TUBB3 Reverses Resistance to Docetaxel and Cabazitaxel in Prostate Cancer



 ,
,
Abstract
:1. Introduction
2. Results
2.1. Characterization of DTX- and CBZ-Resistant PCa Cell Lines
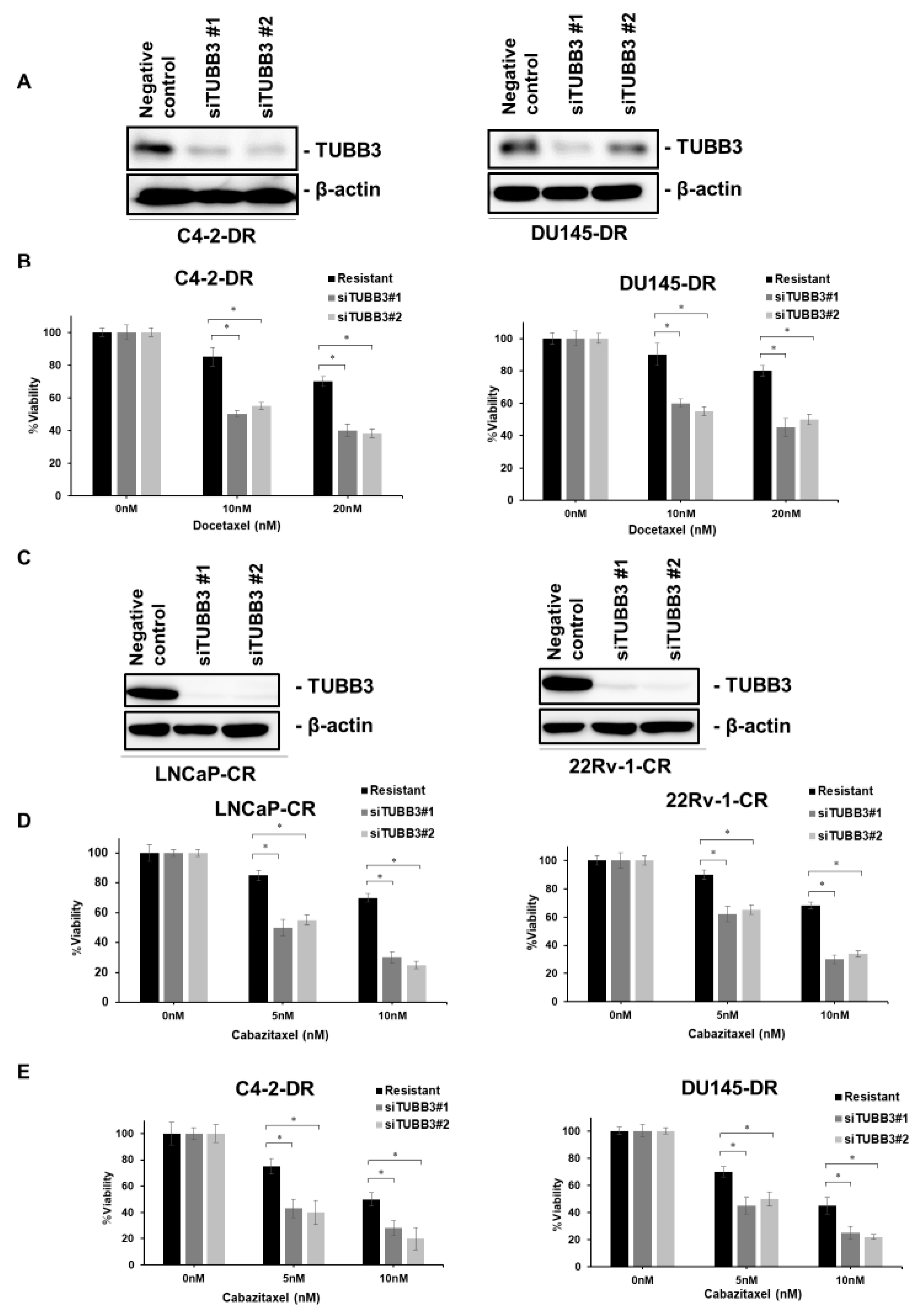
2.2. TUBB3 is Overexpressed in DTX- and CBZ-Resistant PCa Cell Lines
2.3. Inhibition of TUBB3 Reverses DTX and CBZ Resistance
2.4. Interaction Between PTEN and TUBB3 Expression
2.5. LY294002 Improves Taxane Resistance
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. DTX, CBZ, and LY294002 Treatment
4.3. Western Blotting Analysis
4.4. qRT-PCR Analysis
4.5. Tissue Samples
4.6. RNA Interference
4.7. Generation of PTEN Knockout Cells
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CBZ | cabazitaxel |
| c-PARP | cleaved PARP |
| CRPC | castration-resistant prostate cancer |
| DTX | docetaxel |
| MTT | 4,5-dimethylthiazol-2-yl-2,5-diphenyltetrazolium bromide |
| PCa | prostate cancer |
| qRT-PCR | quantitative reverse transcriptase polymerase chain reaction |
| SiRNA | small interfering RNA |
References
- Van Neste, L.; Herman, J.G.; Otto, G.; Bigley, J.W.; Epstein, J.I.; Van Criekinge, W. The epigenetic promise for prostate cancer diagnosis. Prostate 2012, 72, 1248–1261. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, A.J.; Kattan, M.W.; Eastham, J.A.; Dotan, Z.A.; Bianco, F.J., Jr.; Lilja, H.; Scardino, P.T. Defining biochemical recurrence of prostate cancer after radical prostatectomy: A proposal for a standardized definition. J. Clin. Oncol. 2006, 24, 3973–3978. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.; Belldegrun, A. Novel therapeutic strategies for castration resistant prostate cancer: Inhibition of persistent androgen production and androgen receptor mediated signaling. J. Urol. 2011, 185, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Petrylak, D.P.; Tangen, C.M.; Hussain, M.H.; Lara, P.N., Jr.; Jones, J.A.; Taplin, M.E.; Burch, P.A.; Berry, D.; Moinpour, C.; Kohli, M.; et al. Docetaxel and estramustine compared with mitoxantrone and prednisone for advanced refractory prostate cancer. N. Engl. J. Med. 2004, 351, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- De Bono, J.S.; Oudard, S.; Ozguroglu, M.; Hansen, S.; Machiels, J.P.; Kocak, I.; Gravis, G.; Bodrogi, I.; Mackenzie, M.J.; Shen, L.; et al. TROPIC Investigators. Prednisone plus cabazitaxel or mitoxantrone for metastatic castration-resistant prostate cancer progressing after docetaxel treatment: A randomised open-label trial. Lancet 2010, 376, 1147–1154. [Google Scholar] [CrossRef]
