C.; Roduit, R.; Hardy, S.; Joly, E.; Dbaibo, G.; Rosenberg, L.; Prentki, M. Saturated fatty acids synergize with elevated glucose to cause pancreatic β-cell death. Endocrinology 2003, 144, 4154–4163. [Google Scholar]
- Kharroubi, I.; Ladriere, L.; Cardozo, A.K.; Dogusan, Z.; Cnop, M.; Eizirik, D.L. Free fatty acids and cytokines induce pancreatic β-cell apoptosis by different mechanisms: role of nuclear factor-κB and endoplasmic reticulum stress. Endocrinology 2004, 145, 5087–5096. [Google Scholar] [CrossRef] [PubMed]
- Welters, H.J.; Tadayyon, M.; Scarpello, J.H.; Smith, S.A.; Morgan, N.G. Mono-unsaturated fatty acids protect against β-cell apoptosis induced by saturated fatty acids, serum withdrawal or cytokine exposure. FEBS Lett. 2004, 560, 103–108. [Google Scholar] [CrossRef]
- Higa, M.; Shimabukuro, M.; Shimajiri, Y.; Takasu, N.; Shinjyo, T.; Inaba, T. Protein kinase B/Akt signaling is required for palmitate-induced β-cell lipotoxicity. Diabetes Obes. Metab. 2006, 8, 228–233. [Google Scholar] [CrossRef]
- Karaskov, E.; Scott, C.; Zhang, L.; Teodoro, T.; Ravazzola, M.; Volchuk, A. Chronic palmitate but not oleate exposure induces endoplasmic reticulum stress, which may contribute to INS-1 pancreatic β-cell apoptosis. Endocrinology 2006, 147, 3398–3407. [Google Scholar] [CrossRef] [PubMed]
- Hennige, A.M.; Ranta, F.; Heinzelmann, I.; Dufer, M.; Michael, D.; Braumuller, H.; Lutz, S.Z.; Lammers, R.; Drews, G.; Bosch, F.; et al. Overexpression of kinase-negative protein kinase Cδ in pancreatic β-cells protects mice from diet-induced glucose intolerance and β-cell dysfunction. Diabetes 2010, 59, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Eitel, K.; Staiger, H.; Brendel, M.D.; Brandhorst, D.; Bretzel, R.G.; Haring, H.U.; Kellerer, M. Different role of saturated and unsaturated fatty acids in β-cell apoptosis. Biochem. Biophys. Res. Commun. 2002, 299, 853–856. [Google Scholar] [CrossRef]
- Sone, H.; Kagawa, Y. Pancreatic β cell senescence contributes to the pathogenesis of type 2 diabetes in high-fat diet-induced diabetic mice. Diabetologia 2005, 48, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Diakogiannaki, E.; Welters, H.J.; Morgan, N.G. Differential regulation of the endoplasmic reticulum stress response in pancreatic β-cells exposed to long-chain saturated and monounsaturated fatty acids. J. Endocrinol. 2008, 197, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Nemcova-Fürstova, V.; James, R.F.L.; Kovar, J. Inhibitory effect of unsaturated fatty acids on saturated fatty acids-induced apoptosis in human pancreatic β-cells: Activation of caspases and ER stress induction. Cell. Physiol. Biochem. 2011, 27, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Martinez, S.C.; Tanabe, K.; Cras-Méneur, C.; Abumrad, N.A.; Bernal-Mizrachi, E.; Permutt, M.A. Inhibition of FoxO1 protects pancreatic islet β-cells against fatty acid and endoplasmic reticulum stress-induced apoptosis. Diabetes 2008, 57, 846–859. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Cai, K.; Wang, O.; Li, Z.; Chen, J.; Deng, B.; Qian, L.; Le, Y. Fatty acids induce amylin expression and secretion by pancreatic β-cells. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E99–E107. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zhang, X.; Huang, X.; Lu, Y.; Tang, W.; Man, Y.; Wang, S.; Xi, J.; Li, J. NADPH Oxidase 2-Derived Reactive Oxygen Species Mediate FFAs-Induced Dysfunction and Apoptosis of β-Cells via JNK, p38 MAPK and p53 Pathways. PLoS ONE 2010, 5, e15726. [Google Scholar] [CrossRef] [PubMed]
