Advances in Understanding the Mechanism of Action of the Auxin Permease AUX1



Abstract
:
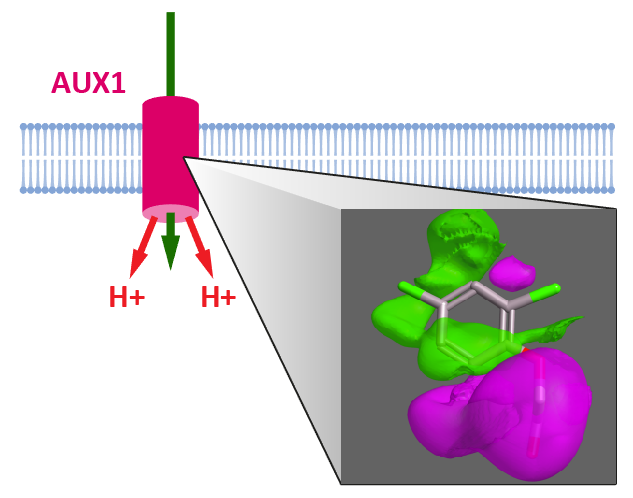{kind=link}
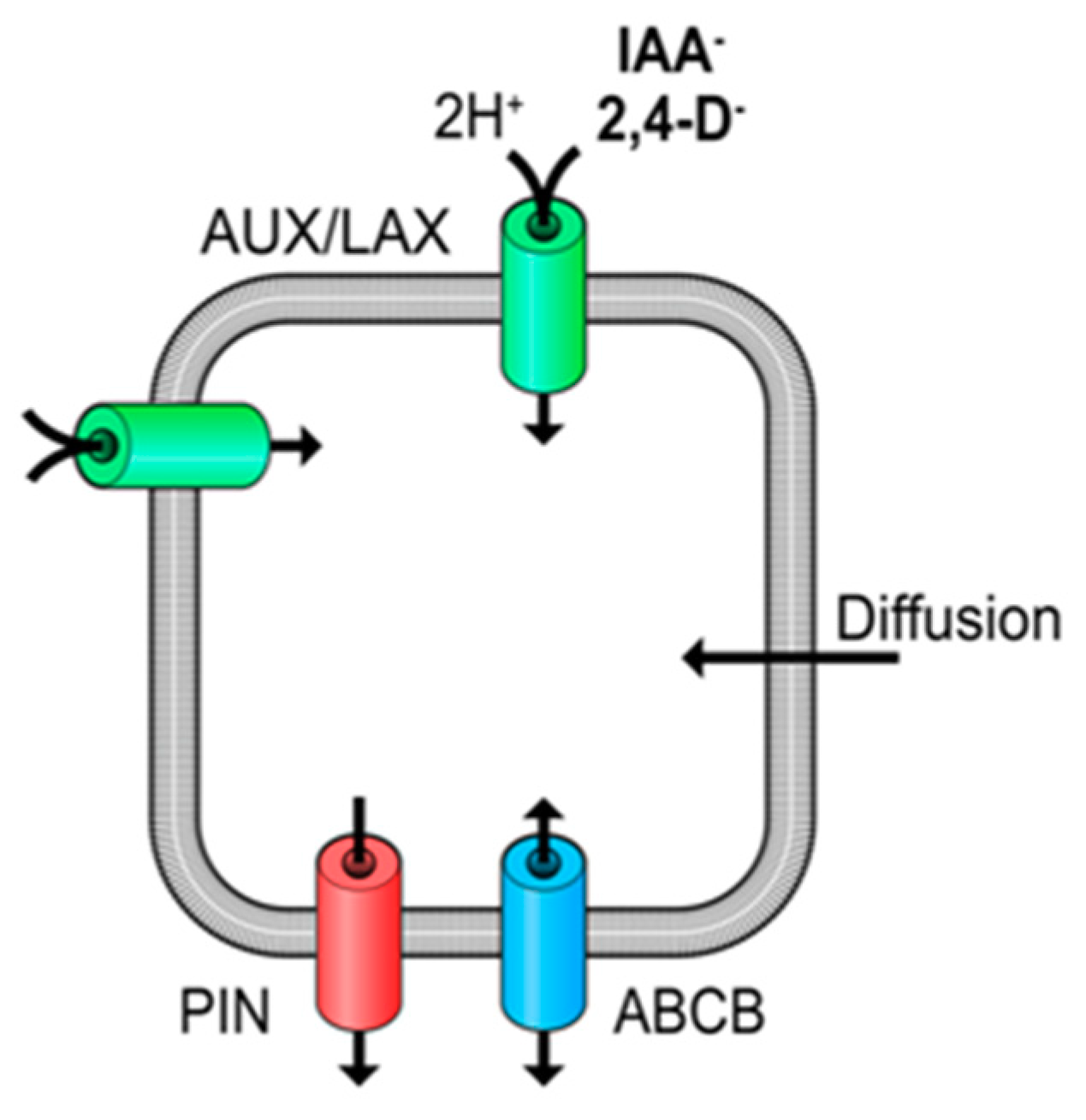{kind=link}
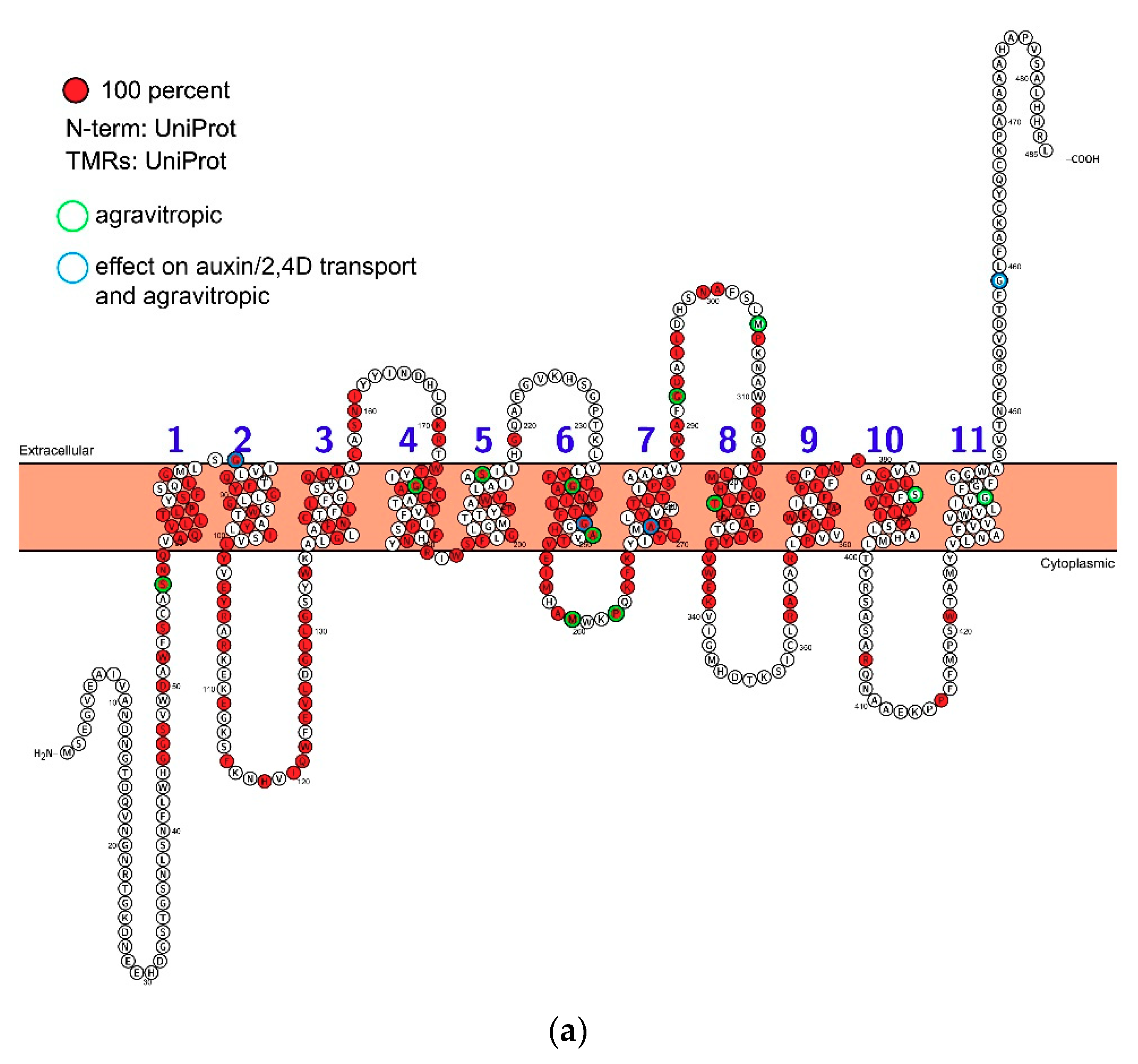{kind=link}