- Lombard, A.P.; Liu, C.; Armstrong, C.M.; Cucchiara, V.; Gu, X.; Lou, W.; Evans, C.P.; Gao, A.C. ABCB1 mediates cabazitaxel-docetaxel cross-resistance in advanced prostate cancer. Mol. Cancer Ther. 2017, 16, 2257–2266. [Google Scholar] [CrossRef]
- Lombard, A.P.; Liu, L.; Cucchiara, V.; Liu, C.; Armstrong, C.M.; Zhao, R.; Yang, J.C.; Lou, W.; Evans, C.P.; Gao, A.C. Intra versus inter cross-resistance determines treatment sequence between taxane and AR-targeting therapies in advanced prostate cancer. Mol. Cancer Ther. 2018, 17, 2197–2205. [Google Scholar]
- Dumontet, C.; Sikic, B.I. Mechanisms of action of and resistance to antitubulin agents: Microtubule dynamics, drug transport, and cell death. J. Clin. Oncol. 1999, 17, 1061–1070. [Google Scholar] [CrossRef]
- Katsetos, C.D.; Herman, M.M.; Mork, S.J. Class III beta-tubulin in human development and cancer. Cell Motil. Cytoskelet. 2003, 55, 77–96. [Google Scholar] [CrossRef]
- Terry, S.; Ploussard, G.; Allory, Y.; Nicolaiew, N.; Boissière-Michot, F.; Maillé, P.; Kheuang, L.; Coppolani, E.; Ali, A.; Bibeau, F.; et al. Increased expression of class III beta-tubulin in castration-resistant human prostate cancer. Br. J. Cancer 2009, 101, 951–956. [Google Scholar] [CrossRef]
- Tame, M.A.; Manjon, A.G.; Belokhvostova, D.; Raaijmakers, J.A.; Medema, R.H. TUBB3 overexpression has a negligible effect on the sensitivity to taxol in cultured cell lines. Oncotarget 2017, 8, 71536–71547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravina, G.L.; Mancini, A.; Colapietro, A.; Marampon, F.; Sferra, R.; Pompili, S.; Biordi, L.A.; Iorio, R.; Flati, V.; Argueta, C.; et al. Pharmacological treatment with inhibitors of nuclear export enhances the antitumor activity of docetaxel in human prostate cancer. Oncotarget 2017, 8, 111225–111245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.E.; Chen, H.; Yuan, S.F.; Wu, L.L.; Zhang, W.; Sun, H.Y.; Chen, W.J. Overexpression of betaIII-tubulin and survivin associated with drug resistance to docetaxel-based chemotherapy in advanced gastric cancer. J. BUON 2012, 17, 284–290. [Google Scholar]
- Ploussard, G.; Terry, S.; Maillé, P.; Allory, Y.; Sirab, N.; Kheuang, L.; Soyeux, P.; Nicolaiew, N.; Coppolani, E.; Paule, B.; et al. Class III beta-tubulin expression predicts prostate tumor aggressiveness and patient response to docetaxel-based chemotherapy. Cancer Res. 2010, 70, 9253–9264. [Google Scholar] [CrossRef] [PubMed]
- Duran, G.E.; Wang, Y.C.; Francisco, E.B.; Rose, J.C.; Martinez, F.J.; Coller, J.; Brassard, D.; Vrignaud, P.; Sikic, B.I. Mechanisms of resistance to cabazitaxel. Mol. Cancer Ther. 2015, 14, 193–201. [Google Scholar] [CrossRef]
