Antidiabetic Potential of Monoterpenes: A Case of Small Molecules Punching above Their Weight

Abstract
:1. Introduction
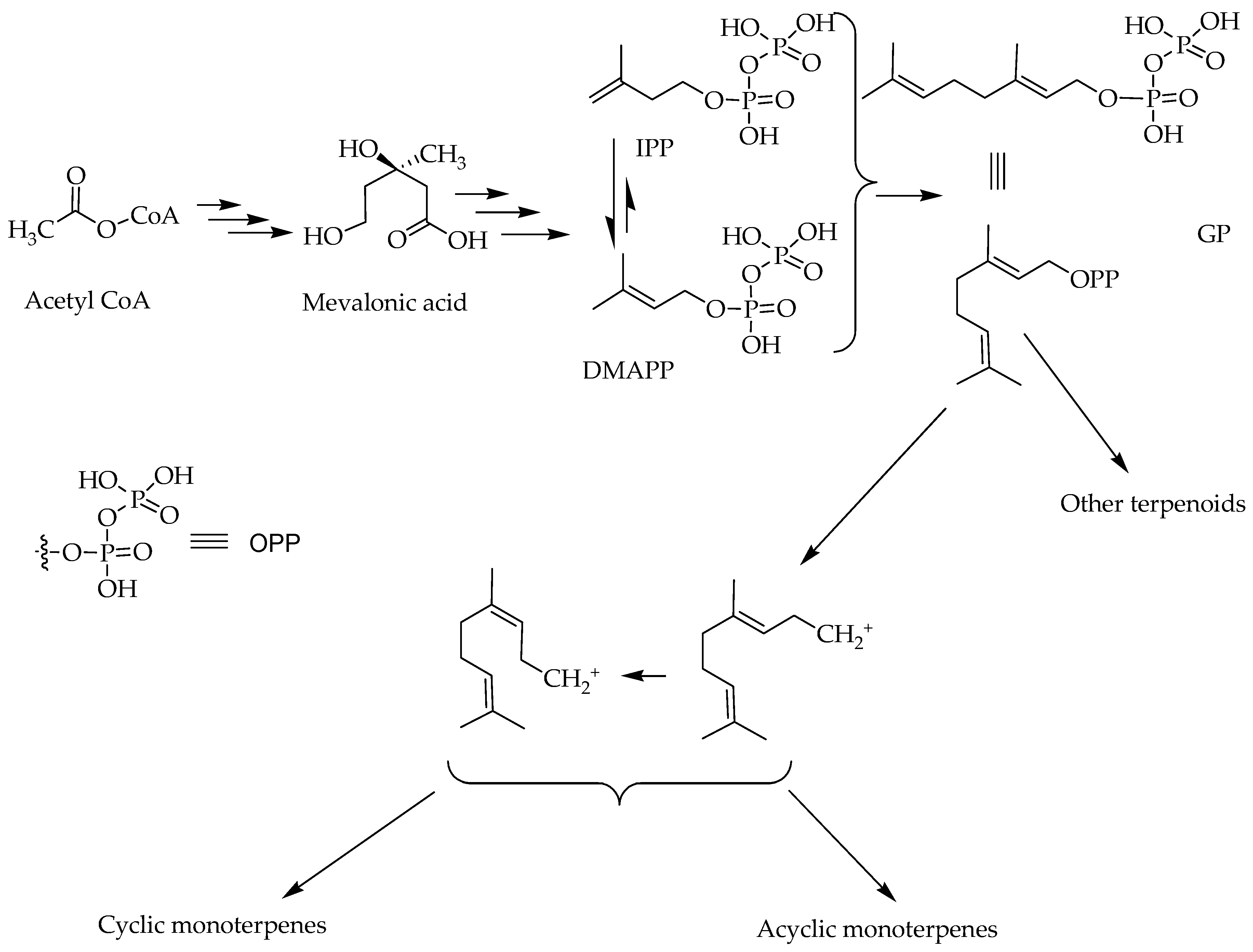
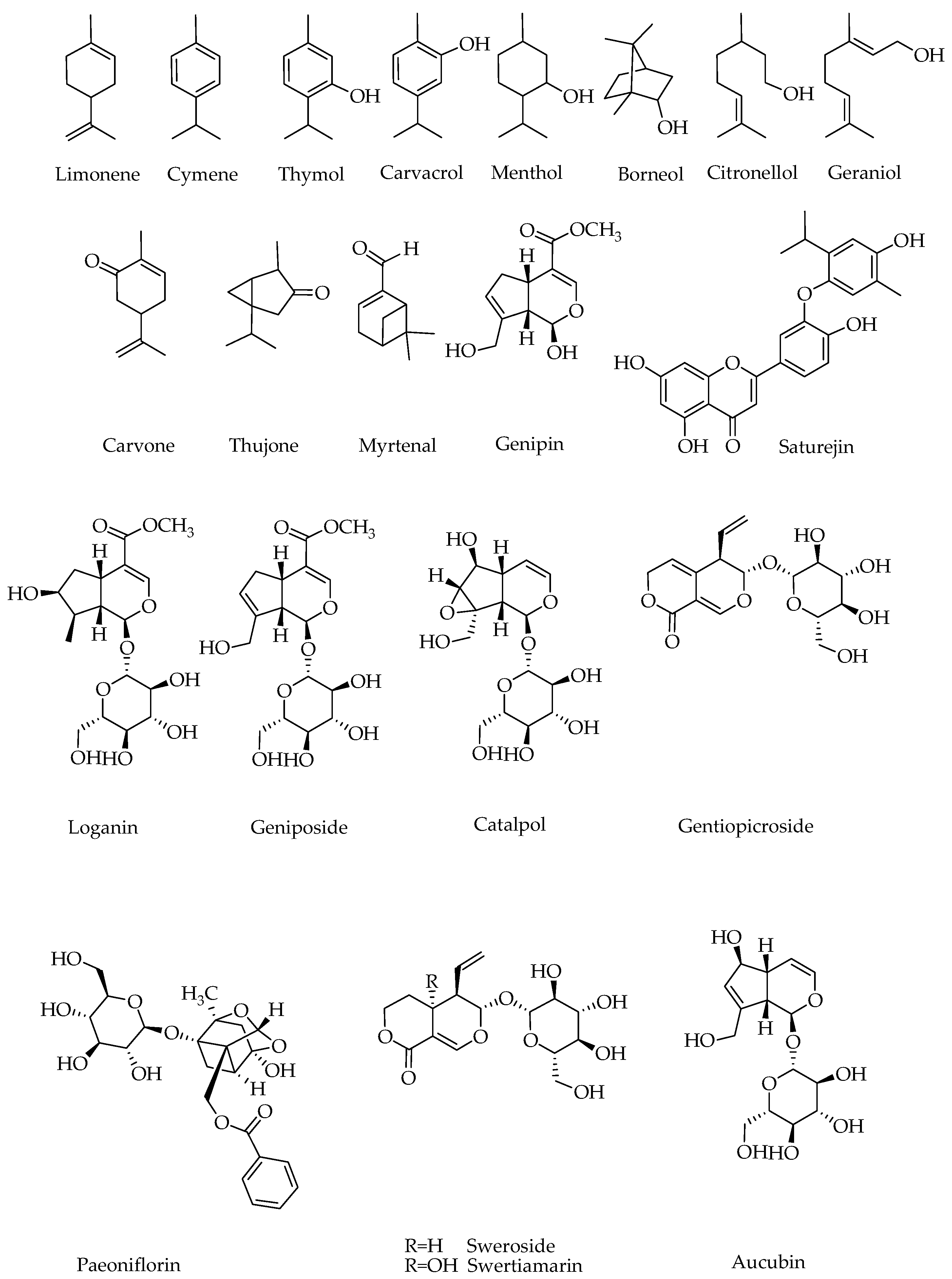
2. Chemistry
3. General Function in Nature
4. Antidiabetic Potential of Monoterpenes
4.1. In Vitro Protective Effects
4.2. In Vivo Antidiabetic Effects
5. Bioavailability
6. General Summary and Discussion
Conflicts of Interest
Abbreviations
| Aβ | Amyloid Beta |
| AMPK | Adenosine Monophosphate-Activated Protein Kinase |
| AKT | Protein Kinase B (PKB) |
| AGE | Advanced Glycation End Products |
| ALP | Alkaline Phosphatase |
| ALT | Alanine Transaminase |
| AST | Aspartate Aminotransferase |
| BACE1 | Beta-Secretase 1 |
| CAT | Catalase |
| CYP2E1 | Cytochrome P450 2E1 |
| DMAPP | Dimethylallyl Diphosphate |
| DPPH | 2,2-Diphenyl-1-Picrylhydrazyl |
| EGTA | Ethylene Glycol-bis(β-Aminoethyl Ether)-N,N,N′,N′-Tetraacetic Acid |
| Erk | Extracellular Signal–Regulated Kinases |
| HFD | High Fat Diet |
| HDL | High Density Lipoproteins |
| HMG-CoAR | 3-Hydroxy-3-Methyl Glutaryl Coenzyme A Reductase |
| FFAs | Free fatty acids |
| FoxO1 | Forkhead Box O1 |
| GLP-1R | Glucagon-Like-1 Receptor |
| GLUT | Glucose Transporter |
| GPx | Glutathione Peroxidase |
| Grb10 | Growth Factor Receptor Bound Protein 10 |
| GSH | Glutathione—Reduced Form |
| GSK3β | Glycogen Synthase Kinase 3 Beta |
| HbA1c | Haemoglobin A1c |
| HUVECs | Human Umbilical Vein Endothelial Cells |
| IDE | Insulin-Degrading Enzyme |
| IGF-1 | Insulin-Like Growth Factor-1 |
| IRS | Insulin Receptor Substrate |
| IPP | Isopentenyl Diphosphate |
| IL | Interleukin |
| JNK | c-Jun N-terminal kinase |
| LDH | Lactate Dehydrogenase |
| LDL | Low Density Lipoproteins-Cholesterol |
| MCP-1 | Monocyte Chemoattractant Protein-1 |
| MDA | Malondialdehyde |
| MMP | Mitochondrial Membrane Potential |
| NF-κB | Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells |
| Pck1 | Phosphoenolpyruvate Carboxykinase 1 |
| PI3K | Phosphatidylinositide 3-Kinase |
| PDK1 | 3-Phosphoinositide-Dependent Protein Kinase-1 |
| PDX-1 | Pancreatic and Duodenal Homeobox 1 or Insulin Promoter Factor 1 |
| PEPCK | Phosphoenolpyruvate Carboxykinase |
| PPAR | Peroxisome Proliferator-Activated Receptor |
| ROS | Reactive Oxygen Species |
| SOD | Superoxide Dismutase |
| T1D | Type-1 Diabetes |
| T2D | Type-2 Diabetes |
| TC | Total Cholesterol |
| TG | Triglycerides |
| TNF | Tumor Necrosis Factor |
| VLDL | Very Low Density Lipoproteins |
References
- WHO. Diabetes. Available online: http://www.who.int/mediacentre/factsheets/fs312/en/ (accessed on 15 August 2017).
- WHO. Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 15 August 2017).
- McCall, A.L.; Farhy, L.S. Treating type 1 diabetes: From strategies for insulin delivery to dual hormonal control. Minerva Endocrinol. 2013, 38, 145–163. [Google Scholar] [PubMed]
- Swinnen, S.G.; Hoekstra, J.B.; DeVries, J.H. Insulin therapy for Type 2 diabetes. Diabetes Care 2009, 32, S253–S259. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, J.; Polyzos, S.A.; Perakakis, N.; Thakkar, B.; Paschou, S.A.; Katsiki, N.; Underwood, P.; Park, K.-H.; Seufert, J.; Kang, E.S.; et al. Pharmacotherapy of type 2 diabetes: An update. Metabolism 2018, 78, 13–42. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, A.; Duvoor, C.; Dendi, V.S.R.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, S.N.; Montales, M.T.; Kuriakose, K.; et al. Clinical review of antidiabetic drugs: Implications for type 2 diabetes mellitus management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Stein, S.A.; Lamos, E.M.; Davis, S.N. A review of the efficacy and safety of oral antidiabetic drugs. Expert Opin. Drug. Saf. 2013, 12, 153–175. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Going back to the good old days: The merit of crude plant drug mixtures in the 21st century. Int. J. Complement. Altern. Med. 2017, 6, 1–5. [Google Scholar] [CrossRef]
- Habtemariam, S.; Varghese, G.K. Antioxidant, anti-α-glucosidase and pancreatic β-cell protective effects of methanolic extract of Ensete superbum Cheesm seeds. Asian Pac. J. Trop. Biomed. 2017, 7, 121–125. [Google Scholar] [CrossRef]
- Habtemariam, S.; Lentini, G. The therapeutic potential of rutin for diabetes: An update. Mini Rev. Med. Chem. 2015, 15, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S.; Varghese, G.K. The antidiabetic therapeutic potential of dietary polyphenols. Curr. Pharm. Biotechnol. 2014, 15, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. The biosynthesis of C5–C25 terpenoid compounds. Nat. Prod. Rep. 2002, 19, 181–222. [Google Scholar] [CrossRef] [PubMed]
- Rehman, R.; Hanif, M.A.; Mushtaq, Z.; Al-Sadi, A.M. Biosynthesis of essential oils in aromatic plants: A review. Food Rev. Int. 2016, 32, 117–160. [Google Scholar] [CrossRef]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Dinda, B.; Debnath, S.; Banik, R. Naturally occurring iridoids and secoiridoids. An updated review, Part 4. Chem. Pharm. Bull. 2011, 59, 803–833. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Llusià, J.; Estiarte, M.; Peñuelas, J. Terpenoids and plant communication. Butll. Inst. Catalana d'Hist. Nat. 1996, 64, 125–133. [Google Scholar]
- Singh, B.; Sharma, R.A. Plant terpenes: Defense responses, phylogenetic analysis, regulation and clinical applications. Biotechnology 2015, 5, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Thoppil, R.J.; Bishayee, A. Terpenoids as potential chemopreventive and therapeutic agents in liver cancer. World J. Hepatol. 2011, 3, 228–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Tang, W.; Bidigare, R.R. Terpenoids as therapeutic drugs and pharmaceutical agents. In Natural Products; Zhang, L., Demain, A.L., Eds.; ACS Publications: Washington, DC, USA, 2005; pp. 197–227. [Google Scholar]
- Dambolena, J.S.; Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Areco, V.A.; Zygadlo, J.A. Terpenes: Natural products for controlling insects of importance to human Health—A structure-activity relationship study. Psyche J. Entomol. 2016, 2016, 4595823. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; de Martino, L.; Coppola, R.; de Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Boskabady, M.H.; Jandaghi, P. Relaxant effects of carvacrol on guinea pig tracheal chains and its possible mechanisms. Pharmazie 2003, 58, 661–663. [Google Scholar] [PubMed]
