Small Molecules Enhance Scaffold-Based Bone Grafts via Purinergic Receptor Signaling in Stem Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
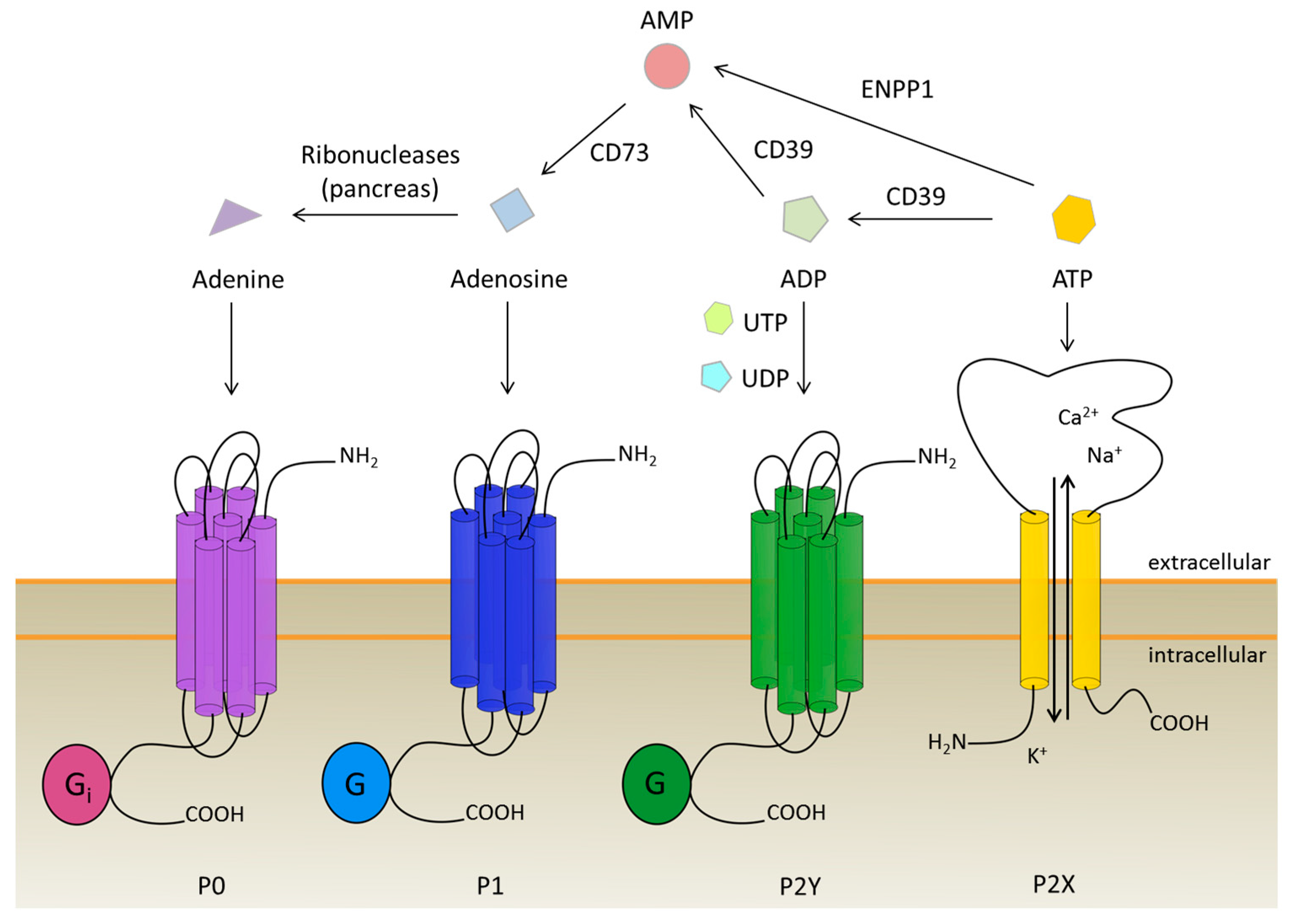
2. The Purinergic Receptor Family: Four Types Triggered by One Molecule and Its Derivatives
2.1. The New Kid on the Block: P0 Receptors
2.2. Powered by Adenosine: P1 Receptors
2.3. The Ion Channels: P2X Receptors
2.4. The Biggest Group: P2Y Receptors
3. The Role of Purinergic Receptors during Osteogenesis and Angiogenesis
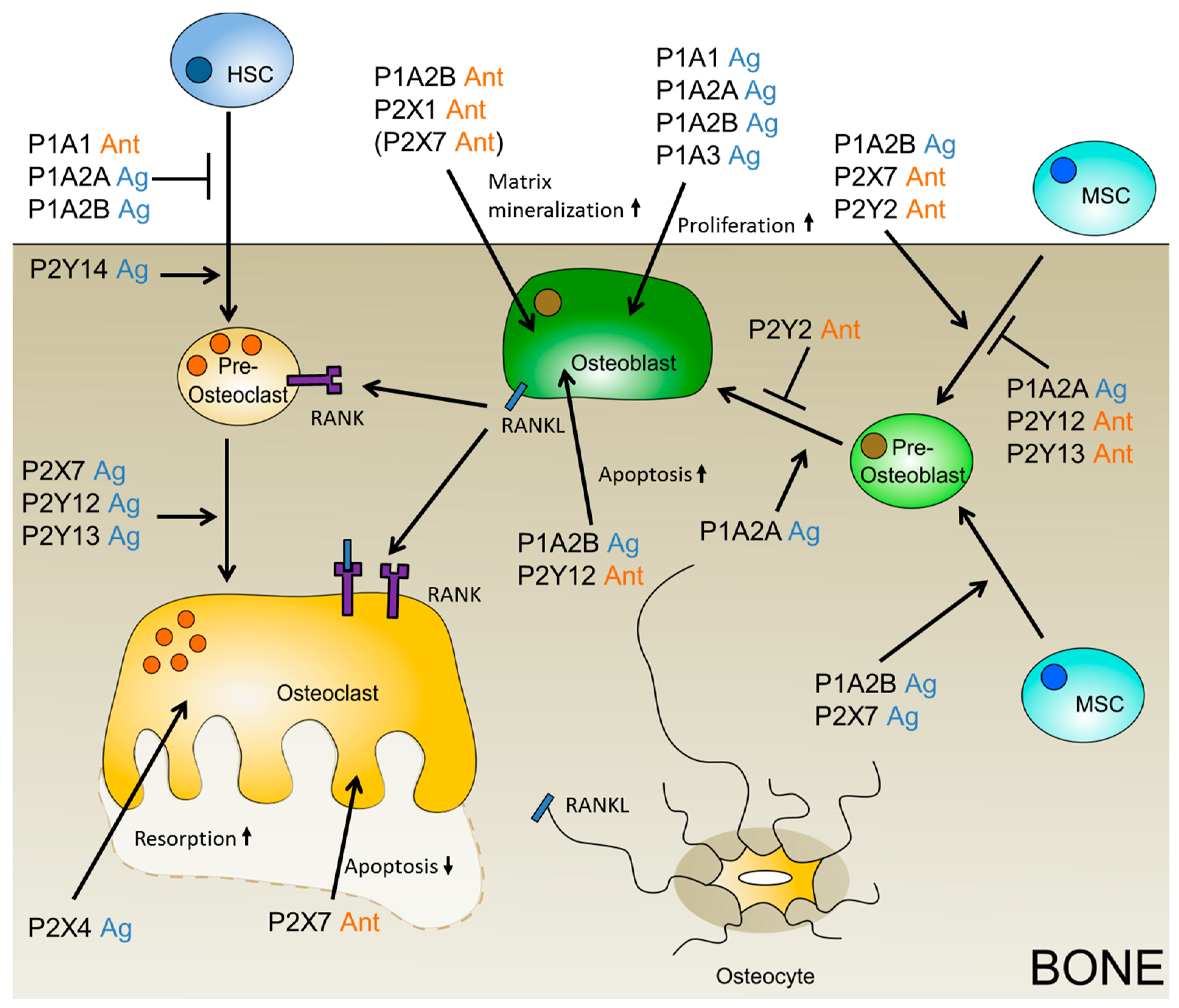
3.1. Insight into Purinergic Signaling during Osteogenesis
3.2. Insight into Purinergic Signaling During Osteoclastogenesis
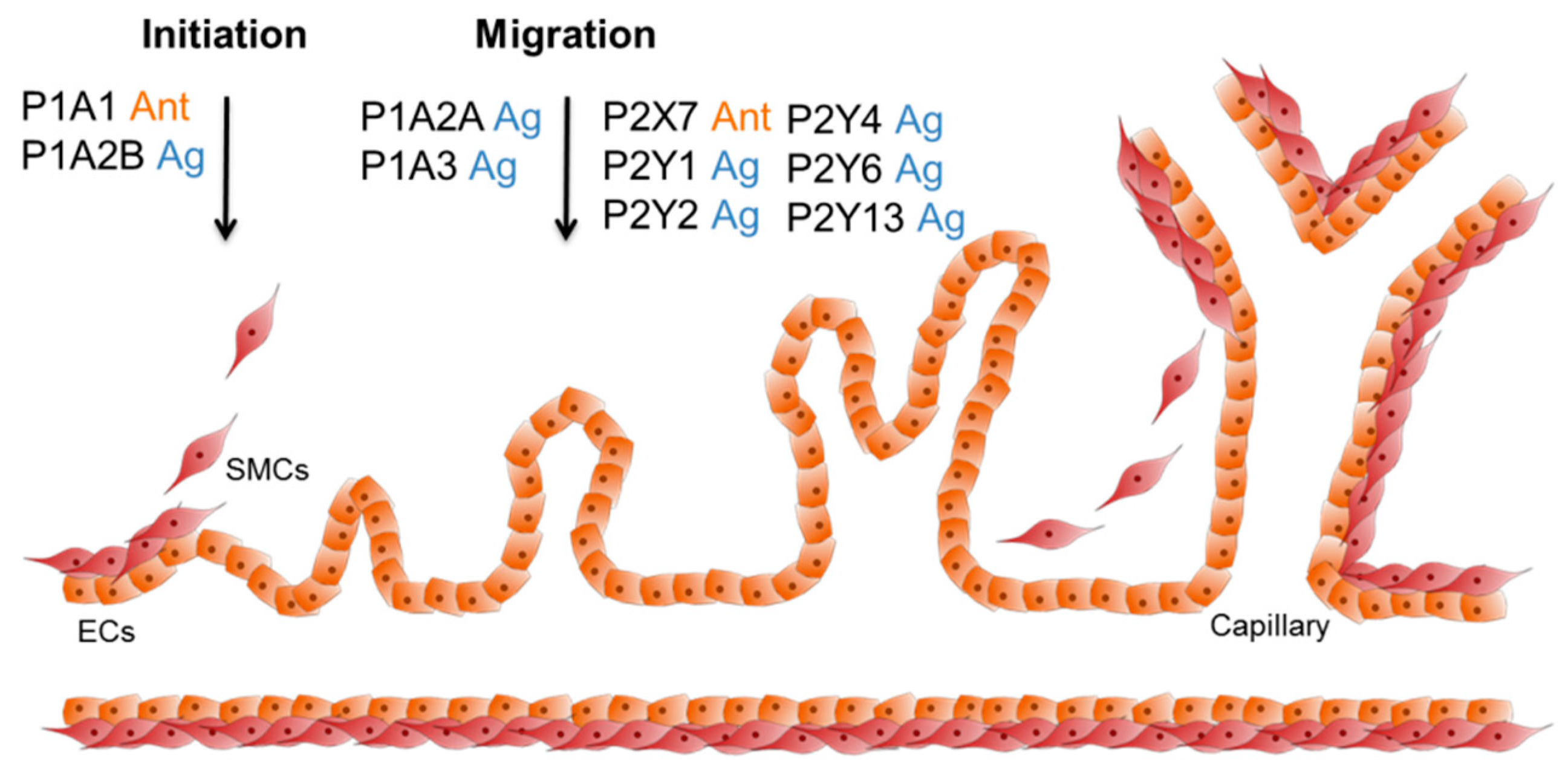
3.3. The Role of Purinergic Receptors during Angiogenesis with Respect to Artificial Bone Grafting
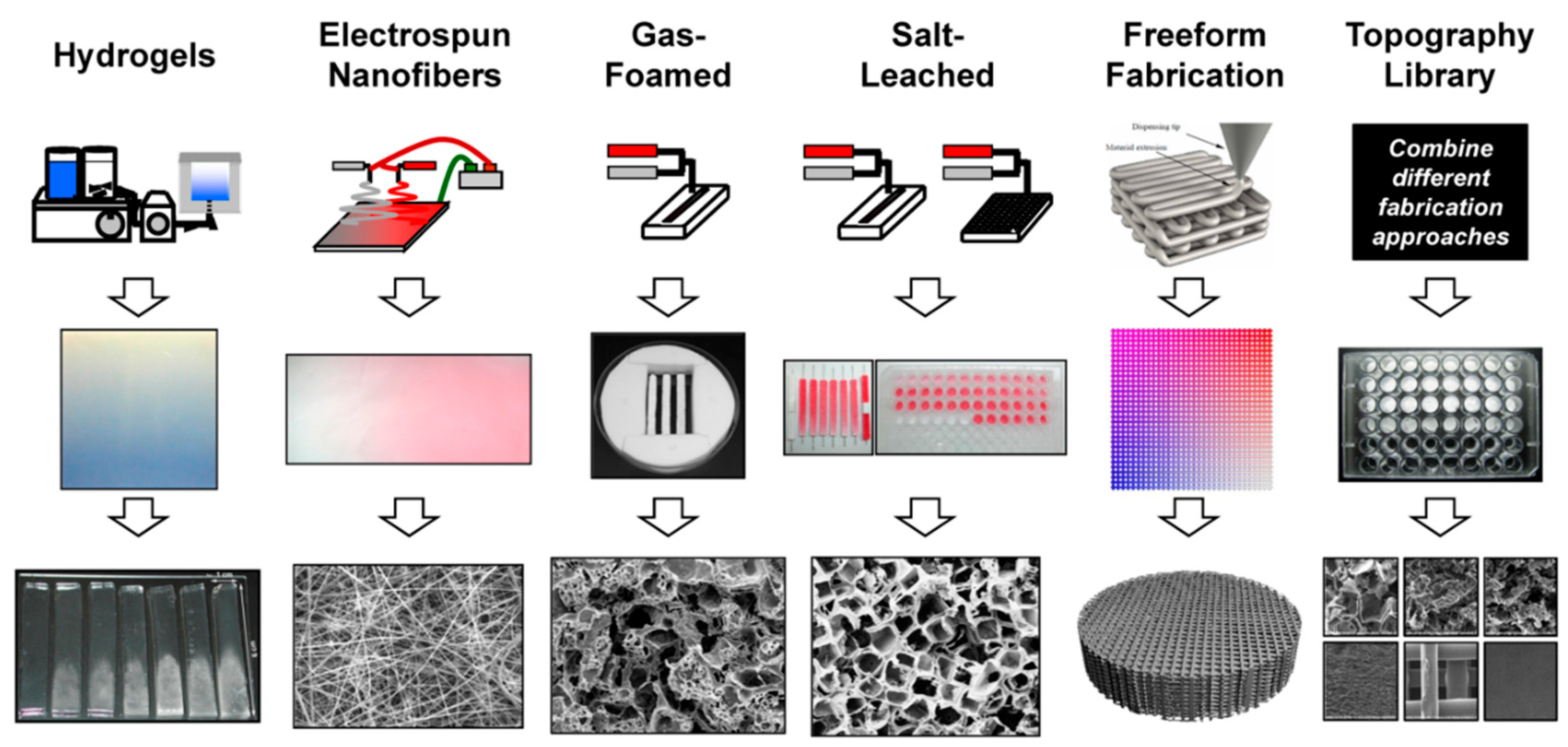
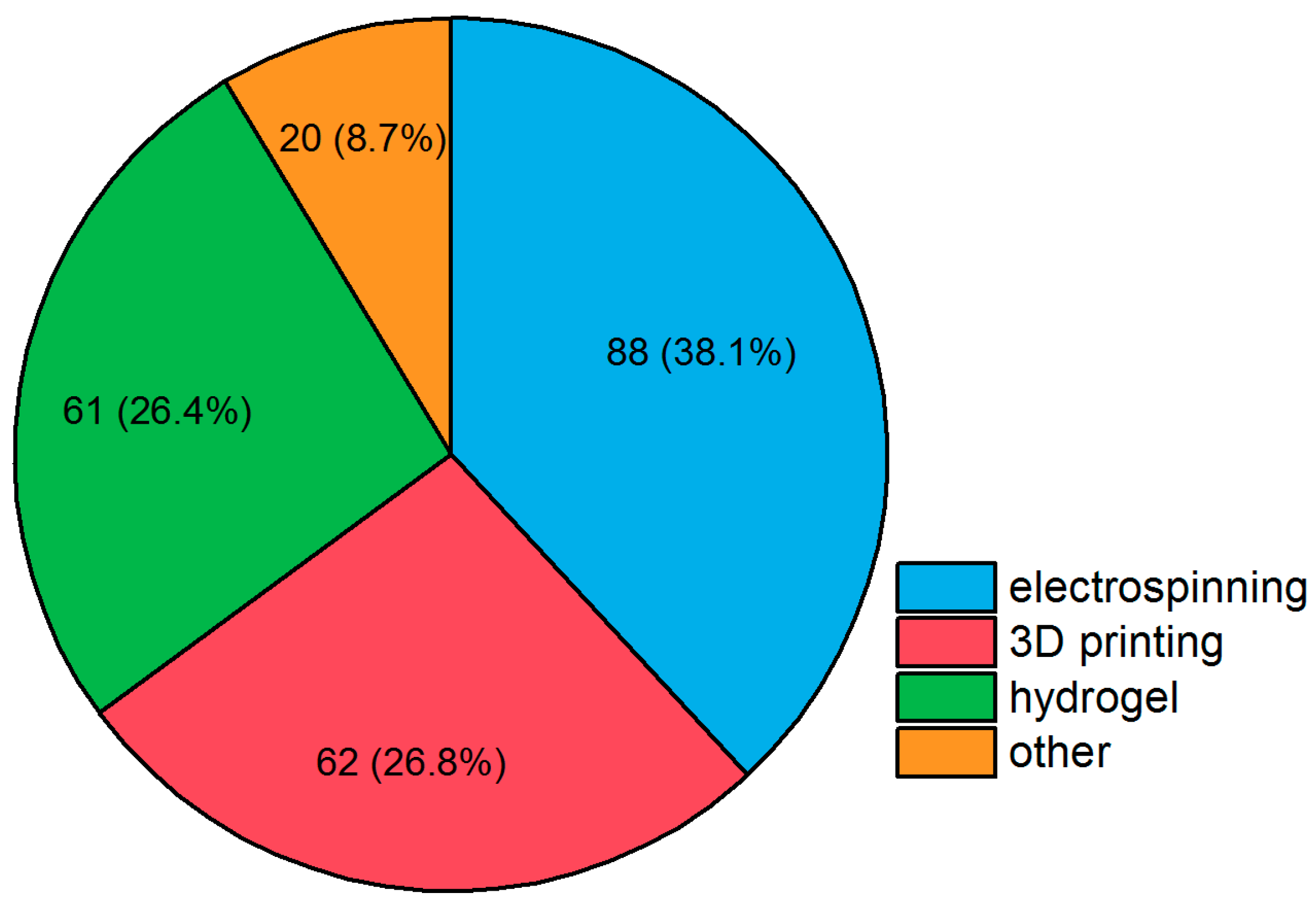
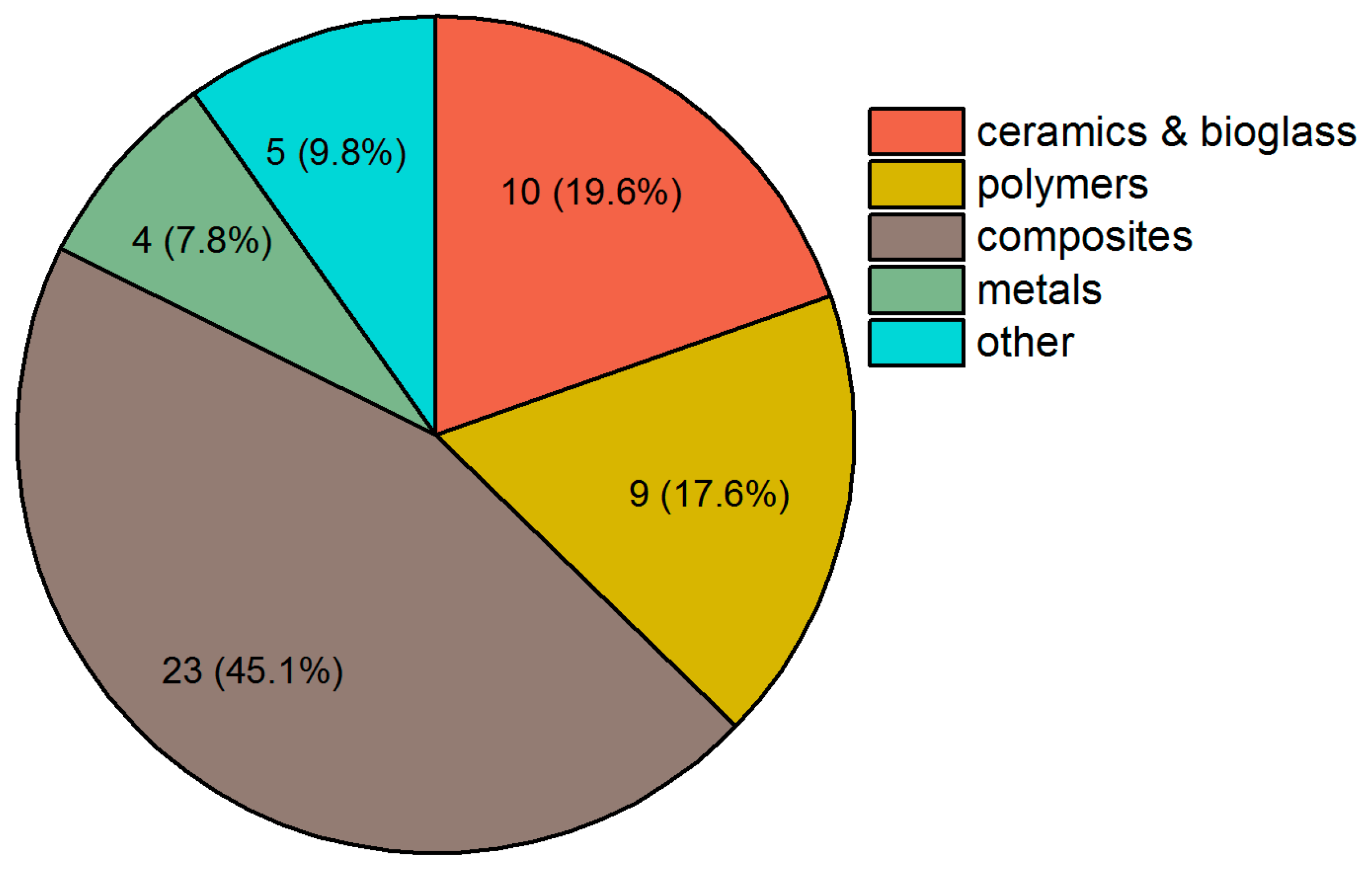
4. Strategies in Scaffold Manufacturing
4.1. 3D Printing
4.2. Electrospinning
4.3. Hydrogels
4.4. Other Approaches
5. Recent Patents and Clinical Trials in Bone Grafting via Scaffold Manufacturing
6. Recent Patents on Purinergic Receptors That May Enhance Bone Grafting Strategies
6.1. Patents on Ectonucleotidases and P1 Receptors
6.2. Patents on P2 Receptors
7. Future Aspects of Purinergic Receptors and Their Ligands in Bone Grafting
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALP | alkaline phosphatase |
| CMS | centrifugal melt spinning |
| DPSC | dental pulp stem cell |
| EC | endothelial cell |
| EPC | endothelial progenitor cell |
| HA | hydroxyapatite |
| HSC | hematopoietic stem cell |
| HUVECs | human umbilical vein endothelial cells |
| ICBG | iliac crest bone graft |
| MCSF-1 | macrophage colony-stimulating factor-1 |
| (MBG)-βTCP | mesoporous bioactive glass-β-tricalcium phosphate |
| MSC | mesenchymal stem cell |
| nGO | nanographene oxide |
| PBS | phosphate-buffered saline |
| PCL | poly-ϵ-caprolactone |
| PHB | polyhydroxybutyrate |
| PHBV | PHB-co-hydroxyvalerate |
| PLA | polylactide |
| PVA | polyvinyl alcohol |
| RANK | receptor activator of nuclear factor κB |
| RANKL | receptor activator of nuclear factor κB ligand |
| SMC | smooth muscle cell |
| SNP | single nucleotide polymorphism |
| TAK1 | transforming growth factor-β-activated kinase 1 |
| TISA | thermally induced nanofiber self-agglomeration |
| TRAF6 | tumor necrosis factor receptor-associated factor 6 |
References
- Tomar, K.; Sahoo, N.K. Evaluation of graft uptake from the iliac crest in secondary alveolar bone grafting: Bergland’s criteria revisited. J. Oral Biol. Craniofac. Res. 2018, 8, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Egol, K.A.; Nauth, A.; Lee, M.; Pape, H.C.; Watson, J.T.; Borrelli, J., Jr. Bone Grafting: Sourcing, Timing, Strategies, and Alternatives. J. Orthop. Trauma. 2015, 29 (Suppl. 12), S510–S514. [Google Scholar] [CrossRef] [PubMed]
- Batista, J.D.; Justino Oliveira Limirio, P.H.; Rocha, F.S.; Gomes Moura, C.C.; Zanetta-Barbosa, D.; Dechichi, P. Influence of Platelet-Poor Plasma on Angiogenesis and Maintenance of Volume in Autogenous Bone Grafts. J. Oral Maxillofac. Surg. 2018. [Google Scholar] [CrossRef] [PubMed]
- Burk, T.; Del Valle, J.; Finn, R.A.; Phillips, C. Maximum Quantity of Bone Available for Harvest From the Anterior Iliac Crest, Posterior Iliac Crest, and Proximal Tibia Using a Standardized Surgical Approach: A Cadaveric Study. J. Oral Maxillofac. Surg. 2016, 74, 2532–2548. [Google Scholar] [CrossRef] [PubMed]
- Kilinc, A.; Korkmaz, I.H.; Kaymaz, I.; Kilinc, Z.; Dayi, E.; Kantarci, A. Comprehensive analysis of the volume of bone for grafting that can be harvested from iliac crest donor sites. Br. J. Oral Maxillofac. Surg. 2017, 55, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; He, W.; Yang, W.; Li, J.; Yang, Z.; Lu, X.; Qin, A.; Qian, Y. The osteogenic potential of human bone callus. Sci. Rep. 2016, 6, 36330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekker, T.J.; White, P.; Adams, S.B. Efficacy of a Cellular Bone Allograft for Foot and Ankle Arthrodesis and Revision Nonunion Procedures. Foot Ankle Int. 2017, 38, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, D.R.; Ortiz-Declet, V.; Chen, A.W.; Lall, A.C.; Mohr, M.R.; Laseter, J.R.; Domb, B.G. Modified Shelf Acetabuloplasty Endoscopic Procedure With Allograft for Developmental Hip Dysplasia Treatment. Arthrosc. Tech. 2018, 7, e779–e784. [Google Scholar] [CrossRef] [PubMed]
- Fishman, J.A.; Greenwald, M.A.; Grossi, P.A. Transmission of infection with human allografts: Essential considerations in donor screening. Clin. Infect. Dis. 2012, 55, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Stopa, Z.; Siewert-Gutowska, M.; Abed, K.; Szubińska-Lelonkiewicz, D.; Kamiński, A.; Fiedor, P. Evaluation of the Safety and Clinical Efficacy of Allogeneic Bone Grafts in the Reconstruction of the Maxilla and Mandible. Transplant. Proc. 2018, 50, 2199–2201. [Google Scholar] [CrossRef] [PubMed]
- El-Sherbiny, Y.M.; El-Jawhari, J.J.; Moseley, T.A.; McGonagle, D.; Jones, E. T cell immunomodulation by clinically used allogeneic human cancellous bone fragments: A potential novel immunotherapy tool. Sci. Rep. 2018, 8, 13535. [Google Scholar] [CrossRef] [PubMed]
- Gharedaghi, M.; Peivandi, M.T.; Mazloomi, M.; Shoorin, H.R.; Hasani, M.; Seyf, P.; Khazaee, F. Evaluation of Clinical Results and Complications of Structural Allograft Reconstruction after Bone Tumor Surgery. Arch. Bone Jt. Surg. 2016, 4, 236–242. [Google Scholar] [PubMed]
- Bagherifard, A.; Ghandhari, H.; Jabalameli, M.; Rahbar, M.; Hadi, H.; Moayedfar, M.; Sajadi, M.M.; Karimpour, A. Autograft versus allograft reconstruction of acute tibial plateau fractures: A comparative study of complications and outcome. Eur. J. Orthop. Surg. Traumatol. 2017, 27, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Zamborsky, R.; Svec, A.; Bohac, M.; Kilian, M.; Kokavec, M. Infection in Bone Allograft Transplants. Exp. Clin. Transplant 2016, 14, 484–490. [Google Scholar] [PubMed]