- Smiyun, G.; Azarenko, O.; Miller, H.; Rifkind, A.; LaPointe, N.E.; Wilson, L.; Jordan, M.A. betaIII-tubulin enhances efficacy of cabazitaxel as compared with docetaxel. Cancer Chemother. Pharmacol. 2017, 80, 151–164. [Google Scholar] [CrossRef]
- Shiota, M.; Itsumi, M.; Yokomizo, A.; Takeuchi, A.; Imada, K.; Kashiwagi, E.; Inokuchi, J.; Tatsugami, K.; Uchiumi, T.; Naito, S. Targeting ribosomal S6 kinases/Y-box binding protein-1 signaling improves cellular sensitivity to taxane in prostate cancer. Prostate 2014, 74, 829–838. [Google Scholar] [CrossRef]
- Shiota, M.; Kashiwagi, E.; Yokomizo, A.; Takeuchi, A.; Dejima, T.; Song, Y.; Tatsugami, K.; Inokuchi, J.; Uchiumi, T.; Naito, S. Interaction between docetaxel resistance and castration resistance in prostate cancer: Implications of Twist1, YB-1, and androgen receptor. Prostate 2013, 73, 1336–1344. [Google Scholar] [CrossRef]
- Sekino, Y.; Oue, N.; Koike, Y.; Shigematsu, Y.; Sakamoto, N.; Sentani, K.; Teishima, J.; Shiota, M.; Matsubara, A.; Yasui, W. KIFC1 Inhibitor CW069 induces apoptosis and reverses resistance to docetaxel in prostate cancer. J. Clin. Med. 2019, 8, 225. [Google Scholar] [CrossRef]
- Shiota, M.; Dejima, T.; Yamamoto, Y.; Takeuchi, A.; Imada, K.; Kashiwagi, E.; Inokuchi, J.; Tatsugami, K.; Kajioka, S.; Uchiumi, T.; et al. Collateral resistance to taxanes in enzalutamide-resistant prostate cancer through aberrant androgen receptor and its variants. Cancer Sci. 2018, 10, 3224–3234. [Google Scholar] [CrossRef]
- McCarroll, J.A.; Gan, P.P.; Erlich, R.B.; Liu, M.; Dwarte, T.; Sagnella, S.S.; Akerfeldt, M.C.; Yang, L.; Parker, A.L.; Chang, M.H.; et al. TUBB3/betaIII-tubulin acts through the PTEN/AKT signaling axis to promote tumorigenesis and anoikis resistance in non-small cell lung cancer. Cancer Res. 2015, 75, 415–425. [Google Scholar] [CrossRef]
- Tsourlakis, M.C.; Weigand, P.; Grupp, K.; Kluth, M.; Steurer, S.; Schlomm, T.; Graefen, M.; Huland, H.; Salomon, G.; Steuber, T.; et al. betaIII-tubulin overexpression is an independent predictor of prostate cancer progression tightly linked to ERG fusion status and PTEN deletion. Am. J. Pathol. 2014, 184, 609–617. [Google Scholar] [CrossRef]
- Skjoth, I.H.; Issinger, O.G. Profiling of signaling molecules in four different human prostate carcinoma cell lines before and after induction of apoptosis. Int. J. Oncol. 2006, 28, 217–229. [Google Scholar] [CrossRef]
- Mulholland, D.J.; Tran, L.M.; Li, Y.; Cai, H.; Morim, A.; Wang, S.; Plaisier, S.; Garraway, I.P.; Huang, J.; Graeber, T.G.; et al. Cell autonomous role of PTEN in regulating castration-resistant prostate cancer growth. Cancer Cell 2011, 19, 792–804. [Google Scholar] [CrossRef]