- Eccles, R.; Jones, A.S. The effect of menthol on nasal resistance to air flow. J. Laryngol. Otol. 1983, 97, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Burrow, A.; Eccles, R.; Jones, A.S. The effects of camphor, eucalyptus and menthol vapour on nasal resistance to airflow and nasal sensation. Acta Otolaryngol. 1983, 96, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Brum, L.F.S.; Elisabetsky, E.; Souza, D. Effects of linalool on [3H]MK801 and [3H] muscimol binding in mouse cortical membranes. Phytother. Res. 2001, 15, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Park, T.J.; Park, Y.S.; Lee, T.G.; Ha, H.; Kim, KT. Inhibition of acetylcholine-mediated effects by borneol. Biochem. Pharmacol. 2003, 65, 83–90. [Google Scholar] [CrossRef]
- Dagli Gul, A.S.; Fadillioglu, E.; Karabulut, I.; Yesilyurt, A.; Delibasi, T. The effects of oral carvacrol treatment against H2O2 induced injury on isolated pancreas islet cells of rats. Islets 2013, 5, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, M.M.; Panaskar, S.N.; Arvindekar, A.U. Inhibition of advanced glycation end product formation by cymene—A common food constituent. J. Funct. Foods 2014, 6, 107–115. [Google Scholar] [CrossRef]
- Ma, C.J.; Nie, A.F.; Zhang, Z.J.; Zhang, Z.G.; Du, L.; Li, X.Y.; Ning, G. Genipin stimulates glucose transport in C2C12 myotubes via an IRS-1 and calcium-dependent mechanism. J. Endocrinol. 2013, 216, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Liu, C.; Yin, F.; Zhang, Y.; Liu, J. 5′-AMP-activated protein kinase plays an essential role in geniposide-regulated glucose-stimulated insulin secretion in rat pancreatic INS-1 β cells. J. Nat. Med. 2017, 71, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Hao, Y.N.; Yin, F.; Zhang, Y.L.; Liu, J.H. Geniposide accelerates proteasome degradation of Txnip to inhibit insulin secretion in pancreatic β-cells. J. Endocrinol. Investig. 2017, 40, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ding, Y.; Zhong, X.; Guo, Q.; Wang, H.; Gao, J.; Bai, T.; Ren, L.; Guo, Y.; Jiao, X.; et al. Geniposide acutely stimulates insulin secretion in pancreatic β-cells by regulating GLP-1 receptor/cAMP signaling and ion channels. Mol. Cell Endocrinol. 2016, 430, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, F.; Liu, J.; Liu, Z.; Guo, L.; Xia, Z.; Zidichouski, J. Geniposide attenuates insulin-deficiency-induced acceleration of β-amyloidosis in an APP/PS1 transgenic model of Alzheimer's disease. Neurochem. Int. 2015, 89, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Liu, J.H.; Yin, F. Regulation of insulin secretion by geniposide: Possible involvement of phosphatidylinositol 3-phosphate kinase. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 1287–1294. [Google Scholar] [PubMed]
- Liu, J.; Zhang, Y.; Deng, X.; Yin, F. Geniposide decreases the level of Aβ1-42 in the hippocampus of streptozotocin-induced diabetic rats. Acta Biochim. Biophys. Sin. 2013, 45, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yin, F.; Xiao, H.; Guo, L.; Gao, X. Glucagon-like peptide 1 receptor plays an essential role in geniposide attenuating lipotoxicity-induced β-cell apoptosis. Toxicol. In Vitro 2012, 26, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Xia, Z.N.; Gao, X.; Yin, F.; Liu, J.H. Glucagon-like peptide 1 receptor plays a critical role in geniposide-regulated insulin secretion in INS-1 cells. Acta Pharmacol. Sin. 2012, 33, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Kojima, K.; Shimada, T.; Nagareda, Y.; Watanabe, M.; Ishizaki, J.; Sai, Y.; Miyamoto, K.; Aburada, M. Preventive effect of geniposide on metabolic disease status in spontaneously obese type 2 diabetic mice and free fatty acid-treated HepG2 cells. Biol. Pharm. Bull. 2011, 34, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.J.; Li, J.; Mei, Z.Y.; Chen, G. Gentiopicroside and sweroside from Veratrilla baillonii Franch. induce phosphorylation of Akt and suppress Pck1 expression in hepatoma cells. Biochem. Cell Biol. 2016, 94, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Chi, R.; Zhang, L.; Wang, N.; Lu, Y. Effects of paeoniflorin on tumor necrosis factor-α-induced insulin resistance and changes of adipokines in 3T3-L1 adipocytes. Fitoterapia 2013, 91, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Qiao, J.; Yang, Y.; Lu, Y. Inhibitory effect of paeoniflorin on the inflammatory vicious cycle between adipocytes and macrophages. J. Cell. Biochem. 2012, 113, 2560–2566. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.T.; Trung, T.N.; Hien, T.T.; Dao, T.T.; Yim, N.; Ngoc, T.M.; Oh, W.K.; Bae, K. Selected compounds derived from MoutanCortex stimulated glucose uptake and glycogen synthesis via AMPK activation in human HepG2 cells. J. Ethnopharmacol. 2010, 131, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.C.; Chua, K.H.; Ravishankar-Ram, M.; Kuppusamy, U.R. Monoterpenes: Novel insights into their biological effects and roles on glucose uptake and lipid metabolism in 3T3-L1 adipocytes. Food Chem. 2016, 196, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Malmir, M.; Gohari, A.R.; Saeidnia, S.; Silva, O. A new bioactive monoterpene–flavonoid from Satureja khuzistanica. Fitoterapia 2015, 105, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.P.; Rawal, K.; Soni, S.; Gupta, S. Swertiamarin ameliorates oleic acid induced lipid accumulation and oxidative stress by attenuating gluconeogenesis and lipogenesis in hepatic steatosis. Biomed. Pharmacother. 2016, 83, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Alkhateeb, H.; Bonen, A. Thujone, a component of medicinal herbs, rescues palmitate-induced insulin resistance in skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R804–R812. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S.; Varghese, G.K. A novel diterpene skeleton: Identification of a highly aromatic, cytotoxic and antioxidant 5-methyl-10-demethyl-abietane-type diterpene from Premna serratifolia. Phyther. Res. 2015, 29, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Investigation into the antioxidant and antidiabetic potential of Moringa stenopetala: Identification of the active principles. Nat. Prod. Commun. 2015, 10, 475–478. [Google Scholar] [PubMed]
- Habtemariam, S.; Varghese, G.K. Extractability of rutin in herbal tea preparations of Moringa stenopetala leaves. Beverages 2015, 1, 169–182. [Google Scholar] [CrossRef]