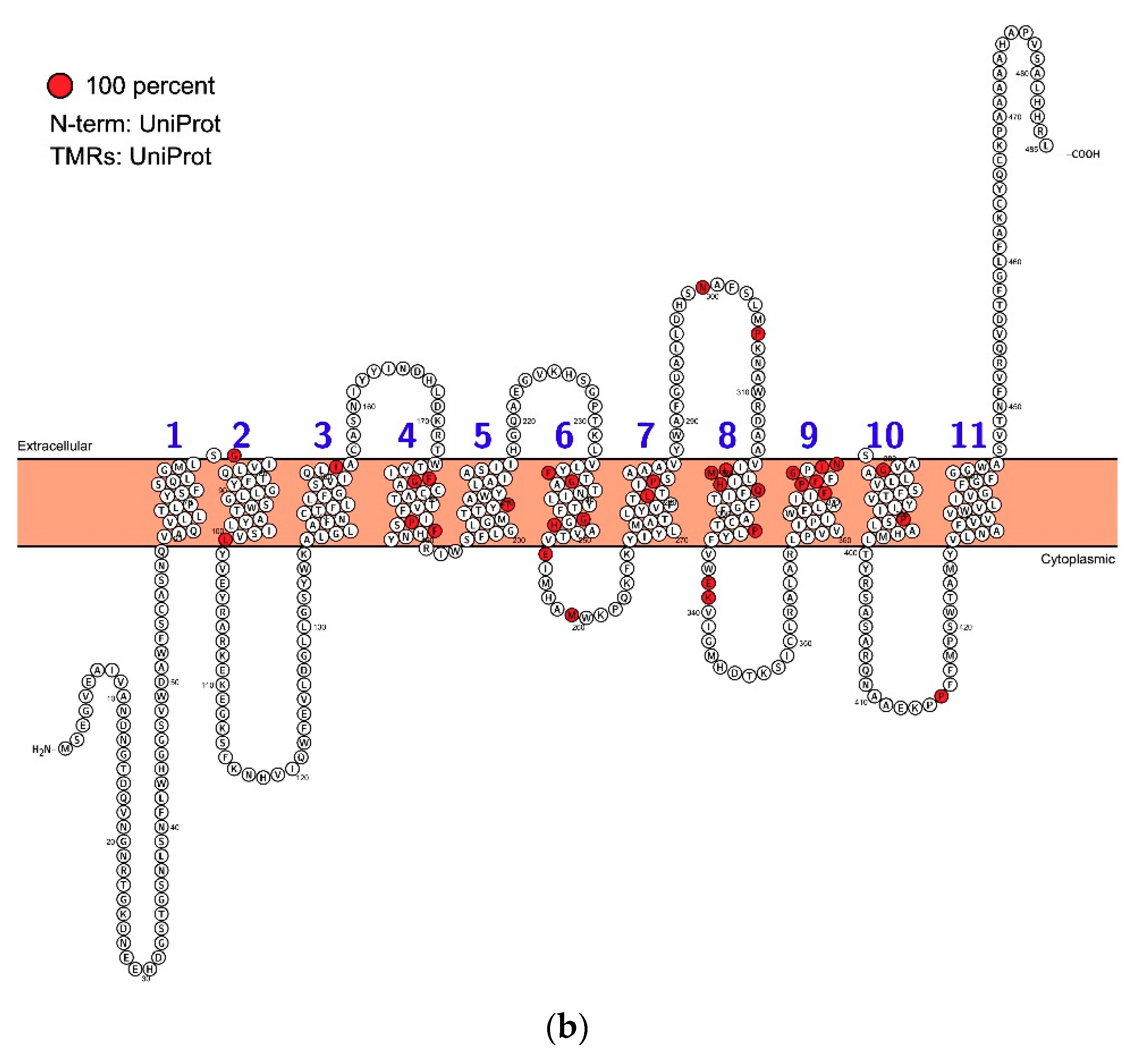{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Early Discoveries and the Chemiosmotic Theory for Auxin Accumulation
3. Auxin Uptake Carriers Are Proton Co-Transporters
4. AUX1 and LAX Proteins Identified as Auxin Transporters