- Carver, B.S.; Chapinski, C.; Wongvipat, J.; Hieronymus, H.; Chen, Y.; Chandarlapaty, S.; Arora, V.K.; Le, C.; Koutcher, J.; Scher, H.; et al. Reciprocal feedback regulation of PI3K and androgen receptor signaling in PTEN-deficient prostate cancer. Cancer Cell 2011, 19, 575–586. [Google Scholar] [CrossRef]
- Marques, R.B.; Aghai, A.; de Ridder, C.M.A.; Stuurman, D.; Hoeben, S.; Boer, A.; Ellston, R.P.; Barry, S.T.; Davies, B.R.; Trapman, J.; et al. High efficacy of combination therapy using PI3K/AKT inhibitors with androgen deprivation in prostate cancer preclinical models. Eur. Urol. 2015, 67, 1177–1185. [Google Scholar] [CrossRef]
- Maira, S.M.; Stauffer, F.; Schnell, C.; García-Echeverría, C. PI3K inhibitors for cancer treatment: Where do we stand? Biochem. Soc. Trans. 2009, 37, 265–272. [Google Scholar] [CrossRef]
- Xiao, M.; Tang, Y.; Chen, W.W.; Wang, Y.L.; Yang, L.; Li, X.; Song, G.L.; Kuang, J. Tubb3 regulation by the Erk and Akt signaling pathways: A mechanism involved in the effect of arginine ADP-ribosyltransferase 1 (Art1) on apoptosis of colon carcinoma CT26 cells. Tumour Biol. 2016, 37, 2353–2363. [Google Scholar] [CrossRef]
- Levallet, G.; Bergot, E.; Antoine, M.; Creveuil, C.; Santos, A.O.; Beau-Faller, M.; de Fraipont, F.; Brambilla, E.; Levallet, J.; Morin, F.; et al. Intergroupe Francophone de Cancérologie Thoracique (IFCT). High TUBB3 expression, an independent prognostic marker in patients with early non-small cell lung cancer treated by preoperative chemotherapy, is regulated by K-Ras signaling pathway. Mol. Cancer Ther. 2012, 11, 1203–1213. [Google Scholar] [CrossRef]
- Wang, S.; Wu, J.; Suburu, J.; Gu, Z.; Cai, J.; Axanova, L.S.; Cramer, S.D.; Thomas, M.J.; Perry, D.L.; Edwards, I.J.; et al. Effect of dietary polyunsaturated fatty acids on castration-resistant Pten-null prostate cancer. Carcinogenesis 2012, 33, 404–412. [Google Scholar] [CrossRef]
- Qian, D.Z.; Rademacher, B.L.; Pittsenbarger, J.; Huang, C.Y.; Myrthue, A.; Higano, C.S.; Garzotto, M.; Nelson, P.S.; Beer, T.M. CCL2 is induced by chemotherapy and protects prostate cancer cells from docetaxel-induced cytotoxicity. Prostate 2010, 70, 433–442. [Google Scholar] [CrossRef]
- Vale, C.L.; Burdett, S.; Rydzewska, L.H.M.; Albiges, L.; Clarke, N.W.; Fisher, D.; Fizazi, K.; Gravis, G.; James, N.D.; Mason, M.D.; et al. STOpCaP Steering Group. Addition of docetaxel or bisphosphonates to standard of care in men with localised or metastatic, hormone-sensitive prostate cancer: A systematic review and meta-analyses of aggregate data. Lancet Oncol. 2016, 17, 243–256. [Google Scholar] [CrossRef]
