Glycosaminoglycan-Mediated Downstream Signaling of CXCL8 Binding to Endothelial Cells

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
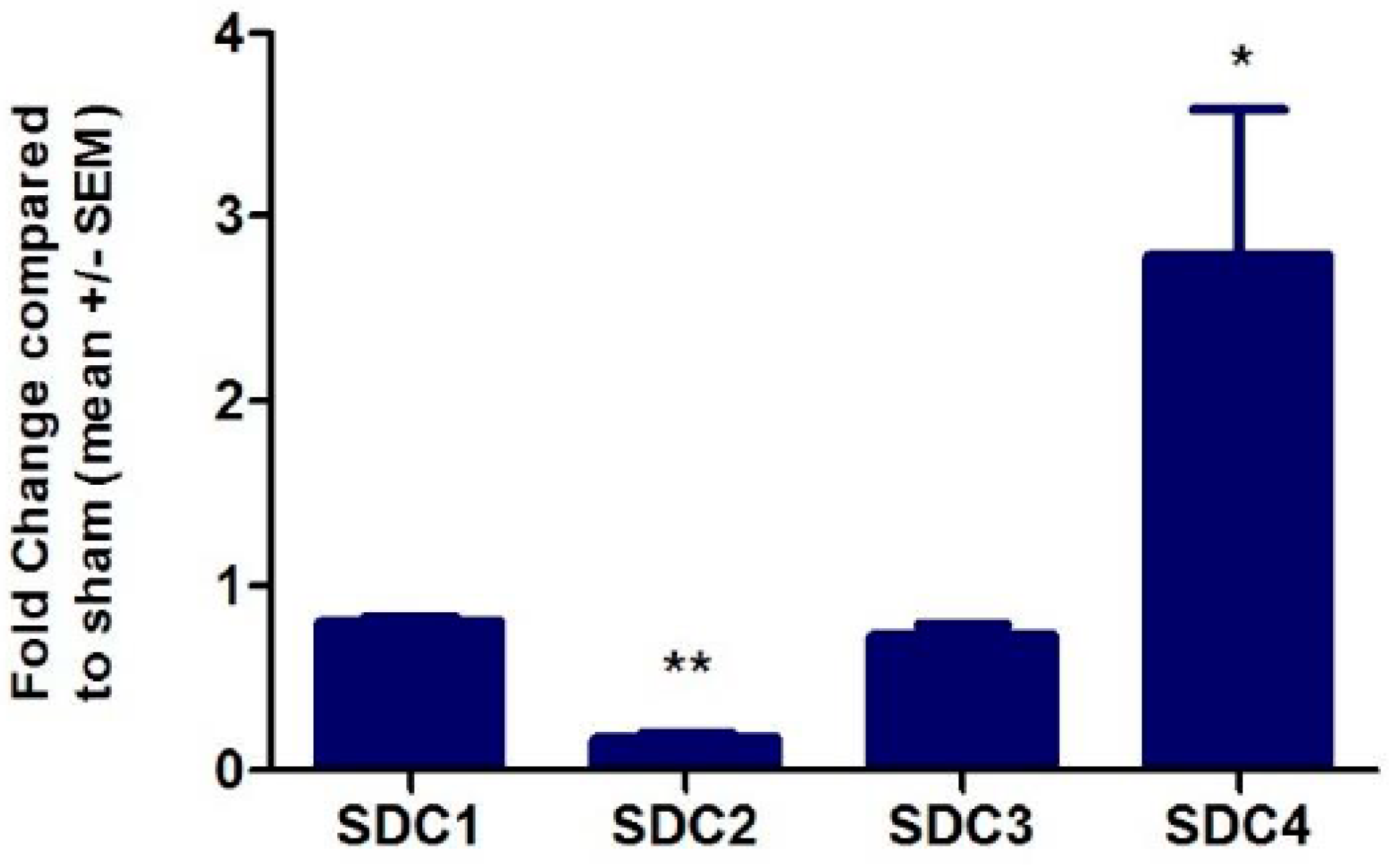
2.1. Effect of TNFα on Proteoglycan Transcription and Expression in HMVECs
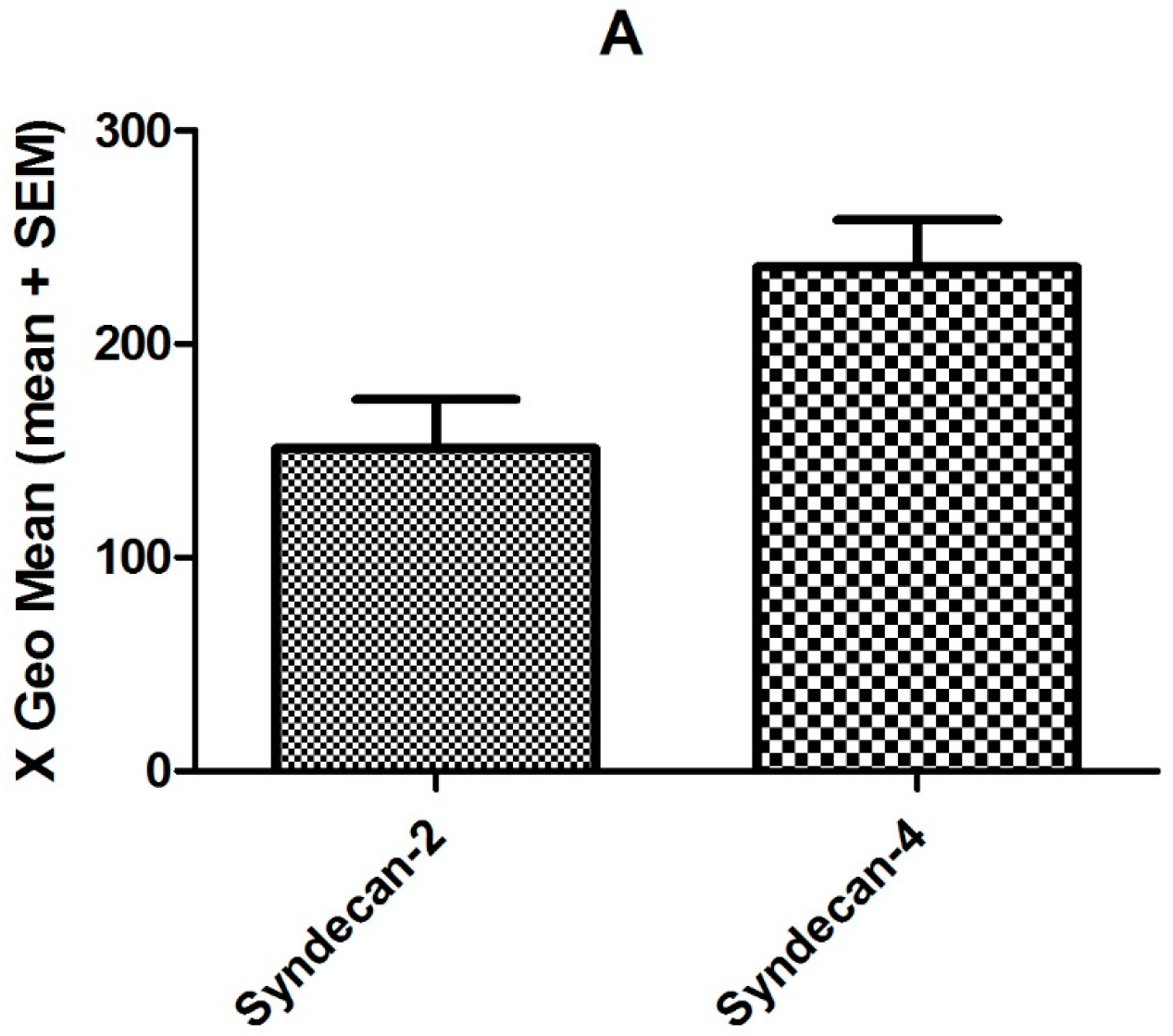
2.2. Effect of CXCL8 Treatment of Preinflamed HMVECs on Protein Expression
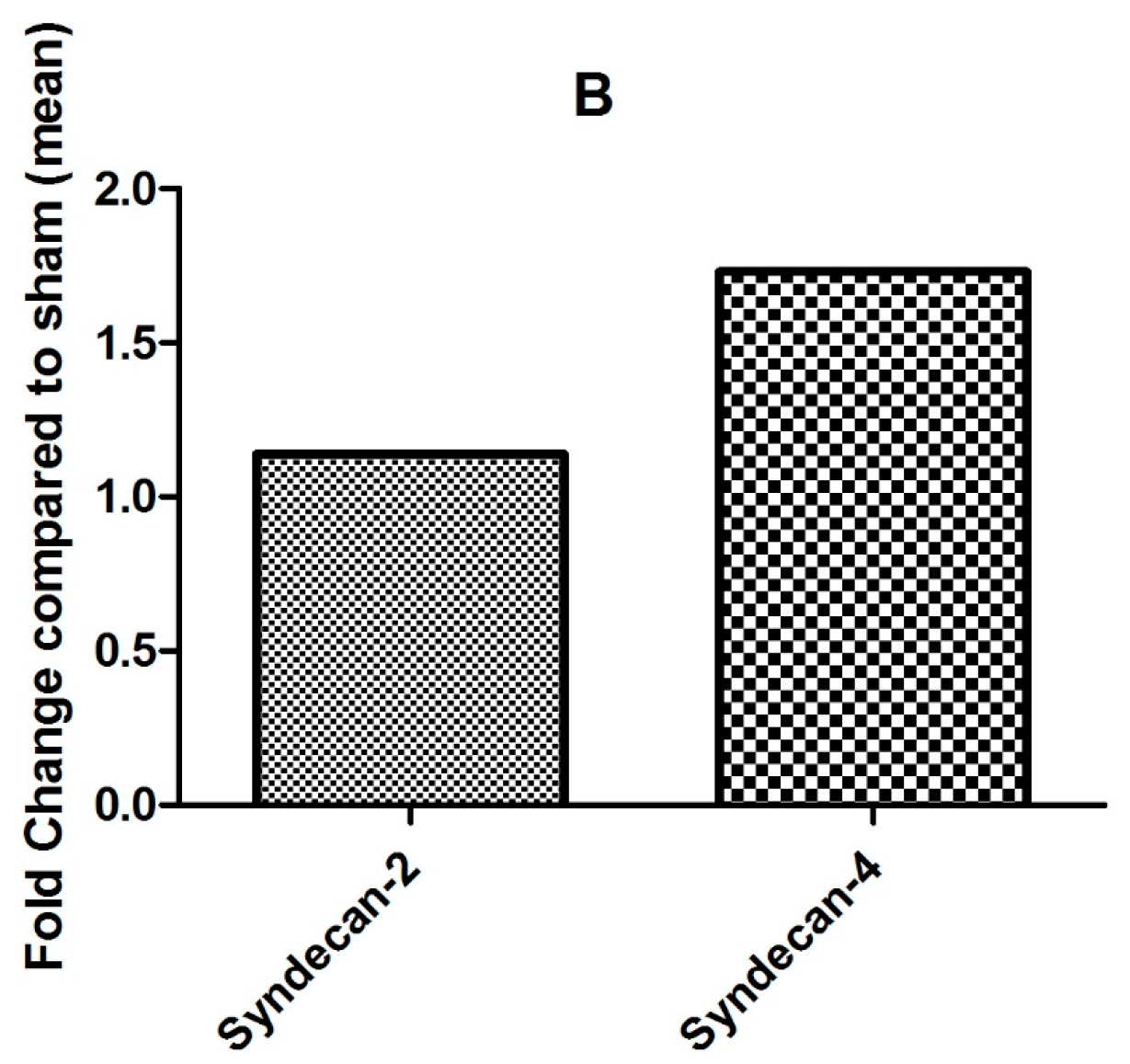
2.3. Effect of CXCL8 Treatment on Protein Expression Levels of TNFα-Preinflamed, Chondroitinase- and Heparinase-Treated HMVECs
3. Materials and Methods
3.1. Cell Culture
3.2. Whole Cell RNA Isolation
3.3. Gene Expression Analysis
3.4. Protein Isolation and Labeling
3.5. 2D Electrophoresis
3.6. Mass Spectrometry
3.7. RT-qPCR
3.8. Flow Cytometry
3.9. Functional Annotation Clustering
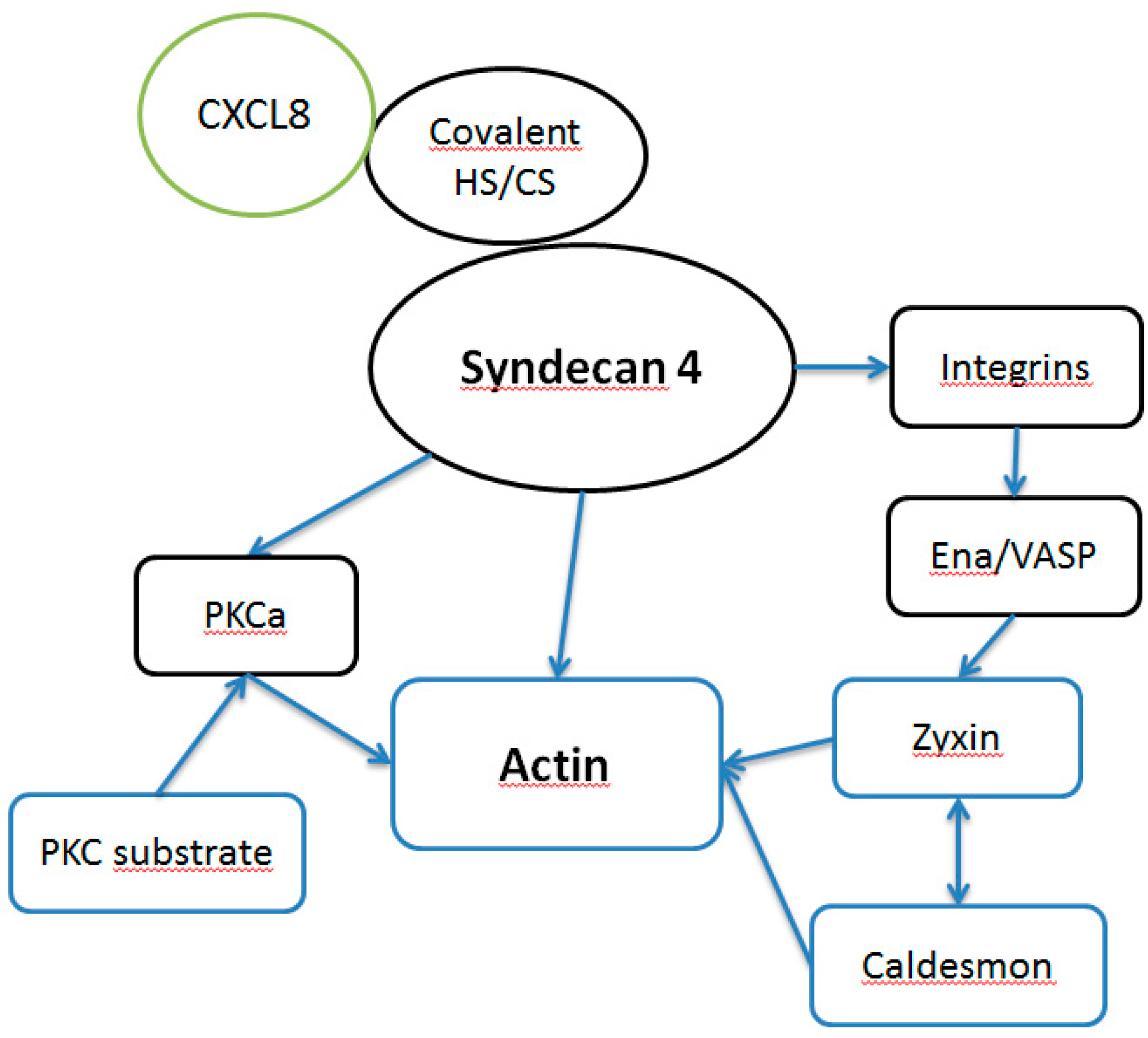
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GAG | Glycosaminoglycan |
| HS | Heparan sulphate |
| CS | Chondroitin sulfate |
| PG | Proteoglycan |
References
- Götte, M. Syndecans in inflammation. FASEB J. 2003, 17, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Esko, J.D.; Selleck, S.B. Order out of chaos: Assembly of ligand binding sites in heparan sulfate. Annu. Rev. Biochem. 2002, 71, 435–471. [Google Scholar] [CrossRef] [PubMed]
- Gesslbauer, B.; Rek, A.; Falsone, F.; Rajkovic, E.; Kungl, A.J. Proteoglycanomics: Tools to unravel the biological function of glycosaminoglycans. Proteomics 2007, 7, 2870–2880. [Google Scholar] [CrossRef] [PubMed]
- Ihrcke, N.S.; Wrenshall, L.E.; Lindman, B.J.; Platt, J.L. Role of heparan sulfate in immune system-blood vessel interactions. Immunol. Today 1993, 14, 500–505. [Google Scholar] [CrossRef]
- Rapraeger, A.; Jalkanen, M.; Endo, E.; Koda, J.; Bernfield, M. The Cell Surface Proteoglycan from Mouse Mammary Epithelial Cells Bears Chondroitin Sulfate and Heparan Sulfate Glycosaminoglycans. J. Biol. Chem. 1985, 260, 11046–11052. [Google Scholar] [PubMed]
- Couchman, J.R.; Gopal, S.; Lim, H.C.; Norgaard, S.; Multhaupt, H.A.B. Syndecans: From peripheral coreceptors to mainstream regulators of cell behaviour. Int. J. Exp. Pathol. 2015, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fico, A.; Maina, F.; Dono, R. Fine-tuning of cell signaling by glypicans. Cell. Mol. Life Sci. 2011, 68, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Palaiologou, M.; Delladetsima, I.; Tiniakos, D. CD138 (syndecan-1) expression in health and disease. Histol. Hostopathol. 2014, 29, 177–189. [Google Scholar]