5. Selectivity of the Auxin Uptake System
6. Uptake Selectivity and Auxin Herbicide Activity
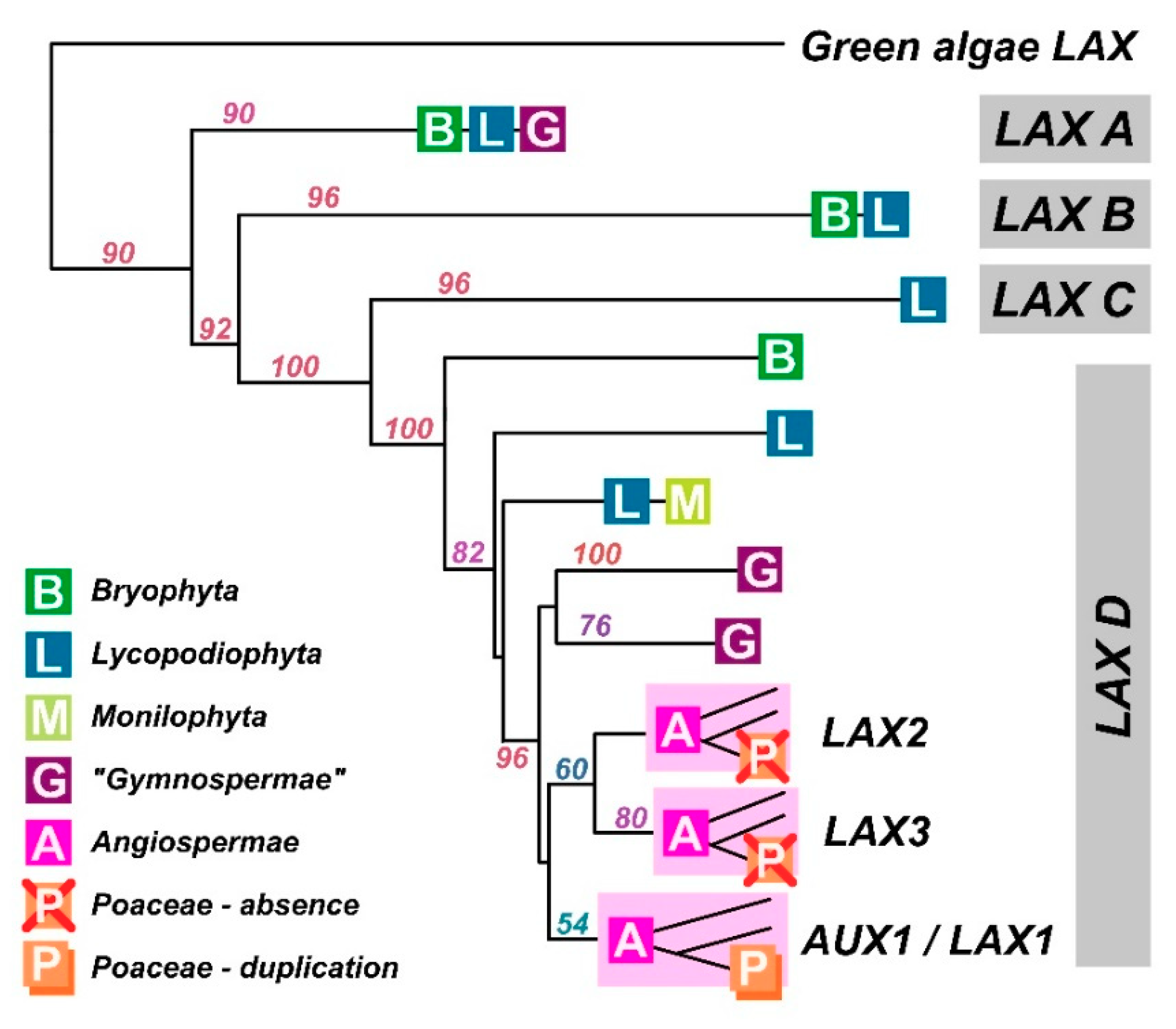
7. Evolutionary Analysis of AUX1
8. Sequence and Structure
9. AUX1 kinetics and Models of Auxin Action
10. Summary and Gaps in Our Knowledge
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Lacek, J.; Retzer, K.; Luschnig, C.; Zažímalová, E. Polar Auxin Transport. In ELS; John Wiley & Sons Ltd.: Chichester, UK, 2017; Available online: http://www.els.net (accessed on 17 April 2017).