- Sweeney, C.J.; Chen, Y.H.; Carducci, M.; Liu, G.; Jarrard, D.F.; Eisenberger, M.; Wong, Y.N.; Hahn, N.; Kohli, M.; Cooney, M.M.; et al. Chemohormonal therapy in metastatic hormone-sensitive prostate cancer. N. Engl. J. Med. 2015, 373, 737–746. [Google Scholar] [CrossRef]
- Kosaka, T.; Hongo, H.; Miyazaki, Y.; Nishimoto, K.; Miyajima, A.; Oya, M. Reactive oxygen species induction by cabazitaxel through inhibiting Sestrin-3 in castration resistant prostate cancer. Oncotarget 2017, 8, 87675–87683. [Google Scholar] [CrossRef] [Green Version]
- Dillon, L.M.; Miller, T.W. Therapeutic targeting of cancers with loss of PTEN function. Curr. Drug Targets 2014, 15, 65–79. [Google Scholar] [CrossRef]
- Rescigno, P.; Lorente, D.; Dolling, D.; Ferraldeschi, R.; Rodrigues, D.N.; Riisnaes, R.; Miranda, S.; Bianchini, D.; Zafeiriou, Z.; Sideris, S.; et al. Docetaxel treatment in PTEN-and ERG-aberrant metastatic prostate cancers. Eur. Urol. Oncol. 2018, 1, 71–77. [Google Scholar] [CrossRef]
- Kosaka, T.; Miyajima, A.; Shirotake, S.; Kikuchi, E.; Oya, M. Phosphorylated Akt up-regulates angiotensin II type-1 receptor expression in castration resistant prostate cancer. Prostate 2011, 71, 1510–1517. [Google Scholar] [CrossRef]
- Prawettongsopon, C.; Asawakarn, S.; Suthiphongchai, T. Suppression of prometastatic phenotype of highly metastatic androgen-independent rat prostate cancer MLL cell line by PI3K inhibitor LY294002. Oncol. Res. 2009, 17, 301–309. [Google Scholar] [CrossRef]
- Shiota, M.; Ushijima, M.; Imada, K.; Kashiwagi, E.; Takeuchi, A.; Inokuchi, J.; Tatsugami, K.; Kajioka, S.; Eto, M. Cigarette smoking augments androgen receptor activity and promotes resistance to antiandrogen therapy. Prostate 2019, 79, 1147–1155. [Google Scholar] [CrossRef]
- Wang, Y.; Kuramitsu, Y.; Baron, B.; Kitagawa, T.; Tokuda, K.; Akada, J.; Maehara, S.I.; Maehara, Y.; Nakamura, K. PI3K inhibitor LY294002, as opposed to wortmannin, enhances AKT phosphorylation in gemcitabine-resistant pancreatic cancer cells. Int. J. Oncol. 2017, 50, 606–612. [Google Scholar] [CrossRef]
- Weigelt, B.; Downward, J. Genomic determinants of PI3K pathway inhibitor response in cancer. Front. Oncol. 2012, 2, 109. [Google Scholar] [CrossRef]
- Sekino, Y.; Oue, N.; Shigematsu, Y.; Ishikawa, A.; Sakamoto, N.; Sentani, K.; Teishima, J.; Matsubara, A.; Yasui, W. KIFC1 induces resistance to docetaxel and is associated with survival of patients with prostate cancer. Urol. Oncol. 2017, 31.e13–31.e20. [Google Scholar] [CrossRef]