- Singh, R.; Singh, D.; Singh, A. Radiation sterilization of tissue allografts: A review. World J. Radiol. 2016, 8, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Chavda, S.; Levin, L. Human Studies of Vertical and Horizontal Alveolar Ridge Augmentation Comparing Different Types of Bone Graft Materials: A Systematic Review. J. Oral Implantol. 2018, 44, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Bracey, D.N.; Seyler, T.M.; Jinnah, A.H.; Lively, M.O.; Willey, J.S.; Smith, T.L.; Van Dyke, M.E.; Whitlock, P.W. A Decellularized Porcine Xenograft-Derived Bone Scaffold for Clinical Use as a Bone Graft Substitute: A Critical Evaluation of Processing and Structure. J. Funct. Biomater. 2018, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Lee, J.M.; Tobiasch, E.; Csizmadia, E.; Smith, N.R.; Gollackes, B.; Robson, S.C.; Bach, F.H.; Lin, Y. Induction of xenograft accommodation by modulation of elicited antibody responses1 2. Transplantation 2002, 74, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.R.; Martens, G.R.; Estrada, J.L.; Reyes, L.M.; Ladowski, J.M.; Galli, C.; Perota, A.; Cunningham, C.M.; Tector, M.; Joseph Tector, A. Silencing porcine genes significantly reduces human-anti-pig cytotoxicity profiles: An alternative to direct complement regulation. Transgenic Res. 2016, 25, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Koo, T.Y.; Lee, H.S.; Lee, W.B.; Kang, B.; Lee, J.G.; Jang, J.Y.; Fang, T.; Ryu, J.H.; Ahn, C.; et al. Role of Human CD200 Overexpression in Pig-to-Human Xenogeneic Immune Response Compared With Human CD47 Overexpression. Transplantation 2018, 102, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Kivovics, M.; Szabó, B.T.; Németh, O.; Tari, N.; Dőri, F.; Nagy, P.; Dobó-Nagy, C.; Szabó, G. Microarchitectural study of the augmented bone following ridge preservation with a porcine xenograft and a collagen membrane: Preliminary report of a prospective clinical, histological, and micro-computed tomography analysis. Int. J. Oral Maxillofac. Surg. 2017, 46, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Khan, D.; Delling, J.; Tobiasch, E. Mechanisms underlying the osteo- and adipo-differentiation of human mesenchymal stem cells. Sci. World. J. 2012, 2012, 793823. [Google Scholar] [CrossRef] [PubMed]
- Coelho de Faria, A.B.; Chiantia, F.B.; Teixeira, M.L.; Aloise, A.C.; Pelegrine, A.A. Comparative Study Between Mesenchymal Stem Cells Derived from Bone Marrow and from Adipose Tissue, Associated with Xenograft, in Appositional Reconstructions: Histomorphometric Study in Rabbit Calvaria. Int. J. Oral Maxillofac. Implants 2016, 31, e155–e161. [Google Scholar] [CrossRef] [PubMed]
- Tobiasch, E. Differentiation Potential of Adult Human Mesenchymal Stem Cells. In Stem Cell Engineering; Artmann, G.M., Hescheler, J., Minger, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 61–76. [Google Scholar]
- Victorelli, G.; Aloise, A.C.; Passador-Santos, F.; de Mello e Oliveira, R.; Pelegrine, A.A. Ectopic Implantation of Hydroxyapatite Xenograft Scaffold Loaded with Bone Marrow Aspirate Concentrate or Osteodifferentiated Bone Marrow Mesenchymal Stem Cells: A Pilot Study in Mice. Int. J. Oral Maxillofac. Implants 2016, 31, e18–e23. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.; Tobiasch, E. Artificial scaffolds and mesenchymal stem cells for hard tissues. In Tissue Engineering III: Cell-Surface Interactions for Tissue Culture; Kasper, C., Witte, F., Pörtner, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 126, pp. 153–194. [Google Scholar]
- Šponer, P.; Kučera, T.; Brtková, J.; Urban, K.; Kočí, Z.; Měřička, P.; Bezrouk, A.; Konrádová, Š.; Filipová, A.; Filip, S. Comparative Study on the Application of Mesenchymal Stromal Cells Combined with Tricalcium Phosphate Scaffold into Femoral Bone Defects. Cell Transplant 2018, 11, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Zippel, N.; Schulze, M.; Tobiasch, E. Biomaterials and mesenchymal stem cells for regenerative medicine. Recent Pat Biotechnol. 2010, 4, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Garai, S.; Sinha, A. Three dimensional biphasic calcium phosphate nanocomposites for load bearing bioactive bone grafts. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 59, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Buser, Z.; Brodke, D.S.; Youssef, J.A.; Meisel, H.J.; Myhre, S.L.; Hashimoto, R.; Park, J.B.; Tim Yoon, S.; Wang, J.C. Synthetic bone graft versus autograft or allograft for spinal fusion: A systematic review. J. Neurosurg. Spine 2016, 25, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Khaldi-Hansen, E.; El-Sayed, F.; Schipper, D.; Tobiasch, E.; Witzleben, S.; Schulze, M. Functionalized 3D Scaffolds for Templatemediated Biomineralization. In Bone Regeneration; Rhaman, A., Ed.; Bentham Science Publishers: Sharjah, United Arab Emirates, 2017; Volume 5, pp. 130–178. [Google Scholar]
- Schipper, D.; Babczyk, P.; El-Sayed, F.; Klein, S.E.; Schulze, M.; Tobiasch, E. The Effect of Nanostructured Surfaces on Stem Cell Fate. In Set Nanostructures for Novel Therapy; Grumezescu, A.M., Ficai, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1, pp. 567–589. [Google Scholar]
- Pansky, A.; Roitzheim, B.; Tobiasch, E. Differentiation potential of adult of human mesenchymal stem cells. Clin. Lab. 2007, 53, 81–84. [Google Scholar] [PubMed]
- Haddouti, E.-M.; Skroch, M.; Zippel, N.; Müller, C.; Birova, B.; Pansky, A.; Kleinfeld, C.; Winter, M.; Tobiasch, E. Human Dental Follicle Precursor Cells of Wisdom Teeth: Isolation and Differentiation towards Osteoblasts for Implants with and without Scaffolds. Mater. Sci. Eng. Technol. 2009, 40, 732–737. [Google Scholar] [CrossRef]
- Grotheer, V.; Schulze, M.; Tobiasch, E. Trends in Bone Tissue Engineering: Proteins for Osteogenic Differentiation and the Respective Scaffolding. In Protein Purification—Principles and Trends; iConcept Press Ltd.: Hong Kong, China, 2016. [Google Scholar]
- Hernigou, P.; Dubory, A.; Roubineau, F.; Homma, Y.; Flouzat-Lachaniette, C.H.; Chevallier, N.; Rouard, H. Allografts supercharged with bone-marrow-derived mesenchymal stem cells possess equivalent osteogenic capacity to that of autograft: A study with long-term follow-ups of human biopsies. Int. Orthop. 2017, 41, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Thangarajah, T.; Shahbazi, S.; Pendegrass, C.J.; Lambert, S.; Alexander, S.; Blunn, G.W. Tendon Reattachment to Bone in an Ovine Tendon Defect Model of Retraction Using Allogenic and Xenogenic Demineralised Bone Matrix Incorporated with Mesenchymal Stem Cells. PLoS ONE 2016, 11, e0161473. [Google Scholar] [CrossRef] [PubMed]
- El-Jawhari, J.J.; Jones, E.; Giannoudis, P.V. The roles of immune cells in bone healing; what we know, do not know and future perspectives. Injury 2016, 47, 2399–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.; Zhou, Z.; Ju, X.; Zhou, Y.; Lan, J.; Chen, D.; Chen, H.; Liu, M.; Pang, L. Combined transplantation of mesenchymal stem cells and endothelial progenitor cells for tissue engineering: A systematic review and meta-analysis. Stem Cell Res. Ther. 2016, 7, 151. [Google Scholar] [CrossRef] [PubMed]