- Biden, T.J.; Boslem, E.; Chu, K.Y.; Sue, N. Lipotoxic endoplasmic reticulum stress, β cell failure, and type 2 diabetes mellitus. Trends Endocrinol. Metab. 2014, 25, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Cvjeticanin, T.; Stojanovic, I.; Timotijevic, G.; Stosic-Grujicic, S.; Miljkovic, D. T cells cooperate with palmitic acid in induction of β cell apoptosis. BMC Immunol. 2009, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jung, I.R.; Choi, S.E.; Lee, S.M.; Lee, S.J.; Han, S.J.; Kim, H.J.; Kim, D.J.; Lee, K.W.; Kang, Y. Toxicity generated through inhibition of pyruvate carboxylase and carnitine palmitoyl transferase-1 is similar to high glucose/palmitate-induced glucolipotoxicity in INS-1 β cells. Mol. Cell. Endocrinol. 2014, 383, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Fang, N.; Lou, J.; Zhang, W.; Xu, S.; Liu, H.; Fang, Q.; Wang, Z.; Liu, J.; Men, X.; et al. TRB3 Is Involved in Free Fatty Acid-Induced INS-1-Derived Cell Apoptosis via the Protein Kinase C δ Pathway. PLoS ONE 2014, 9, e96089. [Google Scholar] [CrossRef] [PubMed]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Blenis, J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, M.; Zhang, S.; Yan, L.; Yang, C.; Lu, W.; Li, Y.; Cheng, H. The role of G protein-coupled receptor 40 in lipoapoptosis in mouse β-cell line NIT-1. Mol. Cell. Endocrinol. 2007, 38, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Quan, X.; Zhang, L.; Li, Y.; Liang, C. TCF2 attenuates FFA-induced damage in islet β-cells by regulating production of insulin and ROS. Int. J. Mol. Sci. 2014, 15, 13317–13332. [Google Scholar] [CrossRef] [PubMed]
- Wrede, C.E.; Dickson, L.M.; Lingohr, M.K.; Briaud, I.; Rhodes, C.J. Fatty acid and phorbol ester-mediated interference of mitogenic signaling via novel protein kinase C isoforms in pancreatic β-cells (INS-1). J. Mol. Endocrinol. 2003, 30, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Avruch, J. pp54 Microtubule-associated protein 2 kinase: A novel serine/threonine protein kinase regulated by phosphorylation and stimulated by poly-L-lysine. J. Biol. Chem. 1990, 265, 17355–17363. [Google Scholar] [PubMed]
- Kyriakis, J.M.; Brautigan, D.L.; Ingebritsen, T.S.; Avruch, J. pp54 microtubuleassociated protein-2 kinase requires both tyrosine and threonine phosphorylation for activity. J. Biol. Chem. 1991, 266, 10043–10046. [Google Scholar] [PubMed]
- Derijard, B.; Hibi, M.; Wu, I.-H.; Barrett, T.; Su, B.; Deng, T.; Karin, M.; Davis, R.J. JNK1: A protein kinase stimulated by UV light and Ha-Ras that binds and phosphorylates the c-Jun activation domain. Cell 1994, 76, 1025–1037. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Banerjee, P.; Nikolakaki, E.; Dai, T.; Rubie, E.A.; Ahmad, M.F.; Avruch, J.; Woodgett, J.R. The stress-activated protein kinase subfamily of c-Jun kinases. Nature 1994, 369, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Kallunki, T.; Su, B.; Tsigelny, I.; Sluss, H.K.; Derijard, B.; Moore, G.; Davis, R.; Karin, M. JNK2 contains a specificity-determining region responsible for efficient C-Jun binding and phosphorylation. Genes Dev. 1994, 8, 2996–3007. [Google Scholar] [CrossRef]
- Davis, R.J. Signal transduction by the JNK group of MAP kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef]
- Hetz, C.; Martinon, F.; Rodriguez, D.; Glimcher, L.H. The unfolded protein response: Integrating stress signals through the stress sensor IRE1α. Physiol. Rev. 2011, 91, 1219–1243. [Google Scholar] [CrossRef] [PubMed]
- Bogoyevitch, M.A.; Kobe, B. Uses for JNK: The many and varied substrates of the c-Jun N-terminal kinases. Microbiol. Mol. Biol. Rev. 2006, 70, 1061–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natalicchio, A.; Labarbuta, R.; Tortosa, F.; Biondi, G.; Marrano, N.; Peschechera, A.; Carchia, E.; Orlando, M.R.; Leonardini, A.; Cignarelli, A.; et al. Exendin-4 protects pancreatic β cells from palmitate-induced apoptosis by interfering with GPR40 and the MKK4/7 stress kinase signalling pathway. Diabetologia 2013, 56, 2456–2466. [Google Scholar] [CrossRef] [PubMed]