- Theocharis, A.D.; Skandalis, S.S.; Tzanakakis, G.N.; Karamanos, N.K. Proteoglycans in health and disease: Novel roles for proteoglycans in malignancy and their pharmacological targeting. FEBS J. 2010, 277, 3904–3923. [Google Scholar] [CrossRef] [PubMed]
- Hacker, U.; Nybakken, K.; Perrimon, N. Heparan sulphate proteoglycans: The sweet side of development. Nat. Rev. Mol. Cell Biol. 2005, 6, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Deepa, S.S.; Yamada, S.; Zako, M.; Goldberger, O.; Sugahara, K. Chondroitin sulfate chains on syndecan-1 and syndecan-4 from normal murine mammary gland epithelial cells are structurally and functionally distinct and cooperate with heparan sulfate chains to bind growth factors. A novel function to control binding of midkine, pleiotrophin, and basic fibroblast growth factor. J. Biol. Chem. 2004, 279, 37368–37376. [Google Scholar] [PubMed]
- Hardingham, T.E.; Fosang, A.J. Proteoglycans: Many forms and many functions. FASEB J. 1992, 6, 861–870. [Google Scholar] [PubMed]
- Zhang, F.; Zhang, Z.; Lin, X.; Beenken, A.; Eliseenkova, A.V.; Mohammadi, M.; Linhardt, R.J. Compositional analysis of heparin/heparan sulfate interacting with fibroblast growth factor.fibroblast growth factor receptor complexes. Biochemistry 2009, 48, 8379–8386. [Google Scholar] [CrossRef] [PubMed]
- Asada, M.; Shinomiya, M.; Suzuki, M.; Honda, E.; Sugimoto, R.; Ikekita, M.; Imamura, T. Glycosaminoglycan affinity of the complete fibroblast growth factor family. Biochim. Biophys. Acta 2009, 1790, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Pichert, A.; Samsonov, S.A.; Theisgen, S.; Thomas, L.; Baumann, L.; Schiller, J.; Beck-Sickinger, A.G.; Huster, D.; Pisabarro, M.T. Characterization of the interaction of interleukin-8 with hyaluronan, chondroitin sulfate, dermatan sulfate and their sulfated derivatives by spectroscopy and molecular modeling. Glycobiology 2012, 22, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.K.; Paavola, C.D.; Johnson, Z.; Gaudry, J.-P.; Geretti, E.; Borlat, F.; Kungl, A.J.; Proudfoot, A.E.; Handel, T.M. Identification of the glycosaminoglycan binding site of the CC chemokine, MCP-1: Implications for structure and function in vivo. J. Biol. Chem. 2004, 279, 22294–22305. [Google Scholar] [CrossRef] [PubMed]
- Campanella, G.S.V.; Lee, E.M.J.; Sun, J.; Luster, A.D. CXCR3 and heparin binding sites of the chemokine IP-10 (CXCL10). J. Biol. Chem. 2003, 278, 17066–17074. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, C.; Mayer, U.; Aumailley, M.; Timpl, R. Basement-membrane heparan sulfate proteoglycan binds to laminin by its heparan sulfate chains and to nidogen by sites in the protein core. Eur. J. Biochem. 