- Babczyk, P.; Conzendorf, C.; Klose, J.; Schulze, M.; Harre, K.; Tobiasch, E. Stem Cells on Biomaterials for Synthetic Grafts to Promote Vascular Healing. J. Clin. Med. 2014, 3, 39–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. Intravital Imaging to Understand Spatiotemporal Regulation of Osteogenesis and Angiogenesis in Cranial Defect Repair and Regeneration. Methods Mol. Biol. 2018, 1842, 229–239. [Google Scholar] [PubMed]
- Seifert, A.; Knapp, S.; Werheid, D.; Tobiasch, E. Role of Hox Genes in Stem Cell Differentiation. World J. Stem Cells 2015, 7, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Picke, A.K.; Campbell, G.M.; Blüher, M.; Krügel, U.; Schmidt, F.N.; Tsourdi, E.; Winzer, M.; Rauner, M.; Vukicevic, V.; Busse, B.; et al. Thy-1 (CD90) promotes bone formation and protects against obesity. Sci. Transl. Med. 2018, 10, eaao6806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tobiasch, E. The role of purinergic receptors in stem cells in their derived consecutive tissues. In Adult Stem Cell Standardization; River Publishers: Roma, Italy, 2011; pp. 73–98. [Google Scholar]
- Kaebisch, C.; Schipper, D.; Babczyk, P.; Tobiasch, E. The role of purinergic receptors in stem cell differentiation. Comput. Struct. Biotechnol. J. 2014, 13, 75–84. [Google Scholar] [CrossRef]
- Burnstock, G.; Verkhratsky, A. Evolutionary origins of the purinergic signalling system. Acta Physiol. (Oxf.) 2009, 195, 415–447. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purine and pyrimidine receptors. Cell Mol. Life Sci. 2007, 64, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Thimm, D.; Schiedel, A.C.; Peti-Peterdi, J.; Kishore, B.K.; Müller, C.E. The nucleobase adenine as a signalling molecule in the kidney. Acta Physiol. (Oxf.) 2015, 213, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Abbracchio, M.P.; Burnstock, G.; Boeynaems, J.M.; Barnard, E.A.; Boyer, J.L.; Kennedy, C.; Knight, G.E.; Fumagalli, M.; Gachet, C.; Jacobson, K.A.; et al. International Union of Pharmacology LVIII: Update on the P2Y G protein-coupled nucleotide receptors: From molecular mechanisms and pathophysiology to therapy. Pharmacol. Rev. 2006, 58, 281–341. [Google Scholar] [CrossRef] [PubMed]
- Borrmann, T.; Abdelrahman, A.; Volpini, R.; Lambertucci, C.; Alksnis, E.; Gorzalka, S.; Knospe, M.; Schiedel, A.C.; Cristalli, G.; Müller, C.E. Structure-activity relationships of adenine and deazaadenine derivatives as ligands for adenine receptors, a new purinergic receptor family. J. Med. Chem. 2009, 52, 5974–5989. [Google Scholar] [CrossRef] [PubMed]
- Bender, E.; Buist, A.; Jurzak, M.; Langlois, X.; Baggerman, G.; Verhasselt, P.; Ercken, M.; Guo, H.Q.; Wintmolders, C.; Van den Wyngaert, I.; et al. Characterization of an orphan G protein-coupled receptor localized in the dorsal root ganglia reveals adenine as a signaling molecule. Proc. Natl. Acad. Sci. USA 2002, 99, 8573–8578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wengert, M.; Adão-Novaes, J.; Assaife-Lopes, N.; Leão-Ferreira, L.R.; Caruso-Neves, C. Adenine-induced inhibition of Na(+)-ATPase activity: Evidence for involvement of the Gi protein-coupled receptor in the cAMP signaling pathway. Arch. Biochem. Biophys. 2007, 467, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Gorzalka, S.; Vittori, S.; Volpini, R.; Cristalli, G.; von Kügelgen, I.; Müller, C.E. Evidence for the functional expression and pharmacological characterization of adenine receptors in native cells and tissues. Mol. Pharmacol. 2005, 67, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; Ijzerman, A.P.; Jacobson, K.A.; Linden, J.; Müller, C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and classification of adenosine receptors--an update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, F.; Felline, A. Dimerization and ligand binding affect the structure network of A(2A) adenosine receptor. Biochim. Biophys. Acta 2011, 1808, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.; Malavasi, F.; Antonioli, L. A Purinergic Trail for Metastases. Trends Pharmacol. Sci. 2017, 38, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Ham, J.; Evans, B.A. An emerging role for adenosine and its receptors in bone homeostasis. Front. Endocrinol. (Lausanne) 2012, 3, 113. [Google Scholar] [CrossRef] [PubMed]
- North, R.A. P2X receptors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.H.; Hao, Y.; Mousawi, F.; Peng, H.; Yang, X. Expression of P2 Purinergic Receptors in Mesenchymal Stem Cells and Their Roles in Extracellular Nucleotide Regulation of Cell Functions. J. Cell Physiol. 2017, 232(2), 287–297. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek-Hájek, K.; Lörinczi, E.; Hausmann, R.; Nicke, A. Molecular and functional properties of P2X receptors--recent progress and persisting challenges. Purinergic Signal. 2012, 8, 375–417. [Google Scholar] [CrossRef] [PubMed]
- Torres, G.E.; Egan, T.M.; Voigt, M.M. Hetero-oligomeric assembly of P2X receptor subunits. Specificities exist with regard to possible partners. J. Biol. Chem. 1999, 274, 6653–6659. [Google Scholar] [CrossRef] [PubMed]
- Barrera, N.P.; Ormond, S.J.; Henderson, R.M.; Murrell-Lagnado, R.D.; Edwardson, J.M. Atomic force microscopy imaging demonstrates that P2X2 receptors are trimers but that P2X6 receptor subunits do not oligomerize. J. Biol. Chem. 2005, 280, 10759–10765. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Villablanca, E.J. Retinoic acid takes effector T cells to the gallows: P2X7, the molecular hangman. Mucosal. Immunol. 2017, 10, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Henriksen, Z.; Syberg, S.; Petersen, S.; Aslan, D.; Solgaard, M.; Nissen, N.; Larsen, T.K.; Schwarz, P.; Steinberg, T.H.; et al. P2X7Rs are involved in cell death, growth and cellular signaling in primary human osteoblasts. Bone 2017, 95, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Gicquel, T.; Le Daré, B.; Boichot, E.; Lagente, V. Purinergic receptors: New targets for the treatment of gout and fibrosis. Fundam. Clin. Pharmacol. 2017, 31, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Zippel, N.; Limbach, C.A.; Ratajski, N.; Urban, C.; Luparello, C.; Pansky, A.; Kassack, M.U.; Tobiasch, E. Purinergic receptors influence the differentiation of human mesenchymal stem cells. Stem Cells Dev. 2012, 21, 884–900. [Google Scholar] [CrossRef] [PubMed]