- Bachar, E.; Ariav, Y.; Ketzinel-Gilad, M.; Cerasi, E.; Kaiser, N.; Leibowitz, G. Glucose amplifies fatty acid-induced endoplasmic reticulum stress in pancreatic β-cells via activation of mTORC1. PLoS ONE 2009, 4, e4954. [Google Scholar] [CrossRef] [PubMed]
- Cunha, D.A.; Hekerman, P.; Ladrière, L.; Bazarra-Castro, A.; Ortis, F.; Wakeham, M.C.; Moore, F.; Rasschaert, J.; Cardozo, A.K.; Bellomo, E.; et al. Initiation and execution of lipotoxic ER stress in pancreatic β-cells. J. Cell Sci. 2008, 121, 2308–2318. [Google Scholar] [CrossRef] [PubMed]
- Cunha, D.A.; Igoillo-Esteve, M.; Gurzov, E.N.; Germano, C.M.; Naamane, N.; Marhfour, I.; Fukaya, M.; Vanderwinden, J.-M.; Gysemans, C.; Mathieu, C.; et al. Death protein 5 and p53-upregulated modulator of apoptosis mediate the endoplasmic reticulum stress—mitochondrial dialog triggering lipotoxic rodent and human β-cell apoptosis. Diabetes 2012, 61, 2763–2775. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.S.; Lee, Y.J.; Kang, Y.; Han, J.; Lim, O.K.; Jun, H.S. Exendin-4 inhibits glucolipotoxic ER stress in pancreatic β cells via regulation of SREBP1c and C/EBPβ transcription factors. J. Endocrinol. 2013, 216, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Prause, M.; Christensen, D.P.; Billestrup, N.; Mandrup-Poulsen, T. JNK1 protects against glucolipotoxicity-mediated β-cell apoptosis. PLoS ONE 2014, 9, e87067. [Google Scholar]
- Abaraviciene, S.M.; Lundquist, I.; Salehi, A. Rosiglitazone counteracts palmitate-induced β-cell dysfunction by suppression of MAP kinase, inducible nitric oxide synthase and caspase 3 activities. Cell. Mol. Life Sci. 2008, 65, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Kim, D.-H.; Kim, Y.H. Compound K protects MIN6N8 pancreatic β-cells against palmitate-induced apoptosis through modulating SAPK/JNK activation. Cell Biol. Int. 2010, 34, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Aikin, R.; Maysinger, D.; Rosenberg, L. Cross-talk between phosphatidylinositol 3-kinase/AKT and c-jun NH2-terminal kinase mediates survival of isolated human islets. Endocrinology 2004, 145, 4522–4531. [Google Scholar] [CrossRef] [PubMed]
- Nemcova-Fürstova, V.; Balusikova, K.; Sramek, J.; James, R.F.; Kovar, J. Caspase-2 and JNK activated by saturated fatty acids are not involved in apoptosis induction but modulate ER stress in human pancreatic β-cells. Cell. Physiol. Biochem. 2013, 31, 277–289. [Google Scholar]
- Komiya, K.; Uchida, T.; Ueno, T.; Koike, M.; Abe, H.; Hirose, T.; Kawamori, R.; Uchiyama, Y.; Kominami, E.; Fujitani, Y.; et al. Free fatty acids stimulate autophagy in pancreatic β-cells via JNK pathway. Biochem. Biophys. Res. Commun. 2010, 401, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, Y.; Cazanave, S.; Mott, J.L.; Elmi, N.; Bronk, S.F.; Kohno, S.; Charlton, M.R.; Gores, G.J. Palmitoleate attenuates palmitate-induced Bim and PUMA up-regulation and hepatocyte lipoapoptosis. J. Hepatol. 2010, 52, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Gunaratnam, K.; Vidal, C.; Boadle, R.; Thekkedam, C.; Duque, G. Mechanisms of palmitate-induced cell death in human osteoblasts. Biol. Open 2013, 2, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Win, S.; Than, T.A.; Le, B.H.; Garcia-Ruiz, C.; Fernandez-Checa, J.C.; Kaplowitz, N. Sab (Sh3bp5) dependence of JNK mediated inhibition of mitochondrial respiration in palmitic acid induced hepatocyte lipotoxicity. J. Hepatol. 2015, 62, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Mellor, H.; Parker, P.J. The extended protein kinase C superfamily. Biochem. J. 1998, 332, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. Intracellular signaling by hydrolysis of phospholipids and activation of PKC. Science 1992, 258, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Kanashiro, C.A.; Khalil, R.A. Signal transduction by protein kinase C in mammalian cells. Clin. Exp. Pharmacol. Physiol. 