1992, 208, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Laterra, J.; Silbert, J.E.; Culp, L.A. Cell surface heparan sulfate mediates some adhesive responses to glycosaminoglycan-binding matrices, including fibronectin. J. Cell Biol. 1983, 96, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Bisio, A.; Vecchietti, D.; Citterio, L.; Guerrini, M.; Raman, R.; Bertini, S.; Eisele, G.; Naggi, A.; Sasisekharan, R.; Torri, G. Structural features of low-molecular-weight heparins affecting their affinity to antithrombin. Thromb. Haemost. 2009, 102, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R.D.; Damus, P.S. The purification and mechanism of action of human antithrombin-heparin cofactor. J. Biol. Chem. 1973, 248, 6490–6505. [Google Scholar] [PubMed]
- Ji, Z.-S.; Dichek, H.L.; Miranda, R.D.; Mahley, R.W. Heparan Sulfate Proteoglycans Participate in Hepatic Lipaseand Apolipoprotein E-mediated Binding and Uptake of Plasma Lipoproteins, Including High Density Lipoproteins. J. Biol. Chem. 1997, 272, 31285–31292. [Google Scholar] [CrossRef] [PubMed]
- Libeu, C.P.; Lund-Katz, S.; Phillips, M.C.; Wehrli, S.; Hernaiz, M.J.; Capila, I.; Linhardt, R.J.; Raffai, R.L.; Newhouse, Y.M.; Zhou, F.; et al. New insights into the heparan sulfate proteoglycan-binding activity of apolipoprotein E. J. Biol. Chem. 2001, 276, 39138–39144. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.-S.; Pitas, R.E.; Mahley, R.W. Differential Cellular Accumulation/Retention of Apolipoprotein E Mediated by Cell Surface Heparan Sulfate Proteoglycans: Apolipoproteins E3 and E2 greater than E4. J. Biol. Chem. 1998, 273, 13452–13460. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Hassan, M.I.; Tomar, A.K.; Kashav, T.; Nautiyal, J.; Singh, S.; Singh, T.P.; Yadav, S. Proteomic analysis of heparin-binding proteins from human seminal plasma: A step towards identification of molecular markers of male fertility. J. Biosci. 2009, 34, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Ori, A.; Wilkinson, M.C.; Fernig, D.G. A systems biology approach for the investigation of the heparin/heparan sulfate interactome. J. Biol. Chem. 2011, 286, 19892–19904. [Google Scholar] [CrossRef] [PubMed]
- Gesslbauer, B.; Derler, R.; Handwerker, C.; Seles, E.; Kungl, A.J. Exploring the glycosaminoglycan-protein interaction network by glycan-mediated pull-down proteomics. Electrophoresis 2016, 37, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Ellyard, J.I.; Simson, L.; Bezos, A.; Johnston, K.; Freeman, C.; Parish, C.R. Eotaxin selectively binds heparin. An interaction that protects eotaxin from proteolysis and potentiates chemotactic activity in vivo. J. Biol. Chem. 2007, 282, 15238–15247. [Google Scholar] [CrossRef] [PubMed]