- Gardinier, J.D.; Gangadharan, V.; Wang, L.; Duncan, R.L. Hydraulic Pressure during Fluid Flow Regulates Purinergic Signaling and Cytoskeleton Organization of Osteoblasts. Cell Mol. Bioeng. 2014, 7, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Boeynaems, J.M.; Communi, D.; Gonzalez, N.S.; Robaye, B. Overview of the P2 receptors. Semin. Thromb. Hemost. 2005, 31, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Webb, T.E.; Barnard, E.A. Characterization of a P2Y purinoceptor in the brain. Pharmacol. Toxicol. 1995, 76, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Guo, D.; Lee, B.X.; Van Rhee, A.M.; Kim, Y.C.; Nicholas, R.A.; Schachter, J.B.; Harden, T.K.; Jacobson, K.A. A mutational analysis of residues essential for ligand recognition at the human P2Y1 receptor. Mol. Pharmacol. 1997, 52, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Erb, L.; Cao, C.; Ajit, D.; Weisman, G.A. P2Y receptors in Alzheimer’s disease. Biol. Cell 2015, 107, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic Signalling: Therapeutic Developments. Front. Pharmacol. 2017, 8, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biver, G.; Wang, N.; Gartland, A.; Orriss, I.; Arnett, T.R.; Boeynaems, J.M.; Robaye, B. Role of the P2Y13 receptor in the differentiation of bone marrow stromal cells into osteoblasts and adipocytes. Stem Cells 2013, 31, 2747–2758. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Doss, C.G. Crucial protein based drug targets and potential inhibitors for osteoporosis: New hope and possibilities. Curr. Drug Targets 2013, 14, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Glaser, T.; Cappellari, A.R.; Pillat, M.M.; Iser, I.C.; Wink, M.R.; Battastini, A.M.; Ulrich, H. Perspectives of purinergic signaling in stem cell differentiation and tissue regeneration. Purinergic Signal. 2012, 8, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yeung, K.W.K. Bone grafts and biomaterials substitutes for bone defect repair: A review. Bioact. Mater. 2017, 2, 224–247. [Google Scholar] [CrossRef] [PubMed]
- Lenertz, L.Y.; Baughman, C.J.; Waldschmidt, N.V.; Thaler, R.; van Wijnen, A.J. Control of bone development by P2X and P2Y receptors expressed in mesenchymal and hematopoietic cells. Gene 2015, 570, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, S.; Wang, C.; Levene, H.; Cheung, H.S.; Huang, C.Y. ATP promotes extracellular matrix biosynthesis of intervertebral disc cells. Cell Tissue Res. 2015, 359, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Takedachi, M.; Oohara, H.; Smith, B.J.; Iyama, M.; Kobashi, M.; Maeda, K.; Long, C.L.; Humphrey, M.B.; Stoecker, B.J.; Toyosawa, S.; et al. CD73-generated adenosine promotes osteoblast differentiation. J. Cell Physiol. 2012, 227, 2622–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noronha-Matos, J.B.; Costa, M.A.; Magalhães-Cardoso, M.T.; Ferreirinha, F.; Pelletier, J.; Freitas, R.; Neves, J.M.; Sévigny, J.; Correia-de-Sá, P. Role of ecto-NTPDases on UDP-sensitive P2Y(6) receptor activation during osteogenic differentiation of primary bone marrow stromal cells from postmenopausal women. J. Cell Physiol. 2012, 227, 2694–2709. [Google Scholar] [CrossRef] [PubMed]
- Manaka, S.; Tanabe, N.; Kariya, T.; Naito, M.; Takayama, T.; Nagao, M.; Liu, D.; Ito, K.; Maeno, M.; Suzuki, N.; et al. Low-intensity pulsed ultrasound-induced ATP increases bone formation via the P2X7 receptor in osteoblast-like MC3T3-E1 cells. FEBS Lett. 2015, 589, 310–318. [Google Scholar] [CrossRef] [PubMed]
- D’Alimonte, I.; Nargi, E.; Lannutti, A.; Marchisio, M.; Pierdomenico, L.; Costanzo, G.; Iorio, P.D.; Ballerini, P.; Giuliani, P.; Caciagli, F.; et al. Adenosine A1 receptor stimulation enhances osteogenic differentiation of human dental pulp-derived mesenchymal stem cells via WNT signaling. Stem Cell Res. 2013, 11, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Eijken, M.; Meijer, I.M.; Westbroek, I.; Koedam, M.; Chiba, H.; Uitterlinden, A.G.; Pols, H.A.; van Leeuwen, J.P. Wnt signaling acts and is regulated in a human osteoblast differentiation dependent manner. J. Cell Biochem. 2008, 104, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Khan, D.; Kleinfeld, C.; Winter, M.; Tobiasch, E. Oral Tissues as Source for Bone Regeneration in Dental Implantology. In Tissue Regeneration. From Basic Biology to Clinical Application; InTech Open: London, UK, 2012; ISBN 978-953-307-876-2. [Google Scholar] [Green Version]
- Mediero, A.; Wilder, T.; Perez-Aso, M.; Cronstein, B.N. Direct or indirect stimulation of adenosine A2A receptors enhances bone regeneration as well as bone morphogenetic protein-2. FASEB J. 2015, 29, 1577–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.A.; Barbosa, A.; Neto, E.; Sá-e-Sousa, A.; Freitas, R.; Neves, J.M.; Magalhães-Cardoso, T.; Ferreirinha, F.; Correia-de-Sá, P. On the role of subtype selective adenosine receptor agonists during proliferation and osteogenic differentiation of human primary bone marrow stromal cells. J. Cell Physiol. 2011, 226, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.H.; Wigner, N.A.; Kulkarni, N.; Johnston-Cox, H.; Gerstenfeld, L.C.; Ravid, K. A2B adenosine receptor promotes mesenchymal stem cell differentiation to osteoblasts and bone formation in vivo. J. Biol. Chem. 2012, 287, 15718–15727. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Kim, M.; Feng, Q.; Lin, S.; Wei, K.; Li, R.; Choi, C.J.; Kim, T.H.; Li, G.; Oh, J.M.; et al. Nanolayered hybrid mediates synergistic co-delivery of ligand and ligation activator for inducing stem cell differentiation and tissue healing. Biomaterials 2017, 149, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Corciulo, C.; Wilder, T.; Cronstein, B.N. Adenosine A2B receptors play an important role in bone homeostasis. Purinergic Signal. 2016, 12, 537–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trincavelli, M.L.; Daniele, S.; Giacomelli, C.; Taliani, S.; Da Settimo, F.; Cosimelli, B.; Greco, G.; Novellino, E.; Martini, C. Osteoblast differentiation and survival: A role for A2B adenosine receptor allosteric modulators. Biochim. Biophys. Acta 2014, 1843, 2957–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajjawi, M.O.; Patel, J.J.; Corcelli, M.; Arnett, T.R.; Orriss, I.R. Lack of effect of adenosine on the function of rodent osteoblasts and osteoclasts in vitro. Purinergic Signal. 2016, 12, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, D.; Gulinelli, S.; Salvestrini, V.; Lucchetti, G.; Zini, R.; Manfredini, R.; Caione, L.; Piacibello, W.; Ciciarello, M.; Rossi, L.; et al. Purinergic stimulation of human mesenchymal stem cells potentiates their chemotactic response to CXCL12 and increases the homing capacity and production of proinflammatory cytokines. Exp. Hematol. 2011, 39, 360–374. [Google Scholar] [CrossRef] [PubMed]