- Roselli, M.; Lentini, G.; Habtemariam, S. Phytochemical, antioxidant and anti-α-glucosidase activity evaluations of Bergenia cordifolia. Phyther. Res. 2012, 26, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S.; Cowley, R.A. Antioxidant and anti-α-glucosidase ccompounds from the rhizome of Peltiphyllum peltatum (Torr.) Engl. Phytother. Res. 2012, 26, 1656–1660. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Methyl-3-O-methyl gallate and gallic acid from the leaves of Peltiphyllum peltatum: Isolation and comparative antioxidant, prooxidant, and cytotoxic effects in neuronal cells. J. Med. Food 2011, 14, 1412–1418. [Google Scholar] [CrossRef] [PubMed]
- Juan-Badaturuge, M.; Habtemariam, S.; Thomas, M.J.K. Antioxidant compounds from a South Asian beverage and medicinal plant, Cassia auriculata. Food Chem. 2011, 125, 221–225. [Google Scholar] [CrossRef]
- Juan-Badaturugea, M.; Habtemariam, S.; Jackson, C.; Thomas, M.J.K. Antioxidant principles of Tanacetum vulgare L. aerial part. Nat. Prod. Commun. 2009, 4, 1561–1564. [Google Scholar]
- Habtemariam, S.; Dagne, E. Comparative antioxidant, prooxidant, and cytotoxic activity of sigmoidin A and eriodictyol. Planta Med. 2010, 76, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Activity-guided isolation and identification of free Radical-scavenging components from ethanolic extract of Boneset (Leaves of Eupatorium perfoliatum). Nat. Prod. Commun. 2008, 3, 1317–1320. [Google Scholar]
- Habtemariam, S.; Jackson, C. Antioxidant and cytoprotective activity of leaves of Peltiphyllum peltatum (Torr.) Engl. Food Chem. 2007, 105, 498–503. [Google Scholar] [CrossRef]
- Habtemariam, S. Flavonoids as inhibitors or enhancers of the cytotoxicity of tumor necrosis factor-α in l-929 tumor cells. J. Nat. Prod. 1997, 60, 775–778. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Modulation of tumour necrosis factor-α-induced cytotoxicity by polyphenols. Phyther. Res. 1997, 11, 277–280. [Google Scholar] [CrossRef]
- Habtemariam, S. Catechols and quercetin reduce MTT through iron ions: A possible artefact in cell viability assay. Phyther. Res. 1995, 9, 603–605. [Google Scholar] [CrossRef]
- Varghese, G.K.; Bose, L.V.; Habtemariam, S. Antidiabetic components of Cassia alata leaves: Identification through α-glucosidase inhibition studies. Pharm. Biol. 2013, 51, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Antihyperlipidemic components of Cassia auriculata aerial parts: Identification through in vitro studies. Phytother. Res. 2013, 27, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. α-Glucosidase inhibitory activity of kaempferol-3-O-rutinoside. Nat. Prod. Commun. 2011, 6, 201–203. [Google Scholar] [PubMed]
- Jin, L.; Xue, H.Y.; Jin, L.J.; Li, S.Y.; Xu, Y.P. Antioxidant and pancreas-protective effect of aucubin on rats with streptozotocin-induced diabetes. Eur. J. Pharmacol. 2008, 582, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Kodikonda, M.; Naik, P.R. Ameliorative effect of borneol, a natural bycyclic monoterpene against hyperglycemia, hyperlipidemia and oxidative stress in streptozotocin-induced diabetic Wistar rats. Biomed. Pharmacother. 2017, 96, 336–347. [Google Scholar] [CrossRef]
- Ezhumalai, M.; Ashokkumar, N.; Pugalendi, K.V. Combination of carvacrol and rosiglitazone ameliorates high fat diet induced changes in lipids and inflammatory markers in C57BL/6J mice. Biochimie 2015, 110, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Ezhumalai, M.; Radhiga, T.; Pugalendi, K.V. Antihyperglycemic effect of carvacrol in combination with rosiglitazone in high-fat diet-induced type 2 diabetic C57BL/6J mice. Mol. Cell. Biochem. 2014, 385, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Lu, H.; Teng, J. Carvacrol attenuates diabetes-associated cognitive deficits in rats. J. Mol. Neurosci. 2013, 51, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Farkhondeh, T.; Samini, F.; Borji, A. Protective Effects of Carvacrol against Oxidative Stress Induced by Chronic Stress in Rat’s Brain, Liver, and Kidney. Biochem. Res. Int. 2016, 2016, 2645237. [Google Scholar] [CrossRef] [PubMed]
- Bayramoglu, G.; Senturk, H.; Bayramoglu, A.; Uyanoglu, M.; Colak, S.; Ozmen, A.; Kolankaya, D. Carvacrol partially reverses symptoms of diabetes in STZ-induced diabetic rats. Cytotechnology 2014, 66, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Muruganathan, U.; Srinivasan, S. Beneficial effect of carvone, a dietary monoterpene ameliorates hyperglycemia by regulating the key enzymes activities of carbohydrate metabolism in streptozotocin-induced diabetic rats. Biomed. Pharmacother. 2016, 84, 1558–1567. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Deng, H.; Zhang, Q.; Xie, J.; Zeng, H.; Jin, X.; Ling, Z.; Shan, Q.; Liu, M.; Ma, Y.; et al. Amelioration of diabetic mouse nephropathy by catalpol correlates with down-regulation of Grb10 expression and activation of insulin-like growth factor 1/insulin-like growth factor 1 receptor signaling. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, G.; Ma, S.; Li, F.; Yuan, M.; Xu, H.; Huang, K. Catalpol ameliorates high-fat diet-induced insulin resistance and adipose tissue inflammation by suppressing the JNK and NF-κB pathways. Biochem. Biophys. Res. Commun. 2015, 467, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.J.; Niu, H.S.; Lin, M.H.; Cheng, J.T.; Hsu, F.L. Antihyperglycemic effect of catalpol in streptozotocin-induced diabetic rats. J. Nat. Prod. 2010, 73, 1170–1172. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.F.; Li, D.Q.; Xue, H.Y.; Hu, B. Oral supplementation of catalpol ameliorates diabetic encephalopathy in rats. Brain Res. 2010, 1307, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Muruganathan, U. Antidiabetic efficacy of citronellol, a citrus monoterpene by ameliorating the hepatic key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2016, 250, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Feng, H.; Gong, D.; Zhao, X.; Cai, L.; Wu, Q.; Yuan, B.; Yang, M.; Zhao, J.; Zou, Y. Genipin ameliorates age-related insulin resistance through inhibiting hepatic oxidative stress and mitochondrial dysfunction. Exp. Gerontol. 2013, 48, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, F.; Liu, J.; Liu, Z. Geniposide Attenuates the phosphorylation of tau protein in cellular and insulin-deficient APP/PS1 transgenic mouse model of Alzheimer’s disease. Chem. Biol. Drug Des. 2016, 87, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Liu, Y.; Jiang, Y.; Ding, J.; Li, L. Geniposide ameliorates learning memory deficits, reduces tau phosphorylation and decreases apoptosis via GSK3β pathway in streptozotocin-induced Alzheimer rat model. Brain Pathol. 