- Sadir, R.; Imberty, A.; Baleux, F.; Lortat-Jacob, H. Heparan sulfate/heparin oligosaccharides protect stromal cell-derived factor-1 (SDF-1)/CXCL12 against proteolysis induced by CD26/dipeptidyl peptidase IV. J. Biol. Chem. 2004, 279, 43854–43860. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Zhang, L.; Yabe, T.; Kuberan, B.; Beeler, D.L.; Love, A.; Rosenberg, R.D. The involvement of heparan sulfate (HS) in FGF1/HS/FGFR1 signaling complex. J. Biol. Chem. 2003, 278, 17121–17129. [Google Scholar] [CrossRef] [PubMed]
- Ecke, S.; Geiger, M.; Binder, B.R. Heparin binding of protein-C inhibitor--analysis of the effect of heparin on the interaction of protein-C inhibitor with tissue kallikrein. Eur. J. Biochem. 1997, 248, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Young, J.H.; Krahn, J.M.; Song, D.; Corbett, K.D.; Chazin, W.J.; Pedersen, L.C.; Esko, J.D. Stable RAGE-heparan sulfate complexes are essential for signal transduction. ACS Chem. Biol. 2013, 8, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Nieto, L.; Canales, A.; Fernandez, I.S.; Santillana, E.; Gonzalez-Corrochano, R.; Redondo-Horcajo, M.; Canada, F.J.; Nieto, P.; Martin-Lomas, M.; Gimenez-Gallego, G.; et al. Heparin modulates the mitogenic activity of fibroblast growth factor by inducing dimerization of its receptor. A 3D view by using NMR. Chembiochem Eur. J. Chem. Biol. 2013, 14, 1732–1744. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.R.; Kunkel, S.L.; Todd, R.F.; Weiss, S.J. Regulation of transendothelial neutrophil migration by endogenous interleukin-8. Science 1991, 254, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Netelenbos, T.; van den Born, J.; Kessler, F.L.; Zweegman, S.; Merle, P.A.; van Oostveen, J.W.; Zwaginga, J.J.; Huijgens, P.C.; Drager, A.M. Proteoglycans on bone marrow endothelial cells bind and present SDF-1 towards hematopoietic progenitor cells. Leukemia 2003, 17, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.M.; Webster, S.J.; Flower, D.; Woll, P.J. Interleukin-8/CXCL8 is a growth factor for human lung cancer cells. Br. J. Cancer 2004, 91, 1970–1976. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Singh, D. Neutrophil chemotaxis caused by chronic obstructive pulmonary disease alveolar macrophages: The role of CXCL8 and the receptors CXCR1/CXCR2. J. Pharmacol. Exp. Ther. 2013, 347, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Szekanecz, Z.; Kim, J.; Koch, A.E. Chemokines and chemokine receptors in rheumatoid arthritis. Semin. Immunol. 2003, 15, 15–21. [Google Scholar] [CrossRef]
