Effect of Omega-3 Fatty Acid Supplementation on Oxylipins in a Routine Clinical Setting

Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Participants
2.2. Effect of n-3 PUFA Intake on Fatty Acid Composition in Blood Cells
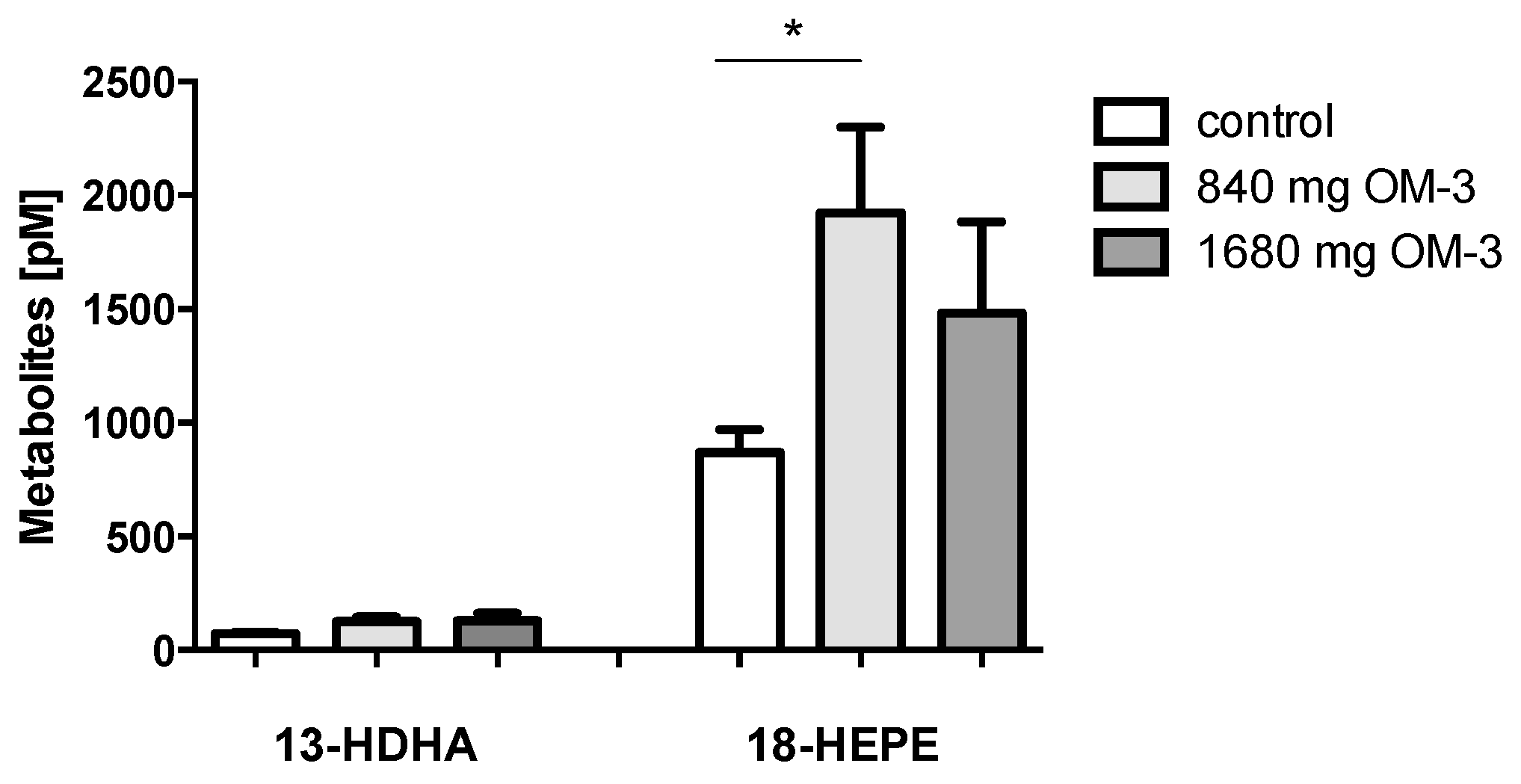
2.3. Effect of n-3 PUFA Treatment on Plasma Levels of 13-HDHA and 18-HEPE
2.4. Changes in the Profile of LOX-Dependent AA-, EPA-, and DHA-Derived Metabolites
2.5. Effect of EPA and DHA Supplementation on AA-, DHA-, and EPA-Derived Epoxy Metabolites
3. Discussion
4. Materials and Methods
4.1. Patients and Blood Sampling
4.2. Determination of Fatty Acid Pattern
4.3. Sample Preparation and LC/ESI-MS/MS
4.4. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| LA | linoleic acid |
| HODE | hydroxyoctadecadienoic acid |
| EpOME | epoxyoctadecenoic acid |
| DiHOME | dihydroxyoctadecenoic acid |
| TriHOME | trihydroxyoctadecenoic acid |
| HETE | hydroxyeicosatetraenoic acid |
| EET | epoxyeicosatrienoic acid |
| DHET | dihydroxyeicosatrienoic acid |
| HOTrE | hydroxyoctadecatrienoic acid |
| EpODE | epoxyoctadecadienoic acid |
| DiHODE | dihydroxyoctadecadienoic acid |
| HEPE | hydroxyeicosapentaenoic acid |
| EEQ | epoxyeicosatetraenoic acid |
| DiHETE | dihydroxyeicosatetraenoic acid |
| HDHA | hydroxydocosahexaenoic acid |
| EDP | epoxydocosapentaenoic acid |
| DHDP | dihydroxydocosapentaenoic acid |
References
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kang, J.X.; Leaf, A. Differential effects of various eicosanoids on the production or prevention of arrhythmias in cultured neonatal rat cardiac myocytes. Prostaglandins 1997, 54, 511–530. [Google Scholar] [CrossRef]
- Siscovick, D.S.; Lemaitre, R.N.; Mozaffarian, D. The fish story: A diet-heart hypothesis with clinical implications: n-3 polyunsaturated fatty acids, myocardial vulnerability, and sudden death. Circulation 2003, 107, 2632–2634. [Google Scholar] [CrossRef] [PubMed]
- Nestel, P.; Shige, H.; Pomeroy, S.; Cehun, M.; Abbey, M.; Raederstorff, D. The n-3 fatty acids eicosapentaenoic acid and docosahexaenoic acid increase systemic arterial compliance in humans. Am. J. Clin. Nutr. 2002, 76, 326–330. [Google Scholar] [PubMed]
- Morris, M.C.; Sacks, F.; Rosner, B. Does fish oil lower blood pressure? A meta-analysis of controlled trials. Circulation 1993, 88, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.A.; Hochland, M.C. A comparison of the influence on plasma lipids and platelet function of supplements of omega 3 and omega 6 polyunsaturated fatty acids. Br. J. Nutr. 1983, 50, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Chiu, C.Y.; Gomolka, B.; Waechter, S.F.; Wiedenmann, B. Omega-3 fatty acids and their lipid mediators: Towards an understanding of resolvin and protectin formation. Prostaglandins Lipid Mediat. 2012, 97, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Worgall, T.S.; Sturley, S.L.; Seo, T.; Osborne, T.F.; Deckelbaum, R.J. Polyunsaturated fatty acids decrease expression of promoters with sterol regulatory elements by decreasing levels of mature sterol regulatory element-binding protein. J. Biol. Chem. 1998, 273, 25537–25540. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.E.; Lambert, M.H.; Montana, V.G.; Parks, D.J.; Blanchard, S.G.; Brown, P.J.; Sternbach, D.D.; Lehmann, J.M.; Wisely, G.B.; Willson, T.M.; et al. Molecular recognition of fatty acids by peroxisome proliferator-activated receptors. Mol. Cell 1999, 3, 397–403. [Google Scholar] [CrossRef]