2014, 24, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, B.; Yu, B. Paeoniflorin protects against nonalcoholic fatty liver disease induced by a high-fat diet in mice. Biol. Pharm. Bull. 2015, 38, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Huang, C.; Zong, G.; Zha, D.; Meng, X.; Li, J.; Tang, W. Hepatoprotective effects of geniposide in a rat model of nonalcoholic steatohepatitis. J. Pharm. Pharmacol. 2011, 63, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Wang, G.F.; Liu, Z.Q.; Rao, J.J.; Lü, L.; Xu, W.; Wu, S.G.; Zhang, J.J. Effect of geniposide, a hypoglycemic glucoside, on hepatic regulating enzymes in diabetic mice induced by a high-fat diet and streptozotocin. Acta Pharmacol. Sin. 2009, 30, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Babukumar, S.; Vinothkumar, V.; Sankaranarayanan, C.; Srinivasan, S. Geraniol, a natural monoterpene, ameliorates hyperglycemia by attenuating the key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Pharm. Biol. 2017, 55, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Murali, R.; Saravanan, R. Antidiabetic effect of d-limonene, a monoterpene in streptozotocin-induced diabetic rats. Biomed. Prevent. Nutr. 2012, 2, 269–275. [Google Scholar] [CrossRef]
- Bacanlı, M.; Anlar, H.G.; Aydın, S.; Çal, T.; Arı, N.; Ündeğer Bucurgat, Ü.; Başaran, A.A.; Başaran, N. d-limonene ameliorates diabetes and its complications in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2017, 110, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Xu, H.; Lv, G.; Liu, B.; Lee, M.K.; Lu, C.; Lv, X.; Wu, Y. Loganin attenuates diabetic nephropathy in C57BL/6J mice with diabetes induced by streptozotocin and fed with diets containing high level of advanced glycation end products. Life Sci. 2015, 123, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Muruganathan, U.; Srinivasan, S.; Vinothkumar, V. Antidiabetogenic efficiency of menthol, improves glucose homeostasis and attenuates pancreatic β-cell apoptosis in streptozotocin-nicotinamide induced experimental rats through ameliorating glucose metabolic enzymes. Biomed Pharmacother. 2017, 92, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, A.; Pari, L. Myrtenal ameliorates hyperglycemia by enhancing GLUT2 through Akt in the skeletal muscle and liver of diabetic rats. Chem. Biol. Interact. 2016, 256, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, A.; Pari, L.; Chandramohan, R.; Sheikh, B.A. Histopathological findings of the pancreas, liver, and carbohydrate metabolizing enzymes in STZ-induced diabetic rats improved by administration of myrtenal. J. Physiol. Biochem. 2014, 70, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Ayyasamy, R.; Leelavinothan, P. Myrtenal alleviates hyperglycaemia, hyperlipidaemia and improves pancreatic insulin level in STZ-induced diabetic rats. Pharm. Biol. 2016, 54, 2521–2527. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, S.; Xu, L.; Wang, H.; Ma, Z.; Fu, Q.; Qu, R.; Ma, S. Paeoniflorin ameliorates cognitive dysfunction via regulating SOCS2/IRS-1 pathway in diabetic rats. Physiol. Behav. 2017, 174, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, H.; Prajapati, A.; Rajani, M.; Sudarsanam, V.; Padh, H.; Goyal, R.K. Beneficial Effects of Swertiamarin on dyslipidaemia in streptozotocin-induced Type 2 diabetic rats. Phytother. Res. 2012, 26, 1259–1261. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, S.; Pari, L. Role of thymol on hyperglycemia and hyperlipidemia in high fat diet-induced type 2 diabetic C57BL/6J mice. Eur. J. Pharmacol. 2015, 761, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, S.; Pari, L. Protective effect of thymol on high fat diet induced diabetic nephropathy in C57BL/6J mice. Chem. Biol. Interact. 2016, 245, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Li, H.; Qin, T.; Li, M.; Ma, S. Thymol improves high-fat diet-induced cognitive deficits in mice via ameliorating brain insulin resistance and upregulating NRF2/HO-1 pathway. Metab. Brain Dis. 2017, 32, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.R.; Ansari, S.H.; Najmi, A.K.; Ahmad, M.A. Monoterpene phenolic compound thymol prevents high fat diet induced obesity in murine model. Toxicol. Mech. Methods 2014, 24, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Jiang, Z.H.; Liu, L.; Hu, M. Mechanisms responsible for poor oral bioavailability of paeoniflorin: Role of intestinal disposition and interactions with sinomenine. Pharm. Res. 2006, 23, 2768–2780. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Lin, J.-Z.; Li, L.; Yang, J.-L.; Jia, W.-W.; Huang, W.-H.; Du, F.-F.; Wang, F.-Q.; Li, M.-J.; Li, Y.F.; et al. Pharmacokinetics and disposition of monoterpene glycosides derived from Paeonia lactiflora roots (Chishao) after intravenous dosing of antiseptic XueBiJing injection in human subjects and rats. Acta Pharmacol. Sin. 2016, 37, 530–544. [Google Scholar] [CrossRef] [PubMed]
- Martey, O.N.K.; Shi, X.; He, X. Advance in pre-clinical pharmacokinetics of paeoniflorin, a major monoterpene glucoside from the root of Paeonia lactiflora. Pharmacol. Pharm. 2013, 4, 4–14. [Google Scholar] [CrossRef]
- Austgulen, L.T.; Solheim, E.; Scheline, R.R. Metabolism in rats of p-cymene derivatives: Carvacrol and thymol. Pharmacol. Toxicol. 1987, 61, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.H.; Fang, Z.Z.; Zhu, L.L.; Liang, S.C.; Ge, G.B.; Liu, Z.Y. Investigation of UDP-glucuronosyltransferases (UGTs) inhibitory properties of carvacrol. Phytother. Res. 2012, 26, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Kohlert, C.; Schindler, G.; März, R.W.; Abel, G.; Brinkhaus, B.; Derendorf, H.; Gräfe, E.U.; Veit, M. Systemic availability and pharmacokinetics of thymol in humans. J. Clin. Pharmacol. 2002, 42, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.A.; Lang, J.E.; Ley, M.; Nagle, R.; Hsu, C.H.; Thompson, P.A.; Cordova, C.; Waer, A.; Chow, H.H. Human breast tissue disposition and bioactivity of limonene in women with early stage breast cancer. Cancer Prev. Res. 2013, 6, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.A.; Hakim, I.A.; Chew, W.; Thompson, P.; Thomson, C.A.; Chow, H.H. Adipose tissue accumulation of d-limonene with the consumption of a lemonade preparation rich in d-limonene content. Nutr. Cancer 2010, 62, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; He, J.C.; Bai, M.; Song, Q.Y.; Feng, E.F.; Rao, G.X.; Xu, G.L. Determination of the plasma pharmacokinetic and tissue distributions of swertiamarin in rats by liquid chromatography with tandem mass spectrometry. Arzneimittelforschung 2012, 62, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Lin, L.C.; Lin, C.H.; Tsai, T.H. Comparative oral bioavailability of geniposide following oral administration of geniposide, Gardenia jasminoides Ellis fruits extracts and Gardenia herbal formulation in rats. J. Pharm. Pharmacol. 2014, 66, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Eldor, R.; DeFronzo, R.A.; Abdul-Ghani, M. In vivo actions of peroxisome proliferator-activated receptors: Glycemic control, insulin sensitivity, and insulin secretion. Diabetes Care 2013, 36 (Suppl. 2), S162–S174. [Google Scholar] [CrossRef] [PubMed]
