Dynamic mRNA Expression Analysis of the Secondary Palatal Morphogenesis in Miniature Pigs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Analysis of mRNA Sequencing in Different Stages of Sus scrofa Secondary Palate Development
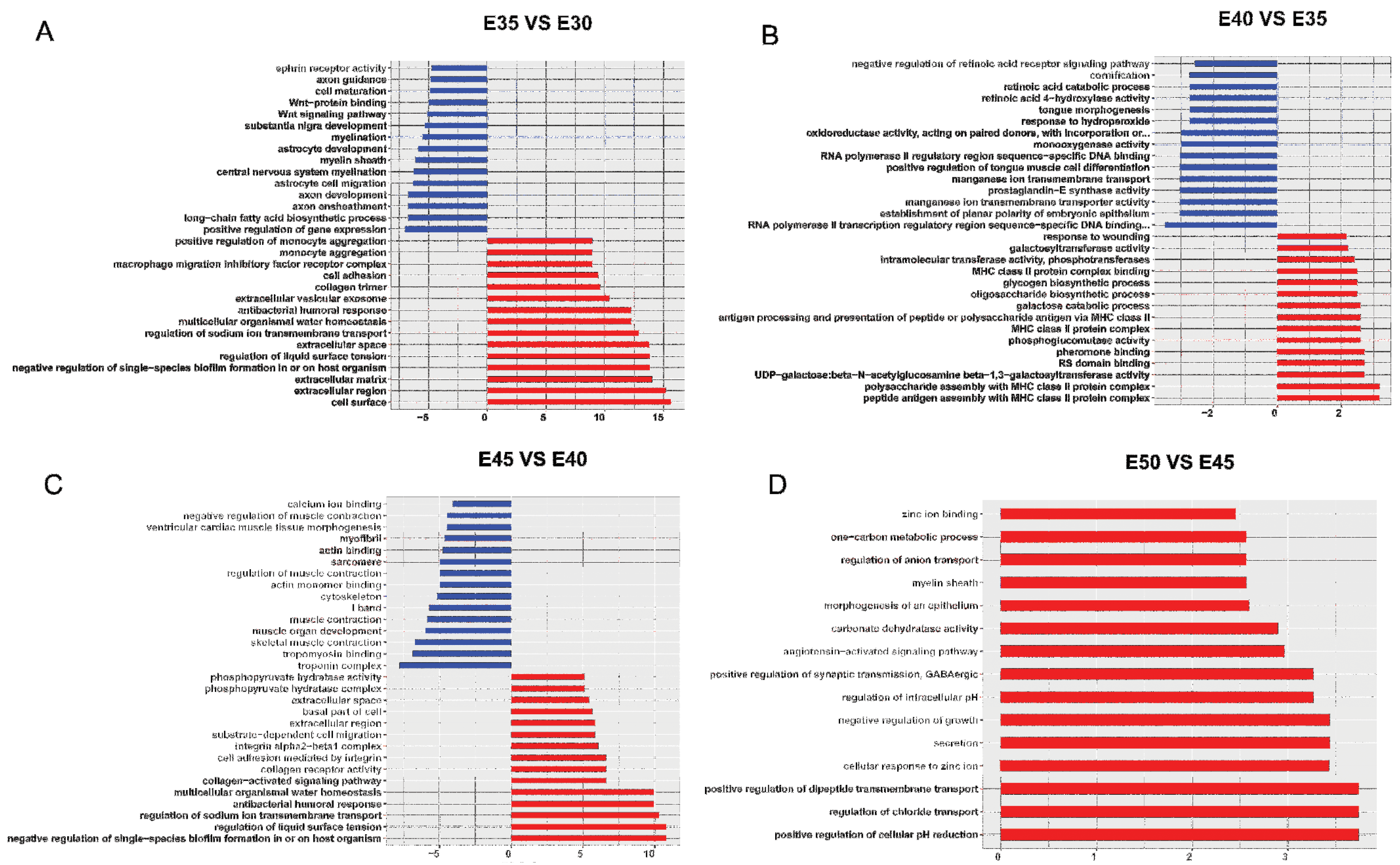
2.2. Gene Ontology Analysis of the Screened Genes in Five Developmental Stages of Palatogenesis
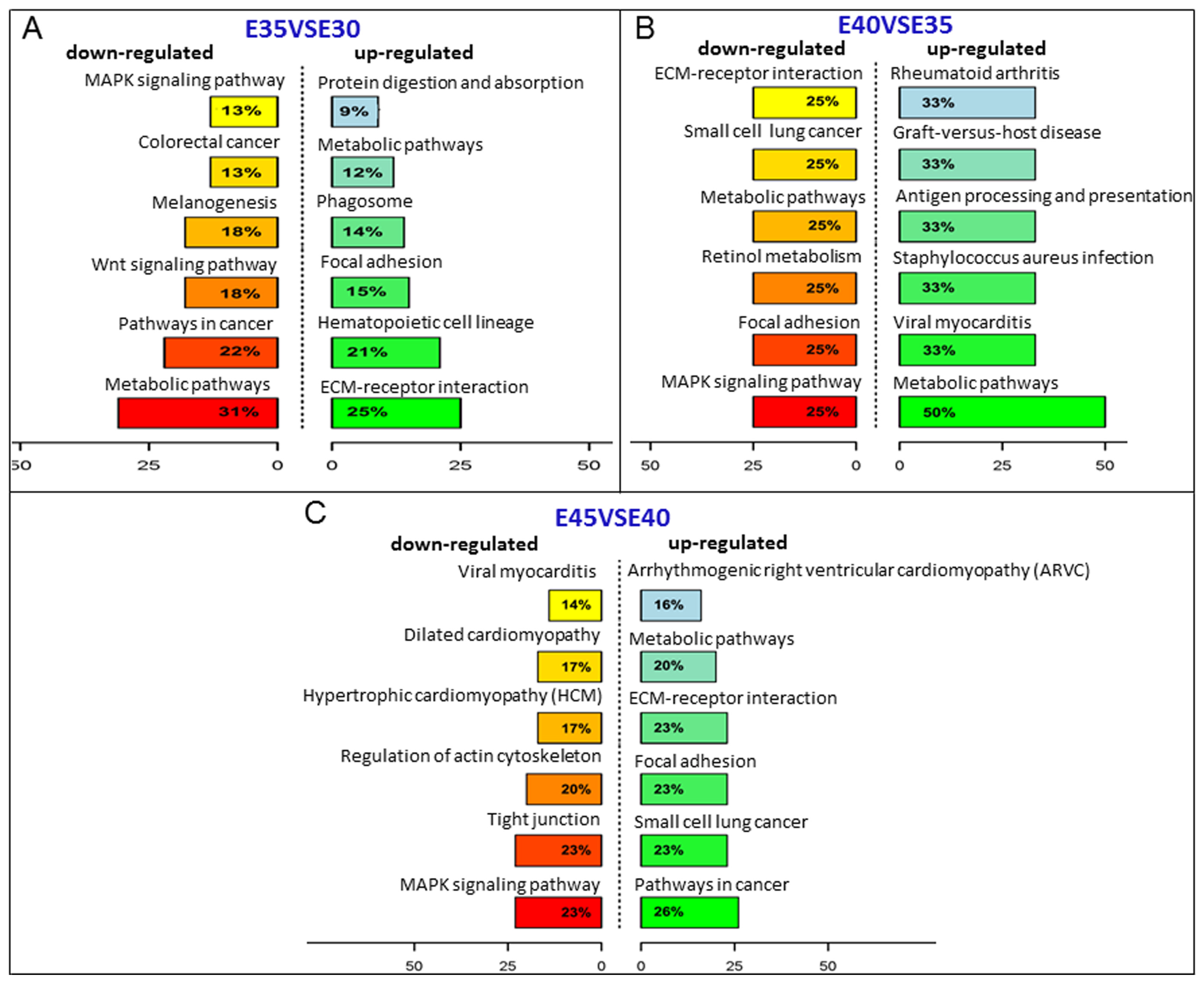
2.3. Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis of the Screened Genes in Five Stages of Palatogenesis
2.4. Quantitative Analysis of mRNA Expression
2.5. The Interaction Network of Differentially Expressed Genes and Pathways (Signal-Gene Net)
3. Discussion
4. Materials and Methods
4.1. Experimental Animals, Tissues Collection and Histological Staining
4.2. Total RNA Isolation, cDNA Libraries Construction and Sequencing
4.3. Gene Expression Profiles of Different Development Stages
4.4. Gene Expression Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CLP | cleft lip/palate |
| E | embryonic days |
| EMT | epithelial-mesenchymal transformation |
| VS | versus |
| MEE | medial edge epithelium |
| DEGs | different expression genes |
| GO | Gene ontology |
| KEGG | Kyoto encyclopedia of genesand genomes |
| qRT-PCR | Real-time reverse transcription-polymerase chain reaction |
| EDTA | Ethylene Diamine Tetraacetic Acid |
| HE | hematoxylin and eosin |
References
- Dixon, M.J.; Marazita, M.L.; Beaty, T.H.; Murray, J.C. Cleft lip and palate: Understanding genetic and environmental influences. Nat. Rev. Genet. 2011, 12, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Mossey, P.A.; Little, J.; Munger, R.G.; Dixon, M.J.; Shaw, W.C. Cleft lip and palate. Lancet 2009, 374, 1773–1785. [Google Scholar] [CrossRef]