- Ye, D.; Zhang, D.; Oltman, C.; Dellsperger, K.; Lee, H.C.; VanRollins, M. Cytochrome p-450 epoxygenase metabolites of docosahexaenoate potently dilate coronary arterioles by activating large-conductance calcium-activated potassium channels. J. Pharmacol. Exp. Ther. 2002, 303, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profile in man primarily via the CYP-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Ostermann, A.I.; Stork, L.; Fritzsch, S.; Kohrs, H.; Greupner, T.; Hahn, A.; Schebb, N.H. Effect of DHA supplementation on oxylipin levels in plasma and immune cell stimulated blood. Prostaglandins Leukot. Essent. Fatty Acids 2017, 121, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Ostermann, A.I.; Stork, L.; Kutzner, L.; Kohrs, H.; Greupner, T.; Hahn, A.; Schebb, N.H. Effects of docosahexaenoic acid supplementation on PUFA levels in red blood cells and plasma. Prostaglandins Leukot. Essent. Fatty Acids 2016, 115, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Schebb, N.H.; Ostermann, A.I.; Yang, J.; Hammock, B.D.; Hahn, A.; Schuchardt, J.P. Comparison of the effects of long-chain omega-3 fatty acid supplementation on plasma levels of free and esterified oxylipins. Prostaglandins Lipid Mediat. 2014, 113–115, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Schmidt, S.; Kressel, G.; Willenberg, I.; Hammock, B.D.; Hahn, A.; Schebb, N.H. Modulation of blood oxylipin levels by long-chain omega-3 fatty acid supplementation in hyper- and normolipidemic men. Prostaglandins Leukot. Essent. Fatty Acids 2014, 90, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Schmidt, S.; Kressel, G.; Dong, H.; Willenberg, I.; Hammock, B.D.; Hahn, A.; Schebb, N.H. Comparison of free serum oxylipin concentrations in hyper- vs. Normolipidemic men. Prostaglandins Leukot. Essent. Fatty Acids 2013, 89, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Berglund, L.; Brunzell, J.D.; Goldberg, A.C.; Goldberg, I.J.; Sacks, F.; Murad, M.H.; Stalenhoef, A.F. Evaluation and treatment of hypertriglyceridemia: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2012, 97, 2969–2989. [Google Scholar] [CrossRef] [PubMed]
- Siscovick, D.S.; Barringer, T.A.; Fretts, A.M.; Wu, J.H.; Lichtenstein, A.H.; Costello, R.B.; Kris-Etherton, P.M.; Jacobson, T.A.; Engler, M.B.; Alger, H.M.; et al. Omega-3 polyunsaturated fatty acid (fish oil) supplementation and the prevention of clinical cardiovascular disease: A science advisory from the american heart association. Circulation 2017, 135, e867–e884. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Omega-3 fatty acids and cardiovascular disease: New recommendations from the American heart association. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Chiang, N.; Serhan, C.N. Elucidation of novel 13-series resolvins that increase with atorvastatin and clear infections. Nat. Med. 2015, 21, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Planaguma, A.; Pfeffer, M.A.; Rubin, G.; Croze, R.; Uddin, M.; Serhan, C.N.; Levy, B.D. Lovastatin decreases acute mucosal inflammation via 15-epi-lipoxin a4. Mucosal Immunol. 