- Soccio, R.E.; Chen, E.R.; Rajapurkar, S.R.; Safabakhsh, P.; Marinis, J.M.; Dispirito, J.R.; Emmett, M.J.; Briggs, E.R.; Fang, B.; Everett, L.J.; et al. Genetic variation determines PPARγ function and anti-diabetic drug response in vivo. Cell 2015, 162, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Delea, T.E.; Edelsberg, J.S.; Hagiwara, M.; Oster, G.; Phillips, L.S. Use of thiazolidinediones and risk of heart failure in people with type 2 diabetes: A retrospective cohort study. Diabetes Care 2003, 26, 2983–2989. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Snaith, J.; Gunton, J.E. Diabetes Medications and Cardiovascular Outcomes in Type 2 Diabetes. Heart Lung Circ. 2017, 26, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H.; et al. Natural product agonists of peroxisome proliferator-activated receptor γ (PPARγ): A review. Biochem. Pharmacol. 2014, 92, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Wang, J.N.; Gu, S.P.; Bu, J.; Kramer, M.P.; Baumgartner, L.; Fakhrudin, N.; Ladurner, A.; Malainer, C.; Vuorinen, A.; et al. Honokiol: A nonadipogenic PPARγ agonist from nature. Biochim. Biophys. Acta 2013, 1830, 4813–4819. [Google Scholar] [CrossRef] [PubMed]
- Chopra, I.; Li, H.F.; Wang, H.; Webster, K.A. Phosphorylation of the insulin receptor by AMP-activated protein kinase (AMPK) promotes ligand-independent activation of the insulin signalling pathway in rodent muscle. Diabetologia 2012, 55, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Cao, J.; He, Q.; Xiong, L.; Chang, E.; Radovick, S.; Wondisford, F.E.; He, L. Metformin activates AMP-activated protein kinase by promoting formation of the αβγ heterotrimeric complex. J. Biol. Chem. 2015, 290, 3793–3802. [Google Scholar] [CrossRef] [PubMed]
- Gaidhu, M.P.; Fediuc, S.; Anthony, N.M.; So, M.; Mirpourian, M.; Perry, R.L.; Ceddia, R.B. Prolonged AICAR-induced AMP-kinase activation promotes energy dissipation in white adipocytes: Novel mechanisms integrating HSL and ATGL. J. Lipid Res. 2009, 50, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; You, Y.H.; Ham, D.S.; Yang, H.K.; Yoon, K.H. The paradoxical effects of AMPK on insulin gene expression and glucose-induced insulin secretion. J. Cell Biochem. 2016, 117, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Yavari, A.; Stocker, C.J.; Ghaffari, S.; Wargent, E.T.; Steeples, V.; Czibik, G.; Pinter, K.; Bellahcene, M.; Woods, A.; Martínez de Morentin, P.B.; et al. Chronic activation of γ2 AMPK induces obesity and reduces β cell function. Cell Metab. 2016, 23, 821–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhong, L.; Wang, F.; Zhu, G. Dissecting the role of AMP-activated protein kinase in human diseases. Acta Pharm. Sin. B 2017, 7, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Belwal, T.; Nabavi, S.F.; Nabavi, S.M.; Habtemariam, S. Dietary anthocyanins and insulin resistance: When food becomes a medicine. Nutrients 2017, 9, 1111. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, B.J.; Griesel, B.A.; King, C.D.; Josey, M.A.; Olson, A.L. Moderate GLUT4 overexpression improves insulin sensitivity and fasting triglyceridemia in high-fat diet–fed transgenic mice. Diabetes 2013, 62, 2249–2258. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Czech, M.P. The GLUT4 Glucose Transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Morgan, B.J.; Chai, S.Y.; Albiston, A.L. GLUT4 associated proteins as therapeutic targets for diabetes. Recent Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Olson, A.L. Regulation of GLUT4 and insulin-dependent glucose flux. Int. Sch. Res. Not. 2012, 2012, 856987. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Lee, S.K.; Kim, J.H.; Kim, N.; You, G.Y.; Moon, J.W.; Kim, S.J.; Park, S.H.; Kim, H.S. Metformin regulates glucose transporter 4 (GLUT4) translocation through AMP-activated protein kinase (AMPK)-mediated Cbl/CAP signaling in 3T3-L1 preadipocyte cells. J. Biol. Chem. 2012, 287, 44121–44129. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.H.; Lee, H.A.; Park, M.H.; Han, J.S. Mulberry (Morus alba L.) fruit extract containing anthocyanins improves glycemic control and insulin sensitivity via activation of AMP-activated protein kinase in diabetic C57BL/Ksj-db/db mice. J. Med. Food 2016, 19, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Talagavadi, V.; Rapisarda, P.; Galvano, F.; Pelicci, P.; Giorgio, M. Cyanidin-3-O-β-glucoside and protocatechuic acid activate AMPK/mTOR/S6K pathway and improve glucose homeostasis in mice. J. Funct. Foods 2016, 21, 338–348. [Google Scholar] [CrossRef]
- Huang, B.; Wang, Z.; Park, J.H.; Ryu, O.H.; Choi, M.K.; Lee, J.Y. Anti-diabetic effect of purple corn extract on C57BL/KsJ db/db mice. Nutr. Res. Pract. 2015, 9, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zheng, X. Anthocyanin-rich mulberry fruit improves insulin resistance and protects hepatocytes against oxidative stress during hyperglycemia by regulating AMPK/ACC/mTOR pathway. J. Funct. Foods 2017, 30, 270–281. [Google Scholar] [CrossRef]
- Zhu, W.; Jia, Q.; Wang, Y.; Zhang, Y.; Xia, M. The anthocyanin cyanidin-3-O-β-glucoside, a flavonoid, increases hepatic glutathione synthesis and protects hepatocytes against reactive oxygen species during hyperglycemia: Involvement of a cAMP-PKA-dependent signaling pathway. Free Rad. Biol. Med. 2012, 52, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Barf, T. Intervention of hepatic glucose production. Small molecule regulators of potential targets for type 2 diabetes therapy. Mini Rev. Med. Chem. 2004, 4, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Agius, L. New hepatic targets for glycaemic control in diabetes. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Kurukulasuriya, R.; Link, J.T.; Madar, D.J.; Pei, Z.; Richards, S.J.; Rohde, J.J.; Souers, A.J.; Szczepankiewicz, B.G. Potential drug targets and progress towards pharmacologic inhibition of hepatic glucose production. Curr. Med. Chem. 2003, 10, 123–153. [Google Scholar] [CrossRef] [PubMed]