- Kuschert, G.S.V.; Coulin, F.; Power, C.A.; Proudfoot, A.E.I.; Hubbard, R.E.; Hoogewerf, A.J.; Wells, T.N.C. Glycosaminoglycans Interact Selectively with Chemokines and Modulate Receptor Binding and Cellular Responses. Biochemistry 1999, 38, 12959–12968. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, C.; Finn, A. Chemokine receptors and their role in inflammation and infectious diseases. Blood 2000, 95, 3032–3043. [Google Scholar] [PubMed]
- Proudfoot, A.E.I. The biological relevance of chemokine-proteoglycan interactions. Biochem. Soc. Trans. 2006, 34, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Esko, J.D. Demystifying heparan sulfate-protein interactions. Annu. Rev. Biochem. 2014, 83, 129–157. [Google Scholar] [CrossRef] [PubMed]
- Marshall, L.J.; Ramdin, L.S.P.; Brooks, T.; DPhil, P.C.; Shute, J.K. Plasminogen Activator Inhibitor-1 Supports IL-8-Mediated Neutrophil Transendothelial Migration by Inhibition of the Constitutive Shedding of Endothelial IL-8/Heparan Sulfate/Syndecan-1 Complexes. J. Immunol. 2003, 171, 2057–2065. [Google Scholar] [CrossRef] [PubMed]
- Horuk, R.; Chitnis, C.E.; Darbonne, W.C.; Colby, T.J.; Rybicki, A.; Hadley, T.J.; Miller, L.H. A receptor for the malarial parasite Plasmodium vivax: The erythrocyte chemokine receptor. Science 1993, 261, 1182–1184. [Google Scholar] [CrossRef] [PubMed]
- Szabo, M.C.; Soo, K.S.; Zlotnik, A.; Schall, T.J. Chemokine Class Differences in Binding to the Duffy Antigen-Erythrocyte Chemokine Receptor. J. Biol. Chem. 1995, 270, 25348–25351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Liu, J.; Zhang, Z.; Chen, Y. Suppressing Syndecan-1 Shedding Ameliorates Intestinal Epithelial Inflammation through Inhibiting NF-kappaB Pathway and TNF-alpha. Gastroenterol. Res. Pract. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Nam, E.J.; Park, P.W. Shedding of cell membrane-bound proteoglycans. Methods Mol. Biol. 2012, 836, 291–305. [Google Scholar] [PubMed]
- Manon-Jensen, T.; Itoh, Y.; Couchman, J.R. Proteoglycans in health and disease: The multiple roles of syndecan shedding. FEBS J. 2010, 277, 3876–3889. [Google Scholar] [CrossRef] [PubMed]
- Hammond, E.; Khurana, A.; Shridhar, V.; Dredge, K. The Role of Heparanase and Sulfatases in the Modification of Heparan Sulfate Proteoglycans within the Tumor Microenvironment and Opportunities for Novel Cancer Therapeutics. Front. Oncol. 2014, 4, 195. [Google Scholar] [CrossRef] [PubMed]
- Nadir, Y.; Brenner, B. Heparanase multiple effects in cancer. Thromb. Res. 2014, 133, S90–S94. [Google Scholar] [CrossRef]