- Swarup, R.; Péret, B. AUX/LAX family of auxin influx carriers-an overview. Front Plant Sci. 2012, 3, 225. [Google Scholar] [CrossRef] [PubMed]
- Terasaka, K.; Blakeslee, J.J.; Titapiwatanakun, B.; Peer, W.A.; Bandyopadhyay, A.; Makam, S.N.; Lee, O.R.; Richards, E.L.; Murphy, A.S.; Sato, F.; et al. PGP4, an ATP binding cassette P-glycoprotein, catalyzes auxin transport in Arabidopsis thaliana roots. Plant Cell 2005, 17, 2922–2939. [Google Scholar] [CrossRef] [PubMed]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; jung, K.; et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Kamimoto, Y.; Terasaka, K.; Hamamoto, M.; Takanashi, K.; Fukuda, S.; Shitan, N.; Sugiyama, A.; Suzuki, H.; Shibata, D.; Wang, B.; et al. Arabidopsis ABCB21 is a facultative auxin importer/exporter regulated by cytoplasmic auxin concentration. Plant Cell Physiol. 2012, 53, 2090–2100. [Google Scholar] [CrossRef] [PubMed]
- Kubeš, M.; Yang, H.; Richter, G.L.; Cheng, Y.; Młodzińska, E.; Wang, X.; Blakeslee, J.J.; Carraro, N.; Petrášek, J.; Zažímalová, E.; et al. The Arabidopsis concentration-dependent influx/efflux transporter ABCB4 regulates cellular auxin levels in the root epidermis. Plant J. 2012, 69, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Swarup, R.; Friml, J.; Marchant, A.; Ljung, K.; Sandberg, G.; Palme, K.; Bennett, M. Localization of the auxin permease AUX1 suggests two functionally distinct hormone transport pathways operate in the Arabidopsis root apex. Genes Dev. 2001, 15, 2648–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchant, A.; Kargul, J.; May, S.T.; Muller, P.; Delbarre, A.; Perrot-Rechenmann, C.; Bennett, M.J. AUX1 regulates root gravitropism in Arabidopsis by facilitating auxin uptake within root apical tissues. EMBO J. 1999, 18, 2066–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbussche, F.; Petrásek, J.; Zádníková, P.; Hoyerová, K.; Pesek, B.; Raz, V.; Bennett, M.; Zažímalová, E.; Benková, E.; Van Der Straeten, D. The auxin influx carriers AUX1 and LAX3 are involved in auxin-ethylene interactions during apical hook development in Arabidopsis thaliana seedlings. Development 2010, 137, 597–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarup, K.; Benková, E.; Swarup, R.; Casimiro, I.; Péret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S.; et al. The auxin influx carrier LAX3 promotes lateral root emergence. Nat. Cell Biol. 2008, 10, 946–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasprzewska, A.; Carter, R.; Swarup, R.; Bennett, M.; Monk, N.; Hobbs, J.K.; Fleming, A. Auxin influx importers modulate serration along the leaf margin. Plant J. 2015, 83, 705–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhosale, R.; Giri, J.; Pandey, B.K.; Giehl, R.F.H.; Hartmann, A.; Traini, R.; Truskina, J.; Leftley, N.; Hanlon, M.; Swarup, K.; et al. A mechanistic framework for auxin dependent Arabidopsis root hair elongation to low external phosphate. Nat Commun. 2018, 9, 1409. [Google Scholar] [CrossRef] [PubMed]
- Dindas, J.; Scherzer, S.; Roelfsema, M.R.G.; von Meyer, K.; Müller, H.M.; Al-Rasheid, K.A.S.; Palme, K.; Dietrich, P.; Becker, D.; Bennett, M.J.; et al. AUX1-mediated root hair auxin influx governs SCF. Nat. Commun. 2018, 9, 1174. [Google Scholar] [CrossRef] [PubMed]
- Fendrych, M.; Akhmanova, M.; Merrin, J.; Glanc, M.; Hagihara, S.; Takahashi, K.; Uchida, N.; Torii, K.U.; Friml, J. Rapid and reversible root growth inhibition by TIR1 auxin signalling. Nat. Plants 2018, 4, 453–459. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, B.M.; Neale, H.C.; Geilfus, C.M.; Jackson, R.W.; Arnold, D.L.; Preston, G.M. Early changes in apoplast composition associated with defence and disease in interactions between Phaseolus vulgaris and the halo blight pathogen Pseudomonas syringae Pv. phaseolicola. Plant Cell Environ. 2016, 39, 2172–2184. [Google Scholar] [CrossRef] [PubMed]