1998, 25, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Reyland, M.E. Protein kinase Cdelta and apoptosis. Biochem. Soc. Trans. 2007, 35, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Xia, L.; Chen, G.Q. Protein kinase C δ in apoptosis: A brief overview. Arch. Immunol. Ther. Exp. 2012, 60, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Murriel, C.L.; Churchill, E.; Inagaki, K.; Szweda, L.I.; Mochly-Rosen, D. Protein kinase C delta activation induces apoptosis in response to cardiac ischemia and reperfusion damage: A mechanism involving BAD and the mitochondria. J. Biol. Chem. 2004, 279, 47985–47991. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hong, F.; Kwon, S.; Kim, S.S.; Kim, D.O.; Kang, H.S.; Lee, S.J.; Ha, J.; Kim, S.S. Activation of p38MAPK induces cell cycle arrest via inhibition of C-Raf/ERK pathway during muscle differentiation. Biochem. Biophys. Res. Commun. 2002, 298, 765–771. [Google Scholar] [CrossRef]
- Efimova, T.; Broome, A.M.; Eckert, R.L. Protein kinase C delta regulates keratinocyte death and survival by regulating activity and subcellular localization of a p38delta-extracellular signal regulated kinase 1/2 complex. Mol. Cell. Biol. 2004, 24, 8167–8183. [Google Scholar] [CrossRef] [PubMed]
- Eitel, K.; Staiger, H.; Rieger, J.; Mischak, H.; Brandhorst, H.; Brendel, M.D.; Bretzel, R.G.; Häring, H.U.; Kellerer, M. Protein kinase C delta activation and translocation to the nucleus are required for fatty acid-induced apoptosis of insulin-secreting cells. Diabetes 2003, 52, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.N.; Azevedo-Martins, A.K.; Amanso, A.M.; Carvalho, C.R.O.; Curi, R. Persistent activation of Akt or ERK prevents the toxicity induced by saturated and polyunsaturated fatty acids in RINm5F β-cells. Toxicol. In Vitro 2008, 22, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Alcáazar, O.; Qiu-yue, Z.; Gine, E.; Tamarit-Rodriguez, J. Stimulation of islet protein kinase C translocation by palmitate requires metabolism of the fatty acid. Diabetes 1997, 46, 1153–1158. [Google Scholar] [CrossRef]
- Carpenter, L.; Cordery, D.; Biden, T.J. Inhibition of protein kinase C protects rat INS-1 cells against interleukin-1β and streptozotocin-induced apoptosis. Diabetes 2002, 51, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Denning, M.F.; Wang, Y.; Nickoloff, B.J.; Wrone-Smith, T. Protein kinase C δ is activated by caspase-dependent proteolysis during ultraviolet radiation-induced apoptosis of human keratinocytes. J. Biol. Chem. 1998, 273, 29995–30002. [Google Scholar] [CrossRef] [PubMed]
- Reyland, M.E.; Anderson, S.M.; Matassa, A.A.; Barzen, K.A.; Quissell, D.O. Protein kinase C delta is essential for etoposide-induced apoptosis in salivary gland acinar cells. J. Biol. Chem. 1999, 274, 19115–19123. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Hur, E.M.; Lee, J.H.; Jun, D.J.; Kim, K.T. Protein kinase C δ-mediated proteasomal degradation of MAP kinase phosphatase-1 contributes to glutamate-induced neuronal cell death. J. Cell Sci. 2006, 119, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, C.; Li, Z.; Guo, W.; Gegner, J.A.; Lin, S.; Han, J. Characterization of the structure and function of a new mitogen-activated protein kinase (p38b). J. Biol. Chem. 1996, 271, 17920–17926. [Google Scholar] [CrossRef] [PubMed]