2010, 3, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.-L. Resolvins. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Von Schacky, C. Omega-3 fatty acids in cardiovascular disease—An uphill battle. Prostaglandins Leukot. Essent. Fatty Acids 2015, 92, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.F.; Pillai, P.S.; Recchiuti, A.; Yang, R.; Serhan, C.N. Pro-resolving actions and stereoselective biosynthesis of 18S E-series resolvins in human leukocytes and murine inflammation. J. Clin. Investig. 2011, 121, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Sapieha, P.; Stahl, A.; Chen, J.; Seaward, M.R.; Willett, K.L.; Krah, N.M.; Dennison, R.J.; Connor, K.M.; Aderman, C.M.; Liclican, E.; et al. 5-lipoxygenase metabolite 4-HDHA is a mediator of the antiangiogenic effect of omega-3 polyunsaturated fatty acids. Sci. Transl. Med. 2011, 3, 69ra12. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C.; Markovic, M.; Blossey, K.; Wallukat, G.; Fischer, R.; Dechend, R.; Konkel, A.; von Schacky, C.; Luft, F.C.; Muller, D.N.; et al. Arachidonic acid-metabolizing cytochrome p450 enzymes are targets of {omega}-3 fatty acids. J. Biol. Chem. 2010, 285, 32720–32733. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Schwichtenberg, K.A.; Hanson, N.Q.; Tsai, M.Y. Incorporation and clearance of omega-3 fatty acids in erythrocyte membranes and plasma phospholipids. Clin. Chem. 2006, 52, 2265–2272. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Von Schacky, C. The omega-3 index: A new risk factor for death from coronary heart disease? Prev. Med. 2004, 39, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467S–1476S. [Google Scholar] [PubMed]
- Jung, F.; Schulz, C.; Blaschke, F.; Muller, D.N.; Mrowietz, C.; Franke, R.P.; Lendlein, A.; Schunck, W.H. Effect of cytochrome p450-dependent epoxyeicosanoids on ristocetin-induced thrombocyte aggregation. Clin. Hemorheol. Microcirc. 2012, 52, 403–416. [Google Scholar] [PubMed]
- Schuchardt, J.P.; Schneider, I.; Willenberg, I.; Yang, J.; Hammock, B.D.; Hahn, A.; Schebb, N.H. Increase of EPA-derived hydroxy, epoxy and dihydroxy fatty acid levels in human plasma after a single dose of long-chain omega-3 PUFA. Prostaglandins Lipid Mediat. 2014, 109–111, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Arnold, W.R.; Baylon, J.L.; Tajkhorshid, E.; Das, A. Asymmetric binding and metabolism of polyunsaturated fatty acids (PUFAs) by CYP2J2 epoxygenase. Biochemistry 2016, 55, 6969–6980. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, Y.; Ye, Y.; Lin, Y.; Freeberg, S.Y.; Huang, M.H.; Perez-Polo, J.R.; Uretsky, B.F. Aspirin augments 15-epi-lipoxin A4 production by lipopolysaccharide, but blocks the pioglitazone and atorvastatin induction of 15-epi-lipoxin A4 in the rat heart. Prostaglandins Lipid Mediat. 2007, 83, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Xing, X.; Hu, C.; Yu, H.; Dong, Q.; Chang, G.; Qin, S.; Liu, J.; Zhang, D. 15-lipoxygenase-1 is involved in the effects of atorvastatin on endothelial dysfunction. Mediat. Inflamm. 2016, 2016, 6769032. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, R.; Gerrity, R.G.; Gu, J.L.; Lanting, L.; Thomas, L.; Nadler, J.L. Role of 12-lipoxygenase and oxidant stress in hyperglycaemia-induced acceleration of atherosclerosis in a diabetic pig model. Diabetologia 2002, 45, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Schmocker, C.; Kassner, U.; Kiesler, S.; Bismarck, M.; Rothe, M.; Steinhagen-Thiessen, E.; Weylandt, K.H. A lipidomic analysis approach in patients undergoing lipoprotein apheresis. Atherosclerosis 2016, 249, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, A.I.; Muller, M.; Willenberg, I.; Schebb, N.H. Determining the fatty acid composition in plasma and tissues as fatty acid methyl esters using gas chromatography—A comparison of different derivatization and extraction procedures. Prostaglandins Leukot. Essent. Fatty Acids 2014, 91, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, A.I.; Willenberg, I.; Schebb, N.H. Comparison of sample preparation methods for the quantitative analysis of eicosanoids and other oxylipins in plasma by means of LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 1403–1414. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control (n = 16) | 840 mg OM-3 (n = 11) | 1680 OM-3 (n = 8) |
|---|---|---|---|
| Age | 57 (42–71) | 56 (43–73) | 62 (52–72) |
| Body weight (kg) | 89 (72.1–113.6) | 91.1 (76.5–110.5) | 80 (64–108) |
| BMI (kg/m2) | 28.9 (24.1–35.5) | 30.4 (25.4–39.2) | 27.8 (22.3–30.3) |
| Lipid Parameters | |||
| Total cholesterol (mg/dL) | 180.8 (104–243) | 178.4 (86–268) | 226.4 (108–461) |
| LDL (mg/dL) | 111.8 (52–281) | 107.6 (41–146) | 147.1 (52–332) |
| HDL (mg/dL) | 48.9 (26–86) | 53.4 (36–108) | 45 (33–71) |
| LDL/HDL quotient | 2.3 | 2.0 | 3.3 |
| TG (mg/dL) | 143.4 (57–281) | 145.6 (84–247) | 261.6 (123–490) |
| Medication | |||
| Aspirin | 16 | 11 | 8 |
| Clopidogrel | 6 | 3 | 2 |
| Prasugrel | 0 | 1 | 1 |
| β-blockers | 9 | 9 | 8 |
| Statins | 15 | 9 | 5 |
| Simvastatin | 9 | 5 | 4 |
| Atorvastatin | 4 | 3 | 0 |
| Ezetimibe | 11 | 6 | 3 |
| Cardiovascular disease | |||
| History of CAD | 16 | 11 | 8 |
| Arterial hypertension | 10 | 11 | 6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmöcker, C.; Zhang, I.W.; Kiesler, S.; Kassner, U.; Ostermann, A.I.; Steinhagen-Thiessen, E.; Schebb, N.H.; Weylandt, K.-H. Effect of Omega-3 Fatty Acid Supplementation on Oxylipins in a Routine Clinical Setting. Int. J. Mol. Sci. 2018, 19, 180. https://doi.org/10.3390/ijms19010180
Schmöcker C, Zhang IW, Kiesler S, Kassner U, Ostermann AI, Steinhagen-Thiessen E, Schebb NH, Weylandt K-H. Effect of Omega-3 Fatty Acid Supplementation on Oxylipins in a Routine Clinical Setting. International Journal of Molecular Sciences. 2018; 19(1):180. https://doi.org/10.3390/ijms19010180
Chicago/Turabian StyleSchmöcker, Christoph, Ingrid W. Zhang, Stefanie Kiesler, Ursula Kassner, Annika I. Ostermann, Elisabeth Steinhagen-Thiessen, Nils H. Schebb, and Karsten-H. Weylandt. 2018. "Effect of Omega-3 Fatty Acid Supplementation on Oxylipins in a Routine Clinical Setting" International Journal of Molecular Sciences 19, no. 1: 180. https://doi.org/10.3390/ijms19010180
APA StyleSchmöcker, C., Zhang, I. W., Kiesler, S., Kassner, U., Ostermann, A. I., Steinhagen-Thiessen, E., Schebb, N. H., & Weylandt, K. -H. (2018). Effect of Omega-3 Fatty Acid Supplementation on Oxylipins in a Routine Clinical Setting. International Journal of Molecular Sciences, 19(1), 180. https://doi.org/10.3390/ijms19010180