- Orriss, I.R.; Key, M.L.; Hajjawi, M.O.; Arnett, T.R. Extracellular ATP released by osteoblasts is a key local inhibitor of bone mineralisation. PLoS ONE 2013, 8, e69057. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Junger, W.G.; Yuan, C.; Zhang, W.; Bao, Y.; Qin, D.; Wang, C.; Tan, L.; Qi, B.; Zhu, D.; et al. Shockwaves induce osteogenic differentiation of human mesenchymal stem cells through ATP release and activation of P2X7 receptors. Stem Cells 2013, 31, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, G.; Zhang, Y.; Wei, S.; Song, M.; Wang, W.; Yuan, X.; Wu, H.; Yang, Y. Role of P2 × 7 receptor in the differentiation of bone marrow stromal cells into osteoblasts and adipocytes. Exp. Cell Res. 2015, 339, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Grol, M.W.; Brooks, P.J.; Pereverzev, A.; Dixon, S.J. P2X7 nucleotide receptor signaling potentiates the Wnt/β-catenin pathway in cells of the osteoblast lineage. Purinergic Signal. 2016, 12, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noronha-Matos, J.B.; Coimbra, J.; Sá-e-Sousa, A.; Rocha, R.; Marinhas, J.; Freitas, R.; Guerra-Gomes, S.; Ferreirinha, F.; Costa, M.A.; Correia-de-Sá, P. P2X7-induced zeiosis promotes osteogenic differentiation and mineralization of postmenopausal bone marrow-derived mesenchymal stem cells. FASEB J. 2014, 28, 5208–5222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues-Ribeiro, R.; Alvarenga, É.C.; Calio, M.L.; Paredes-Gamero, E.J.; Ferreira, A.T. Dual role of P2 receptors during osteoblast differentiation. Cell Biochem. Biophys. 2015, 71, 1225–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orriss, I.R. The role of purinergic signalling in the musculoskeletal system. Auton. Neurosci. 2015, 191, 124–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Wei, S.; Liu, C.; Song, M.; Wu, H.; Yang, Y. Regulation of the osteogenic and adipogenic differentiation of bone marrow-derived stromal cells by extracellular uridine triphosphate: The role of P2Y2 receptor and ERK1/2 signaling. Int. J. Mol. Med. 2016, 37, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Gu, Y.; Bresnahan, J.J.; Paul, E.M.; Donahue, H.J.; You, J. The roles of P2Y2 purinergic receptors in osteoblasts and mechanotransduction. PLoS ONE 2014, 9, e108417. [Google Scholar] [CrossRef] [PubMed]
- Syberg, S.; Brandao-Burch, A.; Patel, J.J.; Hajjawi, M.; Arnett, T.R.; Schwarz, P.; Jorgensen, N.R.; Orriss, I.R. Clopidogrel (Plavix), a P2Y12 receptor antagonist, inhibits bone cell function in vitro and decreases trabecular bone in vivo. J. Bone Miner Res. 2012, 27, 2373–2386. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Floyd, D.H.; Hughes, A.; Xiang, J.; Schneider, J.G.; Uluckan, O.; Heller, E.; Deng, H.; Zou, W.; Craft, C.S.; et al. The ADP receptor P2RY12 regulates osteoclast function and pathologic bone remodeling. J. Clin. Invest. 2012, 122, 3579–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesselius, A.; Bours, M.J.; Henriksen, Z.; Syberg, S.; Petersen, S.; Schwarz, P.; Jørgensen, N.R.; van Helden, S.; Dagnelie, P.C. Association of P2Y(2) receptor SNPs with bone mineral density and osteoporosis risk in a cohort of Dutch fracture patients. Purinergic Signal. 2013, 9, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Gartland, A.; Skarratt, K.K.; Hocking, L.J.; Parsons, C.; Stokes, L.; Jørgensen, N.R.; Fraser, W.D.; Reid, D.M.; Gallagher, J.A.; Wiley, J.S. Polymorphisms in the P2X7 receptor gene are associated with low lumbar spine bone mineral density and accelerated bone loss in post-menopausal women. Eur. J. Hum. Genet. 2012, 20, 559–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strazzulla, L.C.; Cronstein, B.N. Regulation of bone and cartilage by adenosine signaling. Purinergic Signal. 2016, 12, 583–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegatti, P.; Falzoni, S.; Donvito, G.; Lemaire, I.; Di Virgilio, F. P2X7 receptor drives osteoclast fusion by increasing the extracellular adenosine concentration. FASEB J. 2011, 25, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- Mediero, A.; Frenkel, S.R.; Wilder, T.; He, W.; Mazumder, A.; Cronstein, B.N. Adenosine A2A receptor activation prevents wear particle-induced osteolysis. Sci. Transl. Med. 2012, 4, 135ra65. [Google Scholar] [CrossRef] [PubMed]
- Kara, F.M.; Chitu, V.; Sloane, J.; Axelrod, M.; Fredholm, B.B.; Stanley, E.R.; Cronstein, B.N. Adenosine A1 receptors (A1Rs) play a critical role in osteoclast formation and function. FASEB J. 2010, 24, 2325–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Cronstein, B.N. Adenosine A1 receptor regulates osteoclast formation by altering TRAF6/TAK1 signaling. Purinergic Signal. 2012, 8, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Cronstein, B. The roles of adenosine and adenosine receptors in bone remodeling. Front Biosci. (Elite Ed) 2011, 3, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Oh, J.H.; Lee, N.K. The Inactivation of ERK1/2, p38 and NF-kB Is Involved in the Down-Regulation of Osteoclastogenesis and Function by A2B Adenosine Receptor Stimulation. Mol. Cells 2017, 40, 752–760. [Google Scholar] [PubMed]
- Orriss, I.R.; Burnstock, G.; Arnett, T.R. Purinergic signalling and bone remodelling. Curr. Opin. Pharmacol. 2010, 10, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Naemsch, L.N.; Weidema, A.F.; Sims, S.M.; Underhill, T.M.; Dixon, S.J. P2X(4) purinoceptors mediate an ATP-activated, non-selective cation current in rabbit osteoclasts. J. Cell Sci. 1999, 112, 4425–4435. [Google Scholar] [PubMed]
- Wang, N.; Agrawal, A.; Jørgensen, N.R.; Gartland, A. P2X7 receptor regulates osteoclast function and bone loss in a mouse model of osteoporosis. Sci. Rep. 2018, 8, 3507. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Gartland, A. P2X7 receptors: Role in bone cell formation and function. J. Mol. Endocrinol. 2015, 54, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Falzoni, S.; Munerati, M.; Ferrari, D.; Spisani, S.; Moretti, S.; Di Virgilio, F. The purinergic P2Z receptor of human macrophage cells. Characterization and possible physiological role. J. Clin. Invest. 1995, 95, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Iwasawa, M.; Nakashima, T.; Mori, S.; Shigemoto, K.; Nakamura, H.; Katagiri, H.; Takayanagi, H.; Tanaka, S. Intracellular and extracellular ATP coordinately regulate the inverse correlation between osteoclast survival and bone resorption. J. Biol. Chem. 2012, 287, 37808–37823. [Google Scholar] [CrossRef] [PubMed]