- Weber, C. Investigating Chemokine Co-Receptor Interactions. Ph.D. Thesis, University of Graz, Graz, Austria, 2012. [Google Scholar]
- Cheng, S.S.; Lukacs, N.W.; Kunkel, S.L. Eotaxin/CCL11 suppresses IL-8/CXCL8 secretion from human dermal microvascular endothelial cells. J. Immunol. 2002, 168, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y. The role of chemokines in neutrophil biology. Front. Biosci. 2008, 13, 2400–2407. [Google Scholar] [CrossRef] [PubMed]
- Dave, J.M.; Bayless, K.J. Vimentin as an integral regulator of cell adhesion and endothelial sprouting. Microcirculation 2014, 21, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Ivaska, J.; Pallari, H.-M.; Nevo, J.; Eriksson, J.E. Novel functions of vimentin in cell adhesion, migration, and signaling. Exp. Cell Res. 2007, 313, 2050–2062. [Google Scholar] [CrossRef] [PubMed]
- Adage, T.; Konya, V.; Weber, C.; Strutzmann, E.; Fuchs, T.; Zankl, C.; Gerlza, T.; Jeremic, D.; Heinemann, A.; Kungl, A.J. Targeting glycosaminoglycans in the lung by an engineered CXCL8 as a novel therapeutic approach to lung inflammation. Eur. J. Pharmacol. 2015, 748, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; Morisaki, T.; Nishiyama, Y.; Marumoto, T.; Tada, K.; Hara, T.; Masuko, N.; Inagaki, M.; Hatakeyama, K.; Saya, H. Zyxin, a Regulator of Actin Filament Assembly, Targets the Mitotic Apparatus by Interacting with H-Warts/Lats1 Tumor Suppressor. J. Cell Biol. 2000, 149, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Pichon, S.; Bryckaert, M.; Berrou, E. Control of actin dynamics by p38 MAP kinase—Hsp27 distribution in the lamellipodium of smooth muscle cells. J. Cell Sci. 2004, 117, 2569–2577. [Google Scholar] [CrossRef] [PubMed]
- Nagano, K.; Bornhauser, B.C.; Warnasuriya, G.; Entwistle, A.; Cramer, R.; Lindholm, D.; Naaby-Hansen, S. PDGF regulates the actin cytoskeleton through hnRNP-K-mediated activation of the ubiquitin E3-ligase MIR. EMBO J. 2006, 25, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, T.; Nemere, L. Actin and Keratin are Binding Partners of the 1,25D3-MARRS Receptor/PDIA3/ERp57. Immunol. Endocr. Metab. Agents Med. Chem. 2014, 14, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Snee, M. RNA trafficking and stabilization elements associate with multiple brain proteins. J. Cell Sci. 2002, 115, 4661–4669. [Google Scholar] [CrossRef] [PubMed]
- Sobue, K.; Kanda, K.; Tanaka, T.; Ueki, N. Caldesmon: A common actin-linked regulatory protein in the smooth muscle and nonmuscle contractile system. J. Cell. Biochem. 1988, 37, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Echtermeyer, F.; Baciu, P.C.; Saoncella, S.; Ge, Y.; Goetinck, P.F. Syndecan-4 core protein is sufficient for the assembly of focal adhesions and actin stress fibers. J. Cell Sci. 1999, 112 Pt 20, 3433–3441. [Google Scholar] [PubMed]