- Rubery, P.H.; Sheldrake, A.R. Carrier-mediated auxin transport. Planta 1974, 118, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Parry, G.; Delbarre, A.; Marchant, A.; Swarup, R.; Napier, R.; Perrot-Rechenmann, C.; Bennett, M.J. Novel auxin transport inhibitors phenocopy the auxin influx carrier mutation aux1. Plant J. 2001, 25, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubery, P.H.; Sheldrake, A.R. Effect of pH and surface charge on cell uptake of auxin. Nat. New Biol. 1973, 244, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.; Hertel, R. Auxin binding to subcellular fractions from Cucurbita hypocotyls: In vitro evidence for an auxin transport carrier. Planta 1978, 142, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hertel, R.; Lomax, T.L.; Briggs, W.R. Auxin transport in membrane vesicles from Cucurbita pepo L. Planta 1983, 157, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Lomax, T.L.; Mehlhorn, R.J.; Briggs, W.R. Active auxin uptake by zucchini membrane vesicles: Quantitation using ESR volume and delta pH determinations. Proc. Natl. Acad. Sci. USA 1985, 82, 6541–6545. [Google Scholar] [CrossRef] [PubMed]
- Benning, C. Evidence supporting a model of voltage-dependent uptake of auxin into Cucurbita vesicles. Planta 1986, 169, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Geier, U.; Werner, O.; Bopp, M. Indole-3-acetic acid uptake in isolated protoplasts of the moss Funaria hygrometrica. Planta 1990, 80, 584–592. [Google Scholar] [CrossRef]
- Barbez, E.; Dünser, K.; Gaidora, A.; Lendl, T.; Busch, W. Auxin steers root cell expansion via apoplastic pH regulation in. Proc. Natl. Acad. Sci. USA 2017, 114, E4884–E4893. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.I.; Takahashi, K.; Okumura-Noda, H.; Kinoshita, T. Auxin Influx Carrier AUX1 Confers Acid Resistance for Arabidopsis Root Elongation Through the Regulation of Plasma Membrane, H+-ATPase. Plant Cell Physiol. 2016, 57, 2194–2201. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.J.; Marchant, A.; Green, H.G.; May, S.T.; Ward, S.P.; Millner, P.A.; Walker, A.R.; Schulz, B.; Feldmann, K.A. Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 1996, 273, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Maher, E.P.; Martindale, S.J. Mutants of Arabidopsis thaliana with altered responses to auxins and gravity. Biochem. Genet. 1980, 18, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Shimura, Y. Reversible root tip rotation in Arabidopsis seedlings induced by obstacle-touching stimulus. Science 1990, 250, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Swarup, R.; Kargul, J.; Marchant, A.; Zadik, D.; Rahman, A.; Mills, R.; Yemm, A.; May, S.; Williams, L.; Millner, P.; et al. Structure-function analysis of the presumptive Arabidopsis auxin permease AUX1. Plant Cell 2004, 16, 3069–3083. [Google Scholar] [CrossRef] [PubMed]
- Péret, B.; Swarup, K.; Ferguson, A.; Seth, M.; Yang, Y.; Dhondt, S.; James, N.; Casimiro, I.; Perry, P.; Syed, A.; et al. AUX/LAX genes encode a family of auxin influx transporters that perform distinct functions during Arabidopsis development. Plant Cell 2012, 24, 2874–2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarup, R.; Kramer, E.M.; Perry, P.; Knox, K.; Leyser, H.M.; Haseloff, J.; Beemster, G.T.S.; Bhalerao, R.; Bennett, M.J. Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal. Nat. Cell Biol. 2005, 7, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Kramer, E.M.; Bennett, M.J. Auxin transport: A field in flux. Trends Plant Sci. 2006, 11, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hammes, U.Z.; Taylor, C.G.; Schachtman, D.P.; Nielsen, E. High-affinity auxin transport by the AUX1 influx carrier protein. Curr. Biol. 2006, 16, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Carrier, D.J.; Bakar, N.T.; Swarup, R.; Callaghan, R.; Napier, R.M.; Bennett, M.J.; Kerr, I.D. The binding of auxin to the Arabidopsis auxin influx transporter AUX1. Plant Physiol. 