- McCormack, J.G.; Westergaard, N.; Kristiansen, M.; Brand, C.L.; Lau, J. Pharmacological approaches to inhibit endogenous glucose production as a means of anti-diabetic therapy. Curr. Pharm. Des. 2001, 7, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Luft, V.C.; Schmidt, M.I.; Pankow, J.S.; Couper, D.; Ballantyne, C.M.; Young, J.H.; Duncan, B.B. Chronic inflammation role in the obesity-diabetes association: A case-cohort study. Diabetol. Metab. Syndr. 2013, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.S.; Alvarez-Leite, J.I. Low-grade inflammation, obesity, and diabetes. Curr. Obes. Rep. 2014, 3, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Edirisinghe, I.; Banaszewski, K.; Cappozzo, J. Strawberry anthocyanin and its association with postprandial inflammation and insulin. Br. J. Nutr. 2011, 106, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Donath, M.Y.; Dalmas, É.; Sauter, N.S.; Böni-Schnetzler, M. Inflammation in obesity and diabetes: Islet dysfunction and therapeutic opportunity. Cell Metab. 2013, 17, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Zeidan, M.; Rayan, M.; Zeidan, N.; Falah, M.; Rayan, A. Indexing Natural Products for Their Potential Anti-Diabetic Activity: Filtering and Mapping Discriminative Physicochemical Properties. Molecules 2017, 22, 1563. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Compound | Model | Outcome | References |
|---|---|---|---|
| Carvacrol | H2O2-induced cellular injury on isolated pancreas islets—Following 20, 40 and 80 mg/kg/day in vivo treatment | Cytoprotective | [29] |
| Cymene | Advanced glycation end products (AGEs) | 100 μM—Inhibit AGE formation; inhibit glycation specific decline in BSA α-helix content and β-sheet. | [30] |
| Genipin | C2C12 myotubes | 10 μM—Stimulate glucose uptake; promote GLUT4 translocation; increase insulin receptor IRS-1, AKT, and GSK3β phosphorylation; increase ATP levels, close K(ATP) channels; increase intracellular calcium level; effect blocked by wortmannin and EGTA *. | [31] |
| Geniposide | Rat INS-1 pancreatic β cells | Prevent cell damage induced by high (25 mM) glucose through the AMPK pathway | [32,33] |
| Geniposide | Pancreatic β-cells—cultured primary cells of rats origin | 10 µM—Potentiate insulin secretion via activating the glucagon-like-1 receptor (GLP-1R) as well as the adenylyl cyclase (AC)/cAMP signaling pathway; inhibit voltage-dependent potassium channels; activate Ca2+ channels. | [34] |
| Geniposide | Primary cortical neurons; PC12 cells | Enhance PPARγ phosphorylation; accelerate the release of phosphorylated FoxO1 (forkhead box O1) from nuclear fraction to the cytosol; activate the activity of insulin-degrading enzyme promoter in PC12 cells | [35] |
| Geniposide | INS-1 pancreatic β cells | 10 μM—Increase phosphorylation of PDK1 and Akt473; inhibit the phosphorylation of downstream target GSK3β; increase expression of GLUT2; effect abolished by inhibitor of PI3K (LY294002). | [36] |
| Geniposide | INS-1 pancreatic β cells | Up to 10 μM—Enhance glucose-stimulated insulin secretion in response to low or moderately high glucose concentrations; promote glucose uptake and intracellular ATP levels; modulate pyruvate carboxylase expression. | [37] |
| Geniposide | Pancreatic INS-1 cells | Attenuate palmitate-induced β-cell apoptosis and caspase-3 expression; improve the impaired GLP-1R signaling by enhancing the phosphorylation of Akt and Foxo1; increase the expression of PDX-1; effect inhibited by exendin (9–39), an antagonist for GLP-1 receptor. | [38] |
| Geniposide | Pancreatic INS-1 cells | 10 μmol/L—Enhance acute insulin secretion in response to both the low (5.5 mmol/L) and moderately high levels (11 mmol/L) of glucose; Effect inhibited by GLP-1R antagonist exendin (9–39) or knock-down of GLP-1R with shRNA interference in INS-1 cells. | [39] |
| Geniposide | HepG2 fatty liver model- free fatty acid treatment | Suppress the intracellular lipid accumulation; increase the intracellular expression of a fatty acid oxidation-related gene (PPARα). | [40] |
| Gentiopicroside | HL1C hepatoma cells | 50 and 100 µM—Suppress Pck1 expression; induce phosphorylation of components in the insulin signaling cascade (Akt and Erk1/2 phosphorylation). | [41] |
| Paeoniflorin | 3T3-L1 adipocytes treated with tumour necrosis factor (TNF)-α | 50 µg/mL—Increase insulin-stimulated glucose; promote serine phosphorylation of IRS-1 and insulin-stimulated phosphorylation of AKT; inhibit the expressions and secretions of IL-6 and MCP-1; attenuate TNF-α-mediated suppression of the expressions of PPARγ and PPARγ target gene; effect reversed by antagonist of PPARγ activity. | [42] |
| Paeoniflorin | 3T3-L1 adipocytes and RAW 264.7 macrophages | 12.5–100 µg/mL—Inhibit TNF-α and FFA production; inhibit TNF-α-stimulated adipocyte lipolysis; suppress phosphorylation of TNF-α-activated ERK1/2; attenuate (partially) palmitate-induced macrophage TNF-α production. | [43] |
| Paeoniflorin derivatives (methoxyl and glucoside analogues) | Human HepG2 cells and HUVECs | 10 µM—Increase glucose uptake; reverse glucose-induced inhibition of glycogen synthesis in HepG2; increase AMPK and GSK-3β phosphorylation; phosphorylate AMPK and increase phosphorylation of GSK-3β while suppressing lipogenic expression (acetyl-CoA carboxylase and fatty acid synthase); induced eNOS phosphorylation in HUVECs. | [44] |
| (R)-(+)-limonene | 3T3-L1 cell culture; α-amylase and α-glucosidase enzymes | Increase GLUT1 expression at mRNA level; Weak enzyme inhibition (mM range). | [45] |
| Saturejin (3′-(2,5-dihydroxy-p-cymene) 5,7,4′-trihydroxy flavone) from Satureja khuzistanica Jamzad | Antioxidant activity; α- and β-glucosidase inhibitory | 10 μg/mL—Significant in vitro radical (DPPH) scavenging and enzyme inhibitory effects. | [46] |
| Sweroside | HL1C hepatoma cells | Suppress Pck1 expression and induce phosphorylation of components in the insulin signaling cascade (Akt and Erk1/2 phosphorylation). | [41] |
| Swertiamarin | Steatosis in HepG2 cells induced by 1 mM oleic acid | 25 μg/mL—Maintain membrane integrity; prevent apoptosis; increase the expressions of major insulin signaling proteins (insulin receptor, PI3K and pAkt) with concomitant reduction in p307 IRS-1; activate AMPK; modulate PPAR-α; decrease the levels of the gluconeogenic enzyme, PEPCK. | [47] |