- Lechner, C.; Zahalka, M.A.; Giot, J.F.; Moler, N.P.H.; Ullrich, A. ERK6, a mitogen-activated protein kinase involved in C2C12 myoblast differentiation. Proc. Natl. Acad. Sci. USA 1996, 93, 4355–4359. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, Y.; Ulevitch, R.J.; Han, J. The primary structure of p38γ: A new member of p38 group of MAP kinase. Biochem. Biophys. Res. Commun. 1996, 228, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Gram, H.; Zhao, M.; New, L.; Gu, J.; Feng, L.; Di Padova, F.; Ulevitch, R.J.; Han, J. Characterization of the structure and function of the fourth member of p38 group mitogen activated protein kinases, p38δ. J. Biol. Chem. 1997, 272, 30122–30128. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; McDonnell, P.C.; Gum, R.J.; Hand, A.T.; Lee, J.C.; Young, P.R. Novel homologues of CSBP/p38 MAP kinase: activation, substrate specificity and sensitivity to inhibition by pyridinyl imidazoles. Biochem. Biophys. Res. Commun. 1997, 235, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, T.; Kuroyanagi, N.; Yamaguchi, K.; Gotoh, Y.; Irie, K.; Kano, T.; Shirakabe, K.; Muro, Y.; Shibuya, H.; Matsumoto, K.; et al. A novel kinase cascade mediated by mitogen-activated protein kinase kinase 6 and MKK3. J. Biol. Chem. 1996, 271, 13675–13679. [Google Scholar] [PubMed]
- Ichijo, H.; Nishida, E.; Irie, K.; ten Dijke, P.; Saitoh, M.; Moriguchi, T.; Takagi, M.; Matsumoto, K.; Miyazono, K.; Gotoh, Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science 1997, 275, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Han, J.; Sells, M.A.; Chernoff, J.; Knaus, U.G.; Ulevitch, R.J.; Bokoch, G.M. Rho family GTPases regulate p38 MAP kinase through the downstream mediator Pak1. J. Biol. Chem. 1995, 270, 23934–23936. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Dorow, D.S. Differential activation of stress-activated protein kinase kinases SKK4/MKK7 and SKK1/MKK4 by the mixed-lineage kinase-2 and mitogen-activated protein kinase kinase (MKK) kinase-1. Biochem. J. 1998, 333, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Takekawa, M.; Posas, F.; Saito, H. A human homolog of the yeast Ssk2/Ssk22 MAP kinase kinase kinases, MTK1, mediates stress induced activation of the p38 and JNK pathways. EMBO J. 1997, 16, 4973–4982. [Google Scholar] [CrossRef] [PubMed]
- Bagrodia, S.; Derijard, B.; Davis, R.J.; Cerione, R.A. Cdc42 and PAK mediated signaling leads to Jun kinase and p38 mitogen-activated protein kinase activation. J. Biol. Chem. 1995, 270, 27995–27998. [Google Scholar] [PubMed]
- Zarubin, T.; Han, J. Activation and signaling of the p38 MAP kinase pathway. Cell Res. 2005, 15, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2004, 1773, 1358–1375. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chang, F.; Li, F.; Fu, H.; Wang, J.; Zhang, S.; Zhao, J.; Yin, D. Palmitate promotes autophagy and apoptosis through ROS-dependent JNK and p38 MAPK. Biochem. Biophys. Res. Commun. 2015, 463, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Sramek, J.; Nemcova-Fürstova, V.; Balusikova, K.; Daniel, P.; Jelinek, M.; James, R.F.; Kovar, J. p38 MAPK is activated but does not play a key role during apoptosis induction by saturated fatty acid in human pancreatic β-cells. Int. J. Mol. Sci. 2016, 17, 159. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yang, W.; Wu, L.; Wang, R. H2S, endoplasmic reticulum stress, and apoptosis of insulin-secreting β cells. J. Biol. Chem. 2007, 282, 16567–16576. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Nunemaker, C.S.; Wu, R.; Chakrabarti, S.K.; Taylor-Fishwick, D.A.; Nadler, J.L. 12-Lipoxygenase products reduce insulin secretion and β-cell viability in human islets. J. Clin. Endocrinol. Metab. 2010, 95, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Makeeva, N.; Myers, J.W.; Welsh, N. Role of MKK3 and p38 MAPK in cytokine-induced death of insulin-producing cells. Biochem. J. 2006, 393, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Saldeen, J.; Lee, J.C.; Welsh, N. Role of p38 mitogen-activated protein kinase (p38 MAPK) in cytokine-induced rat islet cell apoptosis. Biochem. Pharmacol. 2001, 61, 1561–1569. [Google Scholar] [CrossRef]