2008, 148, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Lanková, M.; Smith, R.S.; Pesek, B.; Kubes, M.; Zazímalová, E.; Petrásek, J.; Hoyerová, K. Auxin influx inhibitors 1-NOA, 2-NOA, and CHPAA interfere with membrane dynamics in tobacco cells. J. Exp. Bot. 2010, 61, 3589–3598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delbarre, A.; Muller, P.; Imhoff, V.; Guern, J. Comparison of mechanisms controlling uptake and accumulation of 2,4-dichlorophenoxy acetic acid, naphthalene-1-acetic acid, and indole-3-acetic acid in suspension-cultured tobacco cells. Planta 1996, 198, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, V.; Muller, P.; Guern, J.; Delbarre, A. Inhibitors of the carrier-mediated influx of auxin in suspension-cultured tobacco cells. Planta 2000, 210, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Hošek, P.; Kubes, M.; Lanková, M.; Dobrev, P.I.; Klíma, P.; Kohoutová, M.; Petrášek, J.; Hoyerová, K.; Jiřina, M.; Zažímalová, E. Auxin transport at cellular level: New insights supported by mathematical modelling. J. Exp. Bot. 2012, 63, 3815–3827. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Kubeš, M.; Baster, P.; Robert, S.; Dobrev, P.I.; Friml, J.; Petrášek, J.; Zažímalová, E. Defining the selectivity of processes along the auxin response chain: A study using auxin analogues. New Phytol. 2013, 200, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Sussman, M.R.; Goldsmith, M.H. Auxin uptake and action of N-1-naphthylphthalamic acid in corn coleoptiles. Planta 1981, 151, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, E.; Yang, H.; Nishimura, T.; Uehara, Y.; Sakai, T.; Furutani, M.; Koshiba, T.; Hirose, M.; Nozaki, H.; Murphy, A.S.; et al. Alkoxy-auxins are selective inhibitors of auxin transport mediated by PIN, ABCB, and AUX1 transporters. J. Biol. Chem. 2011, 286, 2354–2364. [Google Scholar] [CrossRef] [PubMed]
- Hoyerova, K.; Hosek, P.; Quareshy, M.; Li, J.; Klima, P.; Kubes, M.; Yemm, A.A.; Neve, P.; Tripathi, A.; Bennett, M.J.; et al. Auxin molecular field maps define AUX1 selectivity: Many auxin herbicides are not substrates. New Phytol. 2018, 217, 1625–1639. [Google Scholar] [CrossRef] [PubMed]
- Quareshy, M.; Prusinska, J.; Kieffer, M.; Fukui, K.; Pardal, A.J.; Lehmann, S.; Schafer, P.; del Genio, C.I.; Kepinski, S.; Hayashi, K.; et al. The Tetrazole Analogue of the Auxin Indole-3-acetic Acid Binds Preferentially to TIR1 and Not AFB5. ACS Chem. Biol. 2018, 13, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B. Implications of endogenous roles of transporters for drug discovery: Hitchhiking and metabolite-likeness. Nat. Rev. Drug Discov. 2016, 15, 143. [Google Scholar] [CrossRef] [PubMed]
- Busi, R.; Goggin, D.E.; Heap, I.; Horak, M.J.; Jugulam, M.; Masters, R.A.; Napier, R.M.; Riar, D.S.; Satchivi, N.M.; Torra, J.; et al. Weed resistance to synthetic auxin herbicides. Pest Manag. Sci. 2018, 74, 2265–2276. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T.; Brockington, S.F.; Rothfels, C.; Graham, S.W.; Stevenson, D.; Kutchan, T.; Rolf, M.; Thomas, P.; Wong, G.K.-S.; Leyser, O.; et al. Paralogous radiations of PIN proteins with multiple origins of noncanonical PIN structure. Mol. Biol. Evol. 2014, 31, 2042–2060. [Google Scholar] [CrossRef] [PubMed]
- De Smet, I.; Voss, U.; Lau, S.; Wilson, M.; Shao, N.; Timme, R.E.; Swarup, R.; Kerr, I.; Hodgman, C.; Bock, R.; et al. Unraveling the evolution of auxin signaling. Plant Physiol. 2011, 155, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.N.; Kwart, M.; Hummel, S.; Frommer, W.B. Substrate specificity and expression profile of amino acid transporters (AAPs) in Arabidopsis. J. Biol. Chem. 1995, 270, 16315–16320. [Google Scholar] [CrossRef] [PubMed]