- Sekino, Y.; Sakamoto, N.; Goto, K.; Honma, R.; Shigematsu, Y.; Quoc, T.P.; Sentani, K.; Oue, N.; Teishima, J.; Kawakami, F.; et al. Uc.416 + A promotes epithelial-to-mesenchymal transition through miR-153 in renal cell carcinoma. BMC Cancer 2018, 18, 952. [Google Scholar] [CrossRef]
- Sekino, Y.; Oue, N.; Mukai, S.; Shigematsu, Y.; Goto, K.; Sakamoto, N.; Sentani, K.; Hayashi, T.; Teishima, J.; Matsubara, A.; et al. Protocadherin B9 promotes resistance to bicalutamide and is associated with the survival of prostate cancer patients. Prostate 2019, 79, 234–242. [Google Scholar] [CrossRef]
- Sekino, Y.; Sakamoto, N.; Sentani, K.; Oue, N.; Teishima, J.; Matsubara, A.; Yasui, W. miR-130b promotes sunitinib resistance through regulation of PTEN in renal cell carcinoma. Oncology 2019, 1–9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | ID | Dilution |
|---|---|---|
| Primary antibody | ||
| TUBB3 | MMS-435P | 1:500 |
| PTEN | 138G6 | 1:500 |
| Akt | 9272 | 1:500 |
| p-Akt | 9271 | 1:500 |
| Cleaved PARP | 5625 | 1:500 |
| β-actin | A5441 | 1:10,000 |
| Secondary antibody | ||
| Anti-IgG (H+L chain) (Mouse) pAb-HRP | 330 | 1:500 |
| Anti-IgG (H+L chain) (Rabbit) pAb-HRP | 458 | 1:500 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| TUBB3 | GAGATGGAGTTCACCGAGGC | TCGTCTTCGTACATCTCGCC |
| PTEN | ACCCACCACAGCTAGAACTT | GGGAATAGTTACTCCCTTTTTGTC |
| ACTB | TCACCGAGCGCGGCT | TAATGTCACGCACGATTTCCC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekino, Y.; Han, X.; Kawaguchi, T.; Babasaki, T.; Goto, K.; Inoue, S.; Hayashi, T.; Teishima, J.; Shiota, M.; Yasui, W.; et al. TUBB3 Reverses Resistance to Docetaxel and Cabazitaxel in Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 3936. https://doi.org/10.3390/ijms20163936
Sekino Y, Han X, Kawaguchi T, Babasaki T, Goto K, Inoue S, Hayashi T, Teishima J, Shiota M, Yasui W, et al. TUBB3 Reverses Resistance to Docetaxel and Cabazitaxel in Prostate Cancer. International Journal of Molecular Sciences. 2019; 20(16):3936. https://doi.org/10.3390/ijms20163936
Chicago/Turabian StyleSekino, Yohei, Xiangrui Han, Takafumi Kawaguchi, Takashi Babasaki, Keisuke Goto, Shogo Inoue, Tetsutaro Hayashi, Jun Teishima, Masaki Shiota, Wataru Yasui, and et al. 2019. "TUBB3 Reverses Resistance to Docetaxel and Cabazitaxel in Prostate Cancer" International Journal of Molecular Sciences 20, no. 16: 3936. https://doi.org/10.3390/ijms20163936
APA StyleSekino, Y., Han, X., Kawaguchi, T., Babasaki, T., Goto, K., Inoue, S., Hayashi, T., Teishima, J., Shiota, M., Yasui, W., & Matsubara, A. (2019). TUBB3 Reverses Resistance to Docetaxel and Cabazitaxel in Prostate Cancer. International Journal of Molecular Sciences, 20(16), 3936. https://doi.org/10.3390/ijms20163936