- Chai, W.; Liu, Z. p38 mitogen-activated protein kinase mediates palmitate-induced apoptosis but not inhibitor of nuclear factor-kappa B degradation in human coronary artery endothelial cells. Endocrinology 2007, 148, 1622–1628. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cao, W. p38 mitogen-activated protein kinase: a critical node linking insulin resistance and cardiovascular diseases in type 2 diabetes mellitus. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, X.; Li, Y.; Li, N.; Shi, X.; Ding, H.; Zhang, Y.; Li, X.; Liu, G.; Wang, Z. Non-esterified fatty acids activate the ROS-p38-p53/Nrf2 signaling pathway to induce bovine hepatocyte apoptosis in vitro. Apoptosis 2014, 19, 984–997. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, V.; Grasso, S.; Tibullo, D.; Giallongo, C.; Raciti, G.; Viola, M.; Avola, R. Modulation of extracellular signal-related kinase, cyclin D1, glial fibrillary acidic protein, and vimentin expression in estradiol-pretreated astrocyte cultures treated with competence and progression growth factors. J. Neurosci. Res. 2015, 93, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Gudermann, T. Multiple pathways of ERK activation by G protein-coupled receptors. Novartis Found. Symp. 2001, 239, 68–79. [Google Scholar] [PubMed]
- Chang, F.; Steelman, L.S.; Lee, J.T.; Shelton, J.G.; Navolanic, P.M.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Signal transduction mediated by the Ras/Raf/MEK/ERK pathway from cytokine receptors to transcription factors: Potential targeting for therapeutic intervention. Leukemia 2003, 17, 1263–1293. [Google Scholar] [CrossRef] [PubMed]
- Kolch, W.; Heidecker, G.; Kochs, G.; Hummel, R.; Vahidi, H.; Mischak, H.; Finkenzeller, G.; Marme, D.; Rapp, U.R. Protein kinase Ca activates RAF-1 by direct phosphorylation. Nature 1993, 364, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Fabian, J.R.; Daar, I.O.; Morrison, D.K. Critical tyrosine residues regulate the enzymatic and biological activity of Raf-1 kinase. Mol. Cell. Biol. 1993, 13, 7170–7179. [Google Scholar] [CrossRef] [PubMed]
- Diaz, B.; Barnard, D.; Filson, A.; MacDonald, S.; King, A.; Marshall, M. Phosphorylation of Raf-1 serine 338-serine 339 is an essential regulatory event for Ras-dependent activation and biological signaling. Mol. Cell. Biol. 1997, 17, 4509–4516. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gibson, T.B.; Robinson, F.; Silvestro, L.; Pearson, G.; Xu, B.; Wright, A.; Vanderbilt, C.; Cobb, M.H. MAP Kinases. Chem. Rev. 2001, 101, 2449–2476. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, V.; Grasso, S.; Tibullo, D.; Giallongo, C.; Pappa, R.; Brundo, M.V.; Tomassoni, D.; Viola, M.; Amenta, F.; Avola, R. Neuroactive molecules and growth factors modulate cytoskeletal protein expression during astroglial cell proliferation and differentiation in culture. J. Neurosci. Res. 2016, 94, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Cagnol, S.; Chambard, J.C. ERK and cell death: Mechanisms of ERK-induced cell death—Apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Qian, Y.Y.; Xi, X.X.; Hu, X.H.; Zhu, J.X.; Han, X. Blockage of ceramide metabolism exacerbates palmitate inhibition of pro-insulin gene expression in pancreatic β-cells. Mol. Cell. Biochem. 2010, 338, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Plaisance, V.; Perret, V.; Favrea, D.; Abderrahmania, A.; Yanga, J.-Y.; Widmanna, C.; Regazzi, R. Role of the transcriptional factor C/EBP in free fatty acid-elicited β cell failure. Mol. Cell. Endocrinol. 2009, 305, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Fontes, G.; Semache, M.; Hagman, D.K.; Tremblay, C.; Shah, R.; Rhodes, C.J.; Rutter, J.; Poitout, V. Involvement of Per-Arnt-Sim Kinase and extracellular-regulated kinases-1/2 in palmitate inhibition of insulin gene expression in pancreatic β-cells. Diabetes 2009, 58, 2048–2058. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.L.; Macrae, K.; Marley, A.E.; Hundal, H.S. Chronic effects of palmitate overload on nutrient-induced insulin secretion and autocrine signalling in pancreatic MIN6 β cells. PLoS ONE 2011, 6, e25975. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.M.; Wadt, K.A.; Juhl, L.F.; Andersen, H.U.; Karlsen, A.E.; Su, M.S.; Seedorf, K.; Shapiro, L.; Dinarello, C.A.; Mandrup-Poulsen, T. Interleukin-1β-induced rat pancreatic islet nitric oxide synthesis requires both the p38 and extracellular signal-regulated kinase 1/2 mitogen-activated protein kinases. J. Biol. Chem. 1998, 273, 15294–15300. [Google Scholar] [CrossRef] [PubMed]