- Fowler, P.W.; Orwick-Rydmark, M.; Radestock, S.; Solcan, N.; Dijkman, P.M.; Lyons, J.A.; Kwok, J.; Caffrey, M.; Watts, A.; Forrest, L.R.; et al. Gating topology of the proton-coupled oligopeptide symporters. Structure 2015, 23, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Jardetzky, O. Protein dynamics and conformational transitions in allosteric proteins. Prog. Biophys. Mol. Biol. 1996, 65, 171–219. [Google Scholar] [CrossRef]
- Kramer, E.M.; Rutschow, H.L.; Mabie, S.S. AuxV: A database of auxin transport velocities. Trends Plant Sci. 2011, 16, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Naramoto, S. Polar transport in plants mediated by membrane transporters: Focus on mechanisms of polar auxin transport. Curr. Opin. Plant Biol. 2017, 40, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Band, L.R.; Wells, D.M.; Fozard, J.A.; Ghetiu, T.; French, A.P.; Pound, M.P.; Wilson, M.H.; Yu, L.; Li, W.; Hijazi, H.I.; et al. Systems analysis of auxin transport in the Arabidopsis root apex. Plant Cell 2014, 26, 862–875. [Google Scholar] [CrossRef] [PubMed]
- Band, L.R.; Wells, D.M.; Larrieu, A.; Sun, J.; Middleton, A.M.; French, A.P.; Brunoud, G.; Sato, E.M.; Wilson, M.H.; Péret, B.; et al. Root gravitropism is regulated by a transient lateral auxin gradient controlled by a tipping-point mechanism. Proc. Natl. Acad. Sci. USA 2012, 109, 4668–4673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mironova, V.V.; Omelyanchuk, N.A.; Novoselova, E.S.; Doroshkov, A.V.; Kazantsev, F.V.; Kochetov, A.V.; Kolchanov, N.A.; Mjolsness, E.; Likhoshvai, V.A. Combined in silico/in vivo analysis of mechanisms providing for root apical meristem self-organization and maintenance. Ann. Bot. 2012, 110, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieneisen, V.A.; Xu, J.; Marée, A.F.; Hogeweg, P.; Scheres, B. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature 2007, 449, 1008–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wabnik, K.; Kleine-Vehn, J.; Balla, J.; Sauer, M.; Naramoto, S.; Reinöhl, V.; Merks, R.M.; Govaerts, W.; Friml, J. Emergence of tissue polarization from synergy of intracellular and extracellular auxin signaling. Mol. Syst. Biol. 2010, 6, 447. [Google Scholar] [CrossRef] [PubMed]
- Rutschow, H.L.; Baskin, T.I.; Kramer, E.M. The carrier AUXIN RESISTANT (AUX1) dominates auxin flux into Arabidopsis protoplasts. New Phytol. 2014, 204, 536–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, S.; Liu, J.; Zhang, X.; Lindsey, K. A recovery principle provides insight into auxin pattern control in the Arabidopsis root. Sci. Rep. 2017, 7, 43004. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, G.; Retzer, K.; Vosolsobě, S.; Napier, R. Advances in Understanding the Mechanism of Action of the Auxin Permease AUX1. Int. J. Mol. Sci. 2018, 19, 3391. https://doi.org/10.3390/ijms19113391
Singh G, Retzer K, Vosolsobě S, Napier R. Advances in Understanding the Mechanism of Action of the Auxin Permease AUX1. International Journal of Molecular Sciences. 2018; 19(11):3391. https://doi.org/10.3390/ijms19113391
Chicago/Turabian StyleSingh, Gaurav, Katarzyna Retzer, Stanislav Vosolsobě, and Richard Napier. 2018. "Advances in Understanding the Mechanism of Action of the Auxin Permease AUX1" International Journal of Molecular Sciences 19, no. 11: 3391. https://doi.org/10.3390/ijms19113391
APA StyleSingh, G., Retzer, K., Vosolsobě, S., & Napier, R. (2018). Advances in Understanding the Mechanism of Action of the Auxin Permease AUX1. International Journal of Molecular Sciences, 19(11), 3391. https://doi.org/10.3390/ijms19113391