| Thujone | Palmitate-induced insulin resistance in skeletal muscle (Soleus muscles) | Ameliorate palmitate oxidation and enhance insulin-stimulated glucose transport; restore (partially) GLUT4 translocation and AS160 phosphorylation; increase AMPK phosphorylation. | [48] |
| Compound | Model | Outcome | References |
|---|---|---|---|
| Aucubin | STZ-induced diabetic rats—5 mg/kg, i.p. twice daily for the first 5 days followed by single injections daily for 10 days. | Lower blood glucose; reverse lipid peroxidation and the decreased in activities of antioxidant enzymes in liver and kidneys; increase immunoreactive beta cells. | [66] |
| Borneol | STZ-induced diabetic rats—25 or 50 mg/kg, p.o. for 30 days. | Lower blood glucose and HbA1c; increase blood insulin; restore body weight loss; increase liver glycogen level; reverse the diabetes-induced increase in the levels of TC, TGs LDL-C, VLDL-C; restore urea and ALT and AST levels; increase antioxidant status (SOD, catalase, GSH) in the liver and kidney; reduce MDA level. | [67] |
| Carvacrol | HFD-induced C57BL/6J diabetic mice—20 mg/kg p.o. for 35 days. | Suppress elevated TC, TG, phospholipids and FFAs, VLDL-C, LDL-C in plasma and tissues; Suppress liver tissue inflammatory cytokines (TNF-α and IL-6); increase high density lipoproteins-cholesterol (HDL-C) | [68] |
| Carvacrol | HFD-induced type 2 diabetic C57BL/6J mice—20 mg/kg, p.o. for 35 days. | Ameliorate the increased glucose-6-phosphatase and fructose-1,6-bisphosphatase, decreased glucokinase and glucose-6-phosphate dehydrogenase activities; normalize hepatic markers (ASP, ALA, ALP, and γ-glutamyl transpeptidase). | [69] |
| Carvacrol | STZ-induced diabetic rats—25, 50, and 100 mg/kg, p.o. for 7 weeks or 20, 30 and 40 i.p. | Improve diabetes-associated cognitive deficit; suppress oxidative stress (increased MDA level and decreased SOD as well as reduced GSH) and inflammatory and apoptosis markers (NF-κB p65 unit, TNF-α, IL-1β, and caspase-3). | [70,71] |
| Carvacrol | STZ-induced diabetes in rats—25 and 50 mg/kg, p.o. for 7 days. | Suppress serum glucose, total cholesterol, ALA, AST and lactate dehydrogenase; no effect on serum insulin levels, food-water intake values and body weight changes. | [72] |
| Carvone | STZ-induced diabetic rats—50 mg/kg, p.o. for 30 days. | Reduce plasma glucose, HbA1c; improve the levels of hemoglobin and insulin. Revers activities of carbohydrate metabolic enzymes, enzymatic antioxidants and hepatic marker enzymes. | [73] |
| Catalpol | STZ-induced diabetic rats—10 mg/kg, i.p. for 14 days. | Improve impaired renal functions; ameliorate pathological changes in kidneys; abolish the diabetes induced elevation of Grb10 expression in the kidneys; increase IGF-1 mRNA levels and IGF-1R phosphorylation in kidneys. | [74] |
| Catalpol | HFD-fed mice receiving 100 mg/kg, p.o. for 4 weeks | No effect on body weight; improve fasting glucose and insulin levels, glucose tolerance and insulin tolerance; reduce macrophage infiltration into adipose tissue; reduce mRNA expressions of M1 pro-inflammatory cytokines while increasing M2 anti-inflammatory gene expressions in adipose tissue; suppress the JNK and NF-κB signaling pathways in adipose tissue. | [75] |
| Catalpol | STZ-diabetic rats—0.1 mg/kg, i.p. | Enhance glucose uptake in the isolated soleus muscle of diabetic rats; increase glycogen synthesis. | [76] |
| Catalpol | STZ-induced diabetic rats—10, 50 and 100 mg/kg, p.o. for 6 weeks. | Improve neuronal injury and cognitive dysfunction; increase the nerve growth factor concentration and decrease the blood glucose. | [77] |
| Citronellol | STZ-induced diabetic rats—25, 50, and 100 mg/kg, p.o. for 30 days. | Improve the levels of insulin, hemoglobin and hepatic glycogen with significant decrease in glucose and HbA1c levels. Restore altered activities of carbohydrate metabolic enzymes and level of hepatic and kidney markers; improve morphology of hepatic cells and insulin-positive β-cells. | [78] |
| Cymene | STZ-induced diabetic rats—20 mg/kg, p.o. for 60 days | Improve HbA1c and nephropathic parameters (like albumin excretion rate, serum creatinine and creatinine clearance rate). | [30] |
| Genipin | Aging rats—25 mg/kg, i.p. for 12 days | Ameliorate systemic and hepatic insulin resistance, alleviate hyperinsulinemia, hyperglyceridemia and hepatic steatosis, relieve hepatic oxidative stress and mitochondrial dysfunction; improve insulin sensitivity by promoting insulin-stimulated glucose consumption and glycogen synthesis; inhibit cellular ROS overproduction and alleviate the reduction of levels of MMP and ATP. | [79] |
| Geniposide | Insulin-deficient—APP/PS1 transgenic mouse model of Alzheimer’s disease. 5, 10, and 20 mg/kg, intragastric for 4 weeks. | Decrease the phosphorylation of tau protein. | [80] |
| Geniposide | STZ-induced diabetic rats—injection (50 μM, 10 μL) to the lateral ventricle | Prevent spatial learning deficit; reduce tau phosphorylation. | [81] |
| Geniposide | Transgenic mouse model with Streptozotocin—5, 10 and 20 mg/kg, intragastric for 4 weeks | Decreased level of β-amyloid peptides (Aβ1-40 and Aβ1-42); up-regulate the protein levels of β-site APP cleaving enzyme (BACE1) and insulin-degrading enzyme (IDE); decrease the protein levels of ADAM10; enhance the effects of insulin by reducing Aβ1-42 levels in primary cultured cortical neurons. | [35] |
| Geniposide | STZ-induced diabetic rats—800 mg/kg/day, p.o. for 46 days. | Improve insulin and blood glucose; decrease Aβ1-42 level; improve the expression of insulin-degrading enzyme. | [82] |
| Geniposide | Spontaneously obese Type 2 diabetic TSOD mice | Suppress body weight, visceral fat and intrahepatic lipid accumulation; alleviate abnormal lipid metabolism; alleviate abnormal glucose tolerance and hyperinsulinemia. | [40] |
| Geniposide | High fat diet—25, 50 or 100 mg/kg, p.o. for six weeks. | Improve liver histology through reducing the elevated liver index (liver weight/body weight), serum alanine aminotransferase and aspartate aminotransferase; decrease total cholesterol, triglycerides and FFAs in serum and liver; increased serum insulin levels but reduced serum TNF-α level; suppressed expression of CYP2E1 and increased PPARα expression | [83] |