- Maedler, K.; Storling, J.; Sturis, J.; Zuellig, R.A.; Spinas, G.A.; Arkhammar, P.O.; Mandrup-Poulsen, T.; Donath, M.Y. Glucose- and interleukin-1β-induced β-cell apoptosis requires Ca2+ influx and extracellular signal-regulated kinase (ERK) 1/2 activation and is prevented by a sulfonylurea receptor 1/inwardly rectifying K+ channel 6.2 (SUR/Kir6.2) selective potassium channel opener in human islets. Diabetes 2004, 53, 1706–1713. [Google Scholar] [PubMed]
- Lawrence, M.C.; McGlynn, K.; Park, B.H.; Cobb, M.H. ERK1/2-dependent activation of transcription factors required for acute and chronic effects of glucose on the insulin gene promoter. J. Biol. Chem. 2005, 280, 26751–26759. [Google Scholar] [CrossRef] [PubMed]
- Martino, L.; Masini, M.; Novelli, M.; Giacopelli, D.; Beffy, P.; Masiello, P.; de Tata, V. The aryl receptor inhibitor epigallocatechin-3-gallate protects INS-1E β-cell line against acute dioxin toxicity. Chemosphere 2013, 93, 1447–1455. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Wei, F.Y.; Matsunaga, T.; Matsunaga, N.; Kaitsuka, T.; Tomizawa, K. Oxytocin protects against stress-induced cell death in murine pancreatic β-cells. Sci. Rep. 2016, 6, 25185. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Yoder, S.M.; Wang, Z.; Oh, E.; Ramalingam, L.; Tunduguru, R.; Thurmond, D.C. The p21-activated kinase (PAK1) is involved in diet-induced β cell mass expansion and survival in mice and human islets. Diabetologia 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Ahn, M.W.; Baek, S.H.; Lee, I.K.; Kim, Y.W.; Kim, J.Y.; Dan, J.M.; Park, S.Y. AMPK activator, AICAR, inhibits palmitate-induced apoptosis in osteoblast. Bone 2008, 43, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Bi, L.; He, S.; Meng, G.; Wei, B.; Jia, S.; Liu, J. FFAs-ROS-ERK/P38 pathway plays a key role in adipocyte lipotoxicity on osteoblasts in co-culture. Biochimie 2014, 101, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Sun, C.; Gu, W.; Yang, M.; Zhang, X.; Zhai, N.; Lu, Y.; Zhang, Z.; Shou, P.; Zhang, Z.; Ning, G. Free fatty acid receptor 2, a candidate target for type 1 diabetes, induces cell apoptosis through ERK signaling. J. Mol. Endocrinol. 2014, 53, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Hanada, M.; Feng, J.; Hemmings, B.A. Structure, regulation and function of PKB/AKT—A major therapeutic target. Biochim. Biophys. Acta 2004, 1697, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Morales, B.M.; de Plata, A.C. Role of AKT/mTORC1 pathway in pancreatic β-cell proliferation. Colomb. Med. 2012, 43, 235–243. [Google Scholar]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—In control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Winter, K.; Nian, C.; Tsuneoka, M.; Koda, Y.; McIntosh, C.H.S. Glucose-dependent insulinotropic polypeptide (GIP) stimulation of pancreatic Β cell survival is dependent upon phosphatidylinositol3-kinase (PI3K)/protein kinase B (PKB) signaling, inactivation of the forkhead transcription factor FoxO1, and down-regulation of bax expression. J. Biol. Chem. 2005, 280, 22297–22307. [Google Scholar] [PubMed]