- Larsson, C. Protein kinase C and the regulation of the actin cytoskeleton. Cell. Signal. 2006, 18, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Keum, E.; Kim, Y.; Kim, J.; Kwon, S.; Lim, Y.; Han, I.; Oh, E.-S. Syndecan-4 regulates localization, activity and stability of protein kinase C-alpha. Biochem. J. 2004, 378, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Saoncella, S.; Echtermeyer, F.; Denhez, F.; Nowlen, J.K.; Mosher, D.F.; Robinson, S.D.; Hynes, R.O.; Goetinck, P.F. Syndecan-4 signals cooperatively with integrins in a Rhodependent manner in the assembly of focal adhesions and actin stress fibers. Proc. Natl. Acad. Sci. USA 1999, 96, 2805–2810. [Google Scholar] [CrossRef] [PubMed]
- Reinhard, M.; Jarchau, T.; Walter, U. Actin-based motility: Stop and go with Ena/VASP proteins. Trends Biochem. Sci. 2001, 26, 243–249. [Google Scholar] [CrossRef]
- Heukeshoven, J.; Dernick, R. Simplified method for silver staining of proteins in polyacrylamide gels and the mechanism of silver staining. Electrophoresis 1985, 6, 103–112. [Google Scholar] [CrossRef]
- Strutzmann, E. Investigating the Glycan-Mediated Chemokine Mode of Action. Ph.D. Thesis, University of Graz, Graz, Austria, 2014. [Google Scholar]
- Miller, I. Application of 2D DIGE in animal proteomics. Methods Mol. Biol. 2012, 854, 373–396. [Google Scholar] [PubMed]
- Gesslbauer, B.; Poljak, A.; Handwerker, C.; Schuler, W.; Schwendenwein, D.; Weber, C.; Lundberg, U.; Meinke, A.; Kungl, A.J. Comparative membrane proteome analysis of three Borrelia species. Proteomics 2012, 12, 845–858. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Master No. | t-Test | Av. Ratio | Identified Protein | UniProtKB | MW (Da) |
|---|---|---|---|---|---|
| 292 | 0.16 | −1.59 | Prelamin-A/C | P02545 | 74,380 |
| 373 | 0.086 | −1.58 | Alpha-2-HS-glycoprotein | P02765 | 40,098 |
| 389 | 0.25 | −1.61 | Cytoskeleton-associated protein 4 or | Q07065 | 66,097 |
| Heterogeneous nuclear ribonucleoprotein K | P61978 | 51,230 | |||
| 723 | 0.44 | 1.53 | Vimentin | P08670 | 53,676 |
| 1020 | 0.003 | 1.59 * | Vimentin | P08670 | 53,676 |
| 1176 | 0.039 | 1.56 * | Transgelin-2 | P37802 | 22,548 |
| 1194 | 0.063 | 1.73 | 40S ribosomal protein S18 | P62269 | 17,708 |
| 1233 | 0.028 | 1.50 * | 40S ribosomal protein S25/ | P62851 | 13,791 |
| Prefoldin subunit 2 | Q9UHV9 | 16,695 | |||
| 1294 | - | 1.76 | Histone 4 | P62805 | 11,360 |