| Geniposide | HFD and STZ-induced diabetic mice −200 and 400 mg/kg for 2 weeks | Decrease blood glucose, insulin and TG levels; decrease the expression of glycogen phosphorylase and glucose-6-phosphatase at mRNA level and immunoreactive protein levels, as well as enzyme activity. | [84] |
| Geraniol | STZ-induced diabetic rats—100, 200 and 400 mg/kg, p.o. for 45 days | Improve the levels of insulin, hemoglobin and decrease plasma glucose, HbA1c; improve hepatic glycogen content; preserve the normal histological appearance of hepatic cells and pancreatic β-cells. | [85] |
| d-Limonene | STZ-induced diabetic rats—50, 100 and 200 mg/kg, p.o. for 45 days. | Reverse the following diabetic effect: increased blood glucose and glycosylated hemoglobin levels, increased activity of gluconeogenic enzymes (glucose 6-phosphatase and fructose 1,6-bisphosphatase) and decreased activity of glycolytic enzyme, glucokinase and liver glycogen. | [86] |
| d-Limonene | STZ-induced diabetic rats—50 mg/kg, p.o. for 28 days. | Decrease DNA damage, glutathione reductase enzyme activities and MDA levels; increase GSH levels and CAT, SOD and GSH-Px enzyme activities and altered lipid and liver enzyme parameters in diabetic rats. | [87] |
| Logonin | STZ-induced diabetic mice, 20 mg/kg, p.o. for 12 weeks. | Reduce kidney/body weight ratio, 24 h urine protein levels, serum levels of urea nitrogen and creatinine; improve histology of pancreas and kidney; alleviate structural alterations in endothelial cells, mesangial cells and podocytes in renal cortex; reduce AGE levels in serum and kidney; downregulate mRNA and protein expression of receptors for AGEs in kidney; reduce the levels of MDA; increase the levels of SOD in serum and kidney. | [88] |
| Menthol | STZ-nicotinamide induced diabetes in rats—25, 50, and 100 mg/kg, p.o. for 45 days | Reduce blood glucose and glycosylated hemoglobin levels; increase the total hemoglobin, plasma insulin and liver glycogen levels; protect hepatic and pancreatic islets; modulating glucose metabolizing enzymes, suppression of pancreatic β-cells apoptosis and altered hepatic, pancreatic morphology | [89] |
| Myrtenal | STZ-induced diabetic rats—80 mg/kg, p.o. for 28 days | Decrease plasma glucose; increase plasma insulin levels; up-regulate IRS2, Akt and GLUT2 in liver; increase IRS2, Akt and GLUT4 protein expression in skeletal muscle. | [90] |
| Myrtenal | STZ-induced diabetic rats—20, 40, and 80 mg/kg, p.o. for 28 days | Reduce plasma glucose, haemoglobin A1c (HbA1c); increase the levels of insulin and hemoglobin; reverse body weight loss; normalize hexokinase, glucose-6-phosphatase, fructose-1,6-bisphosphatase, glucose-6-phosphate dehydrogenase, and hepatic enzymes AST, ALT, and ALP levels; improve hepatic and muscle glycogen content; restore islet cells and liver histology. | [91] |
| Myrtenal | STZ-induced diabetic rats—80 mg/kg, p.o. for 28 days | Improve plasma glucose, pancreatic insulin and lipid profiles (TC, TG, FFAs, phospholipids, LDL, VLDL, atherogenic index); improve histopathological feature of the liver. | [92] |
| Paeoniflorin | High-sucrose, HFD rat receiving low dose STZ—15 and 30 mg/kg, p.o. for 4 weeks. | Reduce brain inflammatory cytokines (IL-1β and TNF-α), decrease suppressor of cytokine signaling 2 expressions and promote (IRS-1 activity and phosphorylation levels of protein kinase B (Akt) and glycogen synthase kinase-3β (GSK-3β). | [93] |
| Paeoniflorin | HFD—induced obese mice—Diet containing 0.05% (w/w) of paeoniflorin | Lower body weight, hyperlipidemia, and insulin resistance; block inflammation; inhibiting lipid ectopic deposition; lower lipid synthesis pathway (de novo pathway, 3HMG-CoAR), promote fatty acid oxidation (peroxisome proliferator-activated receptor-alpha (PPARα), carnitine palmitoyltransferase-1); increase cholesterol output (PPARγ-liver X receptor-α-ATP-binding cassette transporter-1); block inflammatory genes activation and reduce gluconeogenic genes expression (phosphoenolpyruvate carboxykinase and G6Pase). | [82] |
| Swertiamarin | STZ-induced diabetic rats—50 mg/kg, i.p. for 6 weeks | Reduce serum triglycerides, cholesterol and low-density lipoprotein levels; decrease serum fasting glucose; increase insulin sensitivity index. | [94] |
| Thymol | HFD-induced type 2 diabetes in C57BL/6J mice—10, 20 and 40 mg/kg, intragastric for 5 weeks. | Antihyperglycaemic; lower plasma TG, TC, FFAs, LDL and increase HDL cholesterol; lower hepatic lipid contents TG, total cholesterol, FFAs and phospholipids. | [95] |
| Thymol | HFD- induced type 2 diabetes in C57BL/6J mice—40 mg/kg, intragastric for 5 weeks. | Inhibit diabetic nephropathy; inhibit the activation of transforming growth factor-β1 and vascular endothelial growth factor; increase level of antioxidants and suppress lipid peroxidation markers in erythrocytes and kidney tissues; downregulate the expression level of sterol regulatory element binding protein-1c and reduce lipid accumulation in the kidney. | [96] |
| Thymol | HFD-induced obese C57BL/6 J mice—20, 40 mg/kg daily | Reverse body weight gain; ameliorate peripheral insulin resistance; improve cognitive impairments in the Morris Water Maze test; decrease HFD-induced Aβ deposition and tau hyperphosphorylation in the hippocampus; down-regulate the level of P-Ser307 IRS-1 and enhance the expression of P-Ser473 AKT and P-Ser9 GSK3β; up-regulate nuclear respiratory factor /heme oxygenase-1pathway. | [97] |
| Thymol | HFD-fed rats—14 mg/kg, p.o. for 4 weeks. | Decrease body weight gain, visceral fat -pad weights, lipids, ALT, AST, LDH, blood urea nitrogen, glucose, insulin, and leptin levels; decrease serum lipid peroxidation and increase antioxidant levels. | [98] |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habtemariam, S. Antidiabetic Potential of Monoterpenes: A Case of Small Molecules Punching above Their Weight. Int. J. Mol. Sci. 2018, 19, 4. https://doi.org/10.3390/ijms19010004
Habtemariam S. Antidiabetic Potential of Monoterpenes: A Case of Small Molecules Punching above Their Weight. International Journal of Molecular Sciences. 2018; 19(1):4. https://doi.org/10.3390/ijms19010004
Chicago/Turabian StyleHabtemariam, Solomon. 2018. "Antidiabetic Potential of Monoterpenes: A Case of Small Molecules Punching above Their Weight" International Journal of Molecular Sciences 19, no. 1: 4. https://doi.org/10.3390/ijms19010004
APA StyleHabtemariam, S. (2018). Antidiabetic Potential of Monoterpenes: A Case of Small Molecules Punching above Their Weight. International Journal of Molecular Sciences, 19(1), 4. https://doi.org/10.3390/ijms19010004