- Kohl, F.; Schmitz, J.; Furtmann, N.; Schulz-Fincke, A.-C.; Mertens, M.D.; Küppers, J.; Benkhoff, M.; Tobiasch, E.; Bartz, U.; Bajorath, J.; et al. Design, characterization and cellular uptake studies of fluorescence-labeled prototypic cathepsin inhibitors. Org. Biomol. Chem. 2015, 13, 10310–10323. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Lim, H.; Lee, D.S.; Yim, M. Montelukast inhibits RANKL-induced osteoclast formation and bone loss via CysLTR1 and P2Y12. Mol. Med. Rep. 2018, 18, 2387–2398. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Robaye, B.; Agrawal, A.; Skerry, T.M.; Boeynaems, J.M.; Gartland, A. Reduced bone turnover in mice lacking the P2Y13 receptor of ADP. Mol. Endocrinol. 2012, 26, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Park, J.H.; Lee, S.Y. Selective induction of P2Y14 receptor by RANKL promotes osteoclast formation. Mol. Cell. 2013, 36, 273–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hielscher, D.; Kaebisch, C.; Braun, B.J.V.; Gray, K.; Tobiasch, E. Stem Cell Sources and Graft Material for Vascular Tissue Engineering. Stem Cell Rev. 2018, 14, 642–667. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Yang, X.; Li, M.; Huang, H.; Kang, Q.; Zhang, X.; Hui, H.; Zhang, X.; Chen, C.; Luo, Y.; et al. A novel strategy for in vivo angiogenesis and osteogenesis: Magnetic micro-movement in a bone scaffold. Artif Cell. Nanomed. Biotechnol. 2018, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.N.; Youkey, R.; Liu, X.; Jia, L.; Blatt, R.; Day, Y.J.; Sullivan, G.W.; Linden, J.; Tucker, A.L. A1 adenosine receptor activation promotes angiogenesis and release of VEGF from monocytes. Circ. Res. 2007, 101, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Koszałka, P.; Gołuńska, M.; Urban, A.; Stasiłojć, G.; Stanisławowski, M.; Majewski, M.; Składanowski, A.C.; Bigda, J. Specific Activation of A3, A2A and A1 Adenosine Receptors in CD73-Knockout Mice Affects B16F10 Melanoma Growth, Neovascularization, Angiogenesis and Macrophage Infiltration. PLoS ONE 2016, 11, e0151420. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G.; Auvinen, K.; Rantakari, P.; Hollmén, M.; Karikoski, M.; Grénman, R.; Elima, K.; Jalkanen, S.; Salmi, M. Ecto-5’-nucleotidase/CD73 enhances endothelial barrier function and sprouting in blood but not lymphatic vasculature. Eur. J. Immunol. 2015, 45, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, P.; Jara, C.; Aguilera, V.; Caviedes, L.; Diaz, F.; Radojkovic, C.; Veas, C.; Lamperti, L.; Escudero, C.; Aguayo, C. Adenosine A₂A and A₃ receptors are involved in the human endothelial progenitor cells migration. J. Cardiovasc. Pharmacol. 2012, 59, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Ou, X.; Song, T.; Zhang, W.; Cong, F.; Zhang, S.; Xiong, Y. Adenosine A2B receptor stimulates angiogenesis by inducing VEGF and eNOS in human microvascular endothelial cells. Exp. Biol. Med. (Maywood) 2015, 240, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lau, P.; Pansky, A.; Kassack, M.; Hemmersbach, R.; Tobiasch, E. The influence of simulated microgravity on purinergic signaling is different between individual culture and endothelial and smooth muscle cell coculture. Biomed. Res. Int. 2014, 2014, 413708. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Karlsson, L.; Moses, S.; Hultgårdh-Nilsson, A.; Andersson, M.; Borna, C.; Gudbjartsson, T.; Jern, S.; Erlinge, D. P2 receptor expression profiles in human vascular smooth muscle and endothelial cells. J. Cardiovasc. Pharmacol. 2002, 40, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Chen, X.; Wang, S.; Xie, T.; Du, X.; Liu, H.; Wang, S.; Li, X.; Chen, J.; Zhang, B.; et al. The expression of P2X₇ receptors in EPCs and their potential role in the targeting of EPCs to brain gliomas. Cancer Biol. Ther. 2015, 16, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Brouard, S.; Otterbein, L.E.; Anrather, J.; Tobiasch, E.; Bach, F.H.; Choi, A.M.; Soares, M.P. Carbon monoxide generated by heme oxygenase 1 suppresses endothelial cell apoptosis. J. Exp. Med. 2000, 192, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Brouard, S.; Berberat, P.O.; Tobiasch, E.; Seldon, M.P.; Bach, F.H.; Soares, M.P. Heme oxygenase-1-derived carbon monoxide requires the activation of transcription factor NF-kappa B to protect endothelial cells from tumor necrosis factor-alpha-mediated apoptosis. J. Biol. Chem. 2002, 277, 17950–17961. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.P.; Usheva, A.; Brouard, S.; Berberat, P.O.; Gunther, L.; Tobiasch, E.; Bach, F.H. Modulation of endothelial cell apoptosis by heme oxygenase-1-derived carbon monoxide. Antioxid. Redox Signal. 2002, 4, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; DiCorleto, P.E. ADP stimulates human endothelial cell migration via P2Y1 nucleotide receptor-mediated mitogen-activated protein kinase pathways. Circ. Res. 2008, 102, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Lyubchenko, T.; Woodward, H.; Veo, K.D.; Burns, N.; Nijmeh, H.; Liubchenko, G.A.; Stenmark, K.R.; Gerasimovskaya, E.V. P2Y1 and P2Y13 purinergic receptors mediate Ca2+ signaling and proliferative responses in pulmonary artery vasa vasorum endothelial cells. Am. J. Physiol. Cell Physiol. 2011, 300, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Stoeckius, M.; Erat, A.; Fujikawa, T.; Hiromura, M.; Koulova, A.; Otterbein, L.; Bianchi, C.; Tobiasch, E.; Dagon, Y.; Sellke, F.W.; et al. Essential roles of Raf/ERK/MAPK pathway, YY1, and Ca+2 influx in the growth arrest of human vascular smooth muscle cells by bilirubin. J. Biol. Chem. 2012, 287, 15418–15426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Chrifi, I.; Xu, Y.; Pernow, J.; Duncker, D.J.; Merkus, D.; Cheng, C. Uridine adenosine tetraphosphate acts as a proangiogenic factor in vitro through purinergic P2Y receptors. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Velasco, M.A.; Narvaez-Tovar, C.A.; Garzon-Alvarado, D.A. Design, materials, and mechanobiology of biodegradable scaffolds for bone tissue engineering. Biomed. Res. Int. 2015, 729076, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Leiendecker, A.; Witzleben, S.; Schulze, M.; Tobiasch, E. Template-mediated Biomineralization for Bone Tissue Engineering. Curr. Stem Cell Res. Ther. 2017, 12, 103–123. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.G., Jr. 3D Scaffold Fabrication Approaches. Available online: https://www.nist.gov/sites/default/files/images/mml/bbd/biomaterials/scaffoldomics.jpg. (accessed on 29 August 2018).