- Del Peso, L.; Gonzalez-Garcia, M.; Page, C.; Herrera, R.; Nunez, G. Interleukin-3-induced phosphorylation of BAD through the protein kinase Akt. Science 1997, 278, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Ozes, O.N.; Mayo, L.D.; Gustin, J.A.; Pfeffer, S.R.; Pfeffer, L.M.; Donner, D.B. NF-κB activation by tumour necrosis factor requires the Akt serine-threonine kinase. Nature 1999, 401, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti-Carvalho, J.E.; Nogueira, T.C.; Gorjão, R.; Bromati, C.R.; Yamanaka, T.S.; Boschero, A.C.; Velloso, L.A.; Curi, R.; Anhê, G.F.; Bordin, S. UPR-mediated TRIB3 expression correlates with reduced AKT phosphorylation and inability of interleukin 6 to overcome palmitate-induced apoptosis in RINm5F cells. J. Endocrinol. 2010, 206, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Guo, Q.H.; Wang, X.; Xue, B.; Sun, L.Q.; Meng, Q.T.; Lu, J.M.; Mu, Y.M. LRP16 gene protects mouse insulinoma MIN6 cells against fatty acid-induced apoptosis through Akt/FoxO1 signaling. Chin. Med. J. 2012, 125, 1695–1702. [Google Scholar] [PubMed]
- Shao, S.; Nie, M.; Chen, C.; Chen, X.; Zhang, M.; Yuan, G.; Yu, X.; Yang, Y. Protective action of liraglutide in β cells under lipotoxic stress via PI3K/Akt/FoxO1 pathway. J. Cell. Biochem. 2014, 115, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Kang, J.; Cao, Y.; Fan, S.; Yang, H.; An, Y.; Pan, Y.; Tie, L.; Li, X. Curcumin attenuates palmitate-induced apoptosis in MIN6 pancreatic β-cells through PI3K/Akt/FoxO1 and mitochondrial survival pathways. Apoptosis 2015, 20, 1420–1432. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wang, Z.; Tao, L.; Wang, Y. ER stress negatively regulates AKT/TSC/mTOR pathway to enhance autophagy. Autophagy 2010, 6, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Hiramatsu, N.; Hayakawa, K.; Saito, Y.; Kato, H.; Huang, T.; Yao, J.; Paton, A.W.; Paton, J.C.; Kitamura, M. Selective abrogation of BiP/GRP78 blunts activation of NF-κB through the ATF6 branch of the UPR: Involvement of C/EBPβ and mTOR-dependent dephosphorylation of Akt. Mol. Cell. Biol. 2011, 31, 1710–1718. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, R.L.; Gill, N.S.; Pugh, W.; Lee, J.P.; Koeberlein, B.; Furth, E.E.; Polonsky, K.S.; Naji, A.; Birnbaum, M.J. Regulation of pancreatic β-cell growth and survival by the serine/threonine protein kinase Akt1/PKBalpha. Nat. Med. 2001, 7, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Jeong, S.; Jing, K.; Shin, S.; Kim, S.; Heo, J.Y.; Kweon, G.R.; Park, S.K.; Wu, T.; Park, J.I.; et al. Docosahexaenoic acid induces cell death in human non-small cell lung cancer cells by repressing mTOR via AMPK activation and PI3K/Akt inhibition. BioMed Res. Int. 2015, 2015, 239764. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhou, H.; Tao, Y.; Guo, J.; Guo, Z.; Zhang, S.; Zhang, Y.; Huang, Y.; Tang, Y.; Dong, Q.; et al. G-CSF protects human brain vascular endothelial cells injury induced by high glucose, free fatty acids and hypoxia through MAPK and Akt signaling. PLoS ONE 2015, 10, e0120707. [Google Scholar]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šrámek, J.; Němcová-Fürstová, V.; Kovář, J. Kinase Signaling in Apoptosis Induced by Saturated Fatty Acids in Pancreatic β-Cells. Int. J. Mol. Sci. 2016, 17, 1400. https://doi.org/10.3390/ijms17091400
Šrámek J, Němcová-Fürstová V, Kovář J. Kinase Signaling in Apoptosis Induced by Saturated Fatty Acids in Pancreatic β-Cells. International Journal of Molecular Sciences. 2016; 17(9):1400. https://doi.org/10.3390/ijms17091400
Chicago/Turabian StyleŠrámek, Jan, Vlasta Němcová-Fürstová, and Jan Kovář. 2016. "Kinase Signaling in Apoptosis Induced by Saturated Fatty Acids in Pancreatic β-Cells" International Journal of Molecular Sciences 17, no. 9: 1400. https://doi.org/10.3390/ijms17091400
APA StyleŠrámek, J., Němcová-Fürstová, V., & Kovář, J. (2016). Kinase Signaling in Apoptosis Induced by Saturated Fatty Acids in Pancreatic β-Cells. International Journal of Molecular Sciences, 17(9), 1400. https://doi.org/10.3390/ijms17091400