| Master No. | t-Test | Av. Ratio | Identified Protein | UniProtKB | MW (Da) |
|---|---|---|---|---|---|
| 253 | 0.41 | 1.75 | Zyxin | Q15942 | 62,463 |
| 254 | 0.43 | 1.61 | Caldesmon | Q05682 | 93,251 |
| 257 | - | 1.81 | Caldesmon | Q05682 | 93,251 |
| 259 | 0.040 | 1.64 * | Far upstream element binding-protein 2 | Q92945 | 73,355 |
| 331 | 0.1 | −2.46 | Intercellular adhesion molecule 1 | P05362 | 58,587 |
| 361 | - | 1.62 | Protein kinase C and casein kinase substrate in neurons protein 2 | Q9UNF0 | 56,046 |
| 374 | 0.15 | −2.06 | WAS protein family homolog 1 | A8K0Z3 | 50,354 |
| 378 | 0.13 | −1.88 | Protein disulfide-isomerase A3 | P30101 | 57,146 |
| 386 | 0.086 | −1.56 | Cytoskeleton-associated protein 4 | Q07065 | 66,097 |
| 387 | 0.077 | −1.62 | Cytoskeleton-associated protein 4 | Q07065 | 66,097 |
| 388 | 0.050 | −1.77 | Cytoskeleton-associated protein 4 | Q07065 | 66,097 |
| 392 | 0.55 | −1.87 | Heterogenous ribonucleoprotein K | P61978 | 51,230 |
| 414 | 0.14 | −2.09 | Prolyl 4-hydroxylase subunit alpha-1 | P13674 | 61,296 |
| 552 | 0.21 | 1.57 | S-adenosylmethionine synthase isoform type-2 | P31153 | 43,975 |
| 671 | 0.39 | 1.52 | Actin, cytoplasmic 1/Nestin | P60709 | 42,052 |
| P48681 | 177,439 | ||||
| 694 | 0.28 | 2.17 | Actin, cytoplasmic 1 | P60709 | 42,052 |
| 695 | - | 1.88 | COP9 signalosome complex subunit 4 | Q9BT78 | 46,525 |
| 1012 | 0.27 | 1.53 | Heat shock protein beta-1 | P04792 | 22,826 |
| 1208 | - | −1.86 | 40S ribosomal protein S15 | P62841 | 17,029 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derler, R.; Gesslbauer, B.; Weber, C.; Strutzmann, E.; Miller, I.; Kungl, A. Glycosaminoglycan-Mediated Downstream Signaling of CXCL8 Binding to Endothelial Cells. Int. J. Mol. Sci. 2017, 18, 2605. https://doi.org/10.3390/ijms18122605
Derler R, Gesslbauer B, Weber C, Strutzmann E, Miller I, Kungl A. Glycosaminoglycan-Mediated Downstream Signaling of CXCL8 Binding to Endothelial Cells. International Journal of Molecular Sciences. 2017; 18(12):2605. https://doi.org/10.3390/ijms18122605
Chicago/Turabian StyleDerler, Rupert, Bernd Gesslbauer, Corinna Weber, Elisabeth Strutzmann, Ingrid Miller, and Andreas Kungl. 2017. "Glycosaminoglycan-Mediated Downstream Signaling of CXCL8 Binding to Endothelial Cells" International Journal of Molecular Sciences 18, no. 12: 2605. https://doi.org/10.3390/ijms18122605
APA StyleDerler, R., Gesslbauer, B., Weber, C., Strutzmann, E., Miller, I., & Kungl, A. (2017). Glycosaminoglycan-Mediated Downstream Signaling of CXCL8 Binding to Endothelial Cells. International Journal of Molecular Sciences, 18(12), 2605. https://doi.org/10.3390/ijms18122605