- Costa, P.F.; Puga, A.M.; Diaz-Gomez, L.; Concheiro, A.; Busch, D.H.; Alvarez-Lorenzo, C. Additive manufacturing of scaffolds with dexamethasone controlled release for enhanced bone regeneration. Int. J. Pharm. 2015, 496, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, D.; Yoon, T.R.; Kim, H.K.; Jo, H.H.; Park, J.S.; Lee, J.H.; Kim, W.D.; Kwon, I.K.; Park, S.A. Surface modification of 3D-printed porous scaffolds via mussel-inspired polydopamine and effective immobilization of rhBMP-2 to promote osteogenic differentiation for bone tissue engineering. Acta Biomater. 2016, 40, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, L.; Zhai, D.; Shi, M.; Luo, Y.; Feng, C.; Fang, B.; Yin, J.; Chang, J.; Wu, C. Mesoporous bioactive glass nanolayer-functionalized 3D-printed scaffolds for accelerating osteogenesis and angiogenesis. Nanoscale 2015, 7, 19207–19221. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Tarafder, S.; Bandyopadhyay, A. Effect of Chemistry on Osteogenesis and Angiogenesis Towards Bone Tissue Engineering Using 3D Printed Scaffolds. Ann. Biomed. Eng. 2016, 45, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Kolan, K.; Li, W.; Semon, J.; Day, D.; Leu, M. 3D bioprinting of stem cells and polymer/bioactive glass composite scaffolds for bone tissue engineering. Int. J. Bioprinting 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Castilho, M.; Rodrigues, J.; Pires, I.; Gouveia, B.; Pereira, M.; Moseke, C.; Groll, J.; Ewald, A.; Vorndran, E. Fabrication of individual alginate-TCP scaffolds for bone tissue engineering by means of powder printing. Biofabrication 2015, 7, 015004. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, S.J.; Jo, H.H.; Lee, J.H.; Kim, W.D.; Lee, J.Y.; Su, A. Fabrication and characterization of 3D-printed bone-like β-tricalcium phosphate/polycaprolactone scaffolds for dental tissue engineering. J. Ind. Eng. 2017, 46, 175–181. [Google Scholar] [CrossRef]
- Martínez-Vázquez, F.J.; Cabañas, M.V.; Paris, J.L.; Lozano, D.; Vallet-Regí, M. Fabrication of novel Si-doped hydroxyapatite/gelatine scaffolds by rapid prototyping for drug delivery and bone regeneration. Acta Biomater. 2015, 15, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grémare, A.; Guduric, V.; Bareille, R.; Heroguez, V.; Latour, S.; L’heureux, N.; Fricain, J.C.; Catros, S.; Le Nihouannen, D. Characterization of printed PLA scaffolds for bone tissue engineering. J. Biomed. Mater. Res. 2018, 106, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, Y.; Qin, X.; Wa, Q. 3D printing of concentrated alginate/gelatin scaffolds with homogeneous nano apatite coating for bone tissue engineering. Mater. Des. 2018, 146, 12–19. [Google Scholar] [CrossRef]
- Rajzer, I.; Menaszek, E.; Castano, O. Electrospun polymer scaffolds modified with drugs for tissue engineering. Mater. Sci. Eng. C 2017, 77, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Miszuk, J.M.; Zhao, Y.; Sun, H.; Fong, H. Electrospun Polycaprolactone 3D Nanofibrous Scaffold with Interconnected and Hierarchically Structured Pores for Bone Tissue Engineering. Adv. Healthc. Mater. 2015, 4, 2238–2246. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Cosme, J.G.; Xu, T.; Miszuk, J.M.; Picciani, P.H.; Fong, H.; Sun, H. Three dimensional electrospun PCL/PLA blend nanofibrous scaffolds with significantly improved stem cells osteogenic differentiation and cranial bone formation. Biomaterials 2017, 115, 115–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhand, C.; Ong, S.T.; Dwivedi, N.; Diaz, S.M.; Venugopal, J.R.; Navaneethan, B.; Fazil, M.H.; Liu, S.; Seitz, V.; Wintermantel, E.; et al. Bio-inspired in situ crosslinking and mineralization of electrospun collagen scaffolds for bone tissue engineering. Biomaterials 2016, 104, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Song, Y.; Zhang, J.; Liu, W.; Cui, J.; Li, H.; Chen, F. Laminated electrospun nHA/PHB-composite scaffolds mimicking bone extracellular matrix for bone tissue engineering. Mater. Sci. Eng. C 2017, 72, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Samanta, A.; Srivastava, R.K.; Hakkarainen, M. Starch-Derived Nanographene Oxide Paves the Way for Electrospinnable and Bioactive Starch Scaffolds for Bone Tissue Engineering. Biomacromolecules 2017, 18, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; He, J.; Sang, F.; Ding, B.; Chen, L.; Cui, S.; Li, K.; Han, Q.; Tan, W. Coaxial electrospun aligned tussah silk fibroin nanostructured fiber scaffolds embedded with hydroxyapatite-tussah silk fibroin nanoparticles for bone tissue engineering. Mater. Sci. Eng. C 2016, 58, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.H.; Ahn, S.H.; Yang, G.-H.; Kim, G.H. A MSCs-laden polycaprolactone/collagen scaffold for bone tissue regeneration. RSC Adv. 2016, 6, 6259–6265. [Google Scholar] [CrossRef]
- Cui, L.; Zhang, N.; Cui, W.; Zhang, P.; Chen, X. A Novel Nano/Micro-Fibrous Scaffold by Melt-Spinning Method for Bone Tissue Engineering. J. Bionic. Eng. 2015, 12, 117–128. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, L.; Fan, D.; Xue, W.; Zhu, C.; Li, X. Physicochemical properties and biological behavior of injectable crosslinked hydrogels composed of pullulan and recombinant human-like collagen. J. Mater. Sci. 2017, 52, 3771–3785. [Google Scholar] [CrossRef]
- Nguyen, B.-N.B.; Moriarty, R.A.; Kamalitdinov, T.; Etheridge, J.M.; Fisher, J.P. Collagen Hydrogel Scaffold Promotes Mesenchymal Stem Cell and Endothelial Cell Coculture for Bone Tissue Engineering. J. Biomed. Mater. Res. 2017, 105, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.N.; Tatara, A.M.; Santoro, M.; van den Beucken, J.J.; Leeuwenburgh, S.C.; Jansen, J.A.; Mikos, A.G. Acellular mineral deposition within injectable, dual-gelling hydrogels for bone tissue engineering. J. Biomed. Mater. Res. 2017, 105, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Nourmohammadi, J.; Shahriarpanah, S.; Asadzadehzanjani, N.; Khaleghpanah, S.; Heidari, S. Biomimetic apatite layer formation on a novel citrate starch scaffold suitable for bone tissue engineering applications. Starch Staerke 2016, 68, 1275–1281. [Google Scholar] [CrossRef]
- Paris, J.L.; Román, J.; Manzano, M.; Cabañas, M.V.; Vallet-Regí, M. Tuning dual-drug release from composite scaffolds for bone regeneration. Int. J. Pharm. 2015, 486(1–2), 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Zhu, Y.; Tong, H.; Shen, X.; Chen, L.; Ran, J. A detailed study of homogeneous agarose/hydroxyapatite nanocomposites for load-bearing bone tissue. Int. J. Biol. Macromol. 2016, 82, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kawazoe, N.; Chen, G. Preparation of dexamethasone-loaded biphasic calcium phosphate nanoparticles/collagen porous composite scaffolds for bone tissue engineering. Acta Biomater. 2017, 67, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Iwai, S.; Shimizu, H.; Suzawa, Y.; Akashi, M.; Yura, Y. Hydroxyapatite agarose composite gels as a biochemical material for the repair of alveolar bone defects due to cleft lip and palate. J. Oral Maxillofac. Surgery Med. Pathol. 2014, 27, 637–644. [Google Scholar] [CrossRef]
- Kolanthai, E.; Dikeshwar Colon, V.S.; Sindu, P.A.; Sarath Chandra, V.; Karthikeyan, K.R.; Surendar Babu, M.; Meenakshi Sundaram, S.; Palanichamy, M.; Narayana Kalkura, S. Effect of solvent; enhancing the wettability and engineering the porous structure of a calcium phosphate/agarose composite for drug delivery. RSC Adv. 2015, 5, 18301–18311. [Google Scholar] [CrossRef]
- Kolanthai, E.; Ganesan, K.; Epple, M.; Kalkura, S.N. Synthesis of nanosized hydroxyapatite/agarose powders for bone filler and drug delivery application. Mater. Today Commun. 2016, 8, 31–40. [Google Scholar] [CrossRef]
- Montesi, M.; Panseri, S.; Iafisco, M.; Adamiano, A.; Tampieri, A. Effect of hydroxyapatite nanocrystals functionalized with lactoferrin in osteogenic differentiation of mesenchymal stem cells. J. Biomed. Mater. Res. A 2015, 103, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Abert, J.; Amella, A.; Weigelt, S.; Fischer, H. Degradation and swelling issues of poly-(d,l-lactide)/??-tricalcium phosphate/calcium carbonate composites for bone replacement. J. Mech. Behav. Biomed. Mater. 2016, 54, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Jiao, K.; Niu, L.N.; Li, Q.H.; Chen, F.M.; Zhao, W.; Li, J.J.; Chen, J.H.; Cutler, C.W.; Pashley, D.H.; Tay, F.R. Biphasic silica/apatite co-mineralized collagen scaffolds stimulate osteogenesis and inhibit RANKL-mediated osteoclastogenesis. Acta Biomater. 2015, 19, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thadavirul, N.; Pavasant, P.; Supaphol, P. Fabrication and Evaluation of Polycaprolactone-Poly(hydroxybutyrate) or Poly(3-Hydroxybutyrate- co -3-Hydroxyvalerate) Dual-Leached Porous Scaffolds for Bone Tissue Engineering Applications. Macromol. Mater. Eng. 2016, 302, 1600289. [Google Scholar] [CrossRef]
- Poh, P.S.P.; Hutmacher, D.W.; Holzapfel, B.M.; Solanki, A.K.; Stevens, M.M.; Woodruff, M.A. In vitro and in vivo bone formation potential of surface calcium phosphate-coated polycaprolactone and polycaprolactone/bioactive glass composite scaffolds. Acta Biomater. 2016, 30, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Tamburaci, S.; Tihminlioglu, F. Biosilica incorporated 3D porous scaffolds for bone tissue engineering applications. Mater. Sci. Eng. C 2018, 91, 274–291. [Google Scholar] [CrossRef] [PubMed]
- Brancato, D.H. Method of Using Silicon Substituted Phosphates to Improve Healing of Bone and Soft Tissue. US20150093449A1, 2 April 2015. [Google Scholar]
- Schulz, O.; Howe, J.; Swaim, R.; Huber, B.; Batts, J.; Harness, D.; Belaney, R. (Agnovos Healthcare, LLC). Composition comprising calcium phosphate and sulfate powders and tricalcium phosphate particles used in the treatment of degenerative bone conditions. EP2987507A1, 24 February 2016. [Google Scholar]
- Calabro, A.; Akst, L.; Alam, D.; Chan, J.; Darr, A.B.; Fukamachi, K.; Gross, R.A.; Haynes, D.; Kamohara, K.; Knott, D.P.; et al. (The Cleveland Clinic Foundation) Hydroxyphenyl crosslinked macromolecular network. EP1773943B1, 9 March 2016. [Google Scholar]
- Cooper-White, J.J.; Cao, Y.; Rowlands, A.S. (The University of Queensland Australia) Production of Porous Polymer Blend Structures for Scaffold. EP2010597B1, 17 June 2015. [Google Scholar]
- Das, A.; Basak, P.; Geesala, R.; Bar, N.; Dhoke, N.R.; Kaushik, K. (Council of Scientific & Industrial Research) Porous Polymer Scaffold Useful for Tissue Engineering in Stem Cell Transplantation. US2017112960A1, 27 April 2017. [Google Scholar]
- Saltzman, W.M.; Fahmy, T.; Fong, P. (Yale University) Targeted and High Density Drug Loaded Polymeric Materials. EP1768692B1, 25 March 2015. [Google Scholar]
- McKay, W.F. (Warsaw Orthopedic Inc.) Medical Implant Sheets Useful for Tissue Regeneration Comprising Calcium-Containing Osteoconductive Material. EP2428233B1, 7 October 2015. [Google Scholar]
- Müller, W.E.L.G.; Schröder, H.-C.W.F.; Wang, X. Printable Morphogenetic Phase-Specific Chitosan-Calcium-Polyphosphate Scaffold For Bone Repair. WO2016012583A1, 28 January 2016. [Google Scholar]
- Kipper, M.J.; Ehrhardt, N.P.; Romero, R.; Gonzales, T.R. (Colorado State University Research Foundation) Methods for Coating Bone Allografts with Periosteum-Mimetic Tissue Engineering Scaffolds. WO2015048322A1, 2 April 2015. [Google Scholar]
- Kumta, P.N.; Yu, G.; Roy, A. (University of Pittsburgh) Biomimetic Plywood Motifs for Bone Tissue Engineering. WO2017201371A1, 23 November 2017. [Google Scholar]
- Zhang, Z.; Wang, Z.; Wu, D.; Zhang, Z. (Shanghai Ninth People’s Hospital; Shanghai Jiao Tong University School Of Medicine) Multi-Step Method For Fabricating Tissue Engineering Bone. US20160058911A1, 3 March 2016. [Google Scholar]
- Tayebi, L.; Rasoulianboroujeni, M. (Marquette University) Reinforced Bone Scaffold. WO2017192525A1, 9 November 2017. [Google Scholar]
- Kutkut, A. Assessment of the Quality and Quantity of Bone Regeneration. University of Kentucky, January 31, 2018. Available online: https://clinicaltrials.gov/ct2/show/NCT03417375 (accessed on 31 August 2018).
- Al-Adwan, S. Use of Stem Cells Cultured on a Scaffold for the Treatment of Aneurysmal Bone Cysts (ABC), University of Jordan, February 28, 2017. Available online: https://clinicaltrials.gov/ct2/show/NCT03066245 (accessed on 31 August 2018).
- Rambam Health Care Campus, Lateral Ridge Augmentation Using a Volume Maintaining Collagen Scaffold Versus Allograft With Collagen Membrane. Rambam Health Care Campus, March 7, 2018. Available online: https://clinicaltrials.gov/ct2/show/NCT03457298 (accessed on 31 August 2018).
- Braddock, D.; Albright, R. (Yale University) Compositions and Methods for Treating Pathological Calcification and Ossification. US20170252414, 7 September 2018. [Google Scholar]
- Debien, L.P.P.; Jaen, J.C.; Kalisiak, J.; Lawson, K.; Leleti, M.R.; Lindsey, E.A.; Miles, D.H.; Newcomb, E.; Powers, J.P.; Rosen, B.R.; et al. (Arcus Biosciences Inc.) Modulators of 5’-Nucleotidase, Ecto and the Use Thereof. 20170267710, 21 September 2017. [Google Scholar]
- Billedeau, R.J.; Li, J.; Chen, L. (Calithera Biosciences Inc.) Ectonucleotidase Inhibitors and Methods of Use Thereof. US2018186827A1, 5 July 2018. [Google Scholar]
- Mueller, C.E.; Brunschweiger, A.; Iqbal, J. (University Bonn) Ectonucleotidase Inhibitors. US2010204182A1, 12 August 2010. [Google Scholar]
- Chanteux, S.; Paturel, C.; Perrot, I.; Gauthier, L. (Innate Pharma) CD73 Blockade. 20180030144, 2 February 2018. [Google Scholar]
- Bensussan, A.; Bonnefoyberard, N.; Eliaou, N.; Alberici, G.; Bastid, J. (Orega Biotech; INSERM—Institut National de la Santé et de la Recherche Médicale). Antibodies Against Human CD39 and Use Thereof. EP2654789, 30 October 2013. [Google Scholar]
- Jaschinski, F.; Hilmenyuk, T. (Secarna Pharmaceuticals GmbH & Co KG) Immunosuppression-Reverting Oligonucleotides Inhibiting the Expression of CD39. WO2018065622A1, 12 April 2018. [Google Scholar]
- Beatty, J.; Debien, L.; Jeffrey, J.; Leleti, M.R.; Mandal, D.; Miles, D.; Powers, J.; Rosen, B.; Sharif, E.; Thomastran, R. (Arcus Biosciences Inc.) Azolopyrimidine for the Treatment of Cancer-related Disorders. 20180215730, 2 August 2018. [Google Scholar]
- Cronstein, B.M.; Medierto, M.A. (University of New York) Methods and Compositions for Stimulating Bone Regeneration. US2018055868A1, 1 March 2018. [Google Scholar]
- Zheng, J.; Won, W.; Berlin, M.; Ting, P.; Li, G.; Xiao, D.; Wang, H.; Aslanian, R. (Merck Sharp & Dohme) Aminopyrazine Compounds with A2A Antagonist Properties. US2018193314A1, 12 July 2018. [Google Scholar]
- Lu, B.; Zhang, J.; Jin, F.; He, F.; Tao, W. (Jiangsu Hengrui Medicine Co; Shanghai Hengrui Pharmaceutical Co Ltd.) 1,2,4-Triazine-3-amine Derivative, Preparation Method Therefor, and Use Thereof in Medicine. WO2018130184A1, 19 July 2018. [Google Scholar]
- Sato, N.; Yuki, Y.; Shinohara, H.; Takeji, Y.; Ito, K.; Michikami, D.; Hino, K.; Yamazaki, H. (Otsuka Pharma Co Ltd.) A Novel Cyanopyrimidine Derivative. MY165572A, 5 April 2018. [Google Scholar]
- Lin, Y.-L.; Chern, Y.; Fang, J.-M.; Lin, J.-H.; Huang, N.-K. (Academia Sinica) Dual-Action Compounds Targeting Adenosine A2A Receptor and Adenosine Transporter for Prevention and Treatment of Neurodegenerative Diseases. HUE035188T2, 2 May 2018. [Google Scholar]
- Zablocki, J.; Elzein, E.; Seemayer, R.; Lemons, T. (Gilead Sciences Inc.) Process for Preparing an A2A-Adenosine Receptor Agonists and Ist Polymorphs A2A. HK1127358A1, 5 January 2018. [Google Scholar]
- Ali, A.; Kuang, R.; Lim, Y.H.; Man-chu Lo, M.; Ting, P.C.; Biju, P.; de Lera Ruiz, M.; Degrado, S.J.; Tung, A.L.; Henderson, T.J.; et al. (Merck Sharp & Dome) Aminoquinazoline Compounds as A2A Antagonist. US2018037554A1, 8 February 2018. [Google Scholar]
- Camacho-Gómez, J.A.; Castro-Palomino, L.J.C. (Palobiofarma SL) 4-Aminopyrimidine Derivatives and Their As as Adenosine A2A Receptor Antagonist. ME02100B, 20 October 2015. [Google Scholar]
- Fairhurst, R.A.; Taylor, R.J. (Novartis AG) Purine Derivatives as A2A Agonists. TNSN08544A1, 14 April 2010. [Google Scholar]
- Harter, M.; Kosemund, D.; Delbeck, M.; Kalthof, B.; Wasnaire, P.; Sussmeier, F.; Lustig, K. (Bayer Pharma AG) Heterocyclymethyl-thienouracile as Antagonist of Adenosine –A2B-Receptor. PH12017501758A1, 11 April 2018. [Google Scholar]
- Mendoza Lizaldez, A.; Camacho Gómez, J.A.; Castro Palomino, L.J.C. (Palobiofarma SL)Derivatives of 2-Aminopyridine as Adenosine A2B Receptor Antagonists and Ligands of the Melatonin MT3 Receptors. MX2017010949A, 15 December 2017. [Google Scholar]
- Thompson, R.; Beauglehole, A.; Wang, Q. (Lewis and Clark Pharmaceuticals Inc.) Xanthine-substituted Alkynyl Carbamates/Reverse Carbamates as A2B Antagonists. MX2017012783, 30 January 2018. [Google Scholar]
- Barawkar, D.; Basu, S.; Ramdas, V.; Palle, V.P.; Waman, Y.; Patel, M.; Panmand, A. (Advinus Therapeutics Private Ltd.) Purine Compounds as Prodrugs of A2B Adenosine Receptor Antagonists, Their Process and Medicinal Applications. CN103261200A, 21 August 2013. [Google Scholar]
- Jacobson, K.A.; Tosh, D.K.; Salvemini, D. (Saint Louis University) A3 Adenosine Receptor Agonists. The US Secretary Department of Health and Human Service. US2018230150A1, 16 August 2018. [Google Scholar]
- Castro Palomino Laria, J.; Camacho Goméz, J.; El Maatougui, A. (Palobiofarma SL) Adenosine A3 Receptor Modulators. WO2018134464A1, 26 July 2018. [Google Scholar]
- Lin, J.; Wei, Y.; Zhao, X. (University Nankai) Small-Molecular Antagonist for A3 Adenosine Receptor. CN108017584A, 11 May 2018. [Google Scholar]
- Lee, J.; Kim, S.; Kim, D.; Ahn, K.H.; Lee, G.B.; Kim, D.; Hwang, H.S. (Handoc Inc.) Compounds Antagonizing A3 Adenosine Receptor, Method for Preparing Them, and Medical-use Thereof. CA3011442A1, 20 July 2017. [Google Scholar]
- Finet, M.; Urbán, K.; Szabó, K.; Szabó, T.; Susán, E.; Boer, K.; Kapui, Z.; Tímári, G.; Bátori, S.; Szlávik, Z.; et al. (Sanofi Aventis) Triazolo [1,5-A] Quinolines as Adenosine A3 Receptor Ligands. ME00958B, 20 June 2012. [Google Scholar]
- Lee, P.P.; Draganov, D. (City of Hope). Macrocyclic Lactones and Uses Thereof as Modulators of Purinergic Receptors. 20170119807, 4 May 2017. [Google Scholar]
- Vassilevski, A.A.; Oparin, P.B.; Korolkova, Y.V.; Mosharova, I.V.; Savchenko, G.A.; Boychuk, Y.A.; Krishtal, O.A. (Future Analgesics Ltd.) Peptide Modulator of Purinoceptors. WO2018106142A1, 14 June 2018. [Google Scholar]
- Thompson, S.K.; Venkatesan, A.; Priestley, T.; Kundu, M.; Saha, A. (Asana Biosciences, LLC) P2X3 and /or P2X2/3 Compounds and Methods. 20180186793, 5 July 2018. [Google Scholar]
- Chen, L.; Dillon, M.P.; Feng, L.; Hawley, R.C.; Yang, M. (Roche Palo Alto, LLC) Thiazole and Oxazole Substituted Arylamides as P2X3 and P2X2/3 Antagonists. 20170182057, 29 June 2017. [Google Scholar]
- Darwish, M.; Kaur, S.; Su, D.; Xu, K. (Genentech, Inc.) Bioanalytical Analysis of Site-specific Antibody Drug Conjugates. 20170370906, 12 December 2017. [Google Scholar]
- Barden, J.A.; Brewis, N.; Jones, P.; Grant, S. (Biosceptre (Aust) Pty Ltd.) Anti P2X7 Receptor Antibodies and Fragments Thereof. 10053508, 16 November 2018. [Google Scholar]
- Kilburn, J.P.; Rasmussen, L.K.; Jessing, M.; Eldemenky, E.M.; Chen, B.; Jiang, Y.; Hopper, A.T. (Lundbeck & Co As H) Benzamides. NZ630830A, 27 November 2016. [Google Scholar]
- Cohen, D.; Chumakov, I.; Nabirochkin, S.; Guerassimenko, O.; Graudens, E. (Pharnext) New Therapeutic Approaches for Treating Alzheimer Disease and Related Disorders Through a Modulation of Angiogenesis. EP2282778, 16 February 2011. [Google Scholar]
- Goldman, A.L.; Donlon, C.M.; Cook, N.R.; Manson, J.E.; Buring, J.E.; Copeland, T.; Yu, C.Y.; LeBoff, M.S. VITamin D and OmegA-3 TriaL (VITAL) bone health ancillary study: Clinical factors associated with trabecular bone score in women and men. Osteoporos Int. 2018, 29, 2505–2515. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Walsh, M.C.; Takegahara, N.; Middleton, S.A.; Shin, H.I.; Kim, J.; Choi, Y. The purinergic receptor P2X5 regulates inflammasome activity and hyper-multinucleation of murine osteoclasts. Sci. Rep. 2017, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kajikawa, T.; Walsh, M.C.; Takegahara, N.; Jeong, Y.H.; Hajishengallis, G.; Choi, Y. The purinergic receptor P2X5 contributes to bone loss in experimental periodontitis. BMB Rep. 2018, 51, 468–473. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ottensmeyer, P.F.; Witzler, M.; Schulze, M.; Tobiasch, E. Small Molecules Enhance Scaffold-Based Bone Grafts via Purinergic Receptor Signaling in Stem Cells. Int. J. Mol. Sci. 2018, 19, 3601. https://doi.org/10.3390/ijms19113601
Ottensmeyer PF, Witzler M, Schulze M, Tobiasch E. Small Molecules Enhance Scaffold-Based Bone Grafts via Purinergic Receptor Signaling in Stem Cells. International Journal of Molecular Sciences. 2018; 19(11):3601. https://doi.org/10.3390/ijms19113601
Chicago/Turabian StyleOttensmeyer, Patrick Frank, Markus Witzler, Margit Schulze, and Edda Tobiasch. 2018. "Small Molecules Enhance Scaffold-Based Bone Grafts via Purinergic Receptor Signaling in Stem Cells" International Journal of Molecular Sciences 19, no. 11: 3601. https://doi.org/10.3390/ijms19113601
APA StyleOttensmeyer, P. F., Witzler, M., Schulze, M., & Tobiasch, E. (2018). Small Molecules Enhance Scaffold-Based Bone Grafts via Purinergic Receptor Signaling in Stem Cells. International Journal of Molecular Sciences, 19(11), 3601. https://doi.org/10.3390/ijms19113601