- Strauss, R.P. The organization and delivery of craniofacial health services: The state of the art. Cleft Palate Craniofac. J. 1999, 36, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Satokata, I.; Maas, R. Msx1 deficient mice exhibit cleft palate and abnormalities of craniofacial and tooth development. Nat. Genet. 1994, 6, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Helms, J.A. The role of sonic hedgehog in normal and abnormal craniofacial morphogenesis. Development 1999, 126, 4873–4884. [Google Scholar] [PubMed]
- Iwata, J.; Suzuki, A.; Pelikan, R.C.; Ho, T.V.; Sanchez-Lara, P.A.; Urata, M.; Dixon, M.J.; Chai, Y. Smad4-Irf6 genetic interaction and TGFβ-mediated IRF6 signaling cascade are crucial for palatal fusion in mice. Development 2013, 140, 1220–1230. [Google Scholar] [CrossRef] [PubMed]
- England, D.C.; Winters, L.M.; Carpenter, L.E. The development of a breed of miniature swine; a preliminary report. Growth 1954, 18, 207–214. [Google Scholar] [PubMed]
- Polejaeva, I.A.; Chen, S.H.; Vaught, T.D.; Page, R.L.; Mullins, J.; Ball, S.; Dai, Y.; Boone, J.; Walker, S.; Ayares, D.L.; et al. Cloned pigs produced by nuclear transfer from adult somatic cells. Nature 2000, 407, 86–90. [Google Scholar] [CrossRef]
- Wang, S.; Liu, Y.; Fang, D.; Shi, S. The miniature pig: A usefullarge animal model for dental and orofacial research. Oral Dis. 2007, 1, 530–537. [Google Scholar] [CrossRef]
- Wang, F.; Xiao, J.; Cong, W.; Li, A.; Wei, F.; Xu, J.; Zhang, C.; Fan, Z.; He, J.; Wang, S. Stage-specific differential gene expression profiling and functional network analysis during morphogenesis of diphyodont dentition in miniature pigs, Sus Scrofa. BMC Genom. 2014, 15, 103. [Google Scholar] [CrossRef]
- Ferguson, M.W. Palate development. Development 1988, 103, 41–60. [Google Scholar] [PubMed]
- Yoon, H.; Chung, I.S.; Seol, E.Y.; Park, B.Y.; Park, H.W. Development of the lip and palate in staged human embryos and early fetuses. Yonsei Med. J. 2000, 41, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, J.O.; Jiang, R. Palatogenesis: Morphogenetic and molecular mechanisms of secondary palate development. Development 2012, 139, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, J.; Liu, H.; Fan, Z.; Wang, S.; Du, J. A comprehensive study of palate development in miniature pig. Anat. Rec. 2017, 300, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Mou, Y.; Huang, Z.; Li, Y.; Han, L.; Zhang, Y.; Feng, Y.; Chen, Y.; Jiang, X.; Zhao, W.; et al. The sequence and analysis of a Chinese pig genome. Gigascience 2012, 1, 16. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.A.; Spacek, D.; Snyder, M.P. High-Throughput Sequencing Technologies. Mol. Cell. 2015, 58, 586–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.L.; Knott, L.; Halligan, E.; Yarram, S.J.; Mansell, J.P.; Sandy, J.R. Microarray analysis of murine palatogenesis: Temporal expression of genes during normal palate development. Dev. Growth Differ. 2003, 45, 153–165. [Google Scholar] [CrossRef] [PubMed]
- LaGamba, D.; Nawshad, A.; Hay, E.D. Microarray analysis of gene expression during epithelial-mesenchymal transformation. Dev. Dyn. 2005, 234, 132–142. [Google Scholar] [CrossRef]
- Bouleftour, W.; Juignet, L.; Bouet, G.; Granito, R.N.; Vanden-Bossche, A.; Laroche, N.; Aubin, J.E.; Lafage-Proust, M.H.; Vico, L.; Malaval, L. The role of the SIBLING, Bone Sialoprotein in skeletal biology—Contribution of mouse experimental genetics. Matrix Biol. 2016, 52–54, 60–77. [Google Scholar] [CrossRef]
- Stanier, P.; Moore, G.E. Genetics of cleft lip and palate: Syndromic genes contribute to the incidence of non-syndromic clefts. Hum. Mol. Genet. 2004, 13 (Suppl. 1), R73–R81. [Google Scholar] [CrossRef]
- Leslie, E.J.; Marazita, M.L. Genetics of cleft lip and cleft palate. Am. J. Med. Genet. C Semin. Med. Genet. 2013, 163, 246–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnick, M.; Chen, H.; Buckley, S.; Warburton, D.; Jaskoll, T. Insulin-like growth factor II receptor, transforming growth factor-beta, and Cdk4 expression and the developmental epigenetics of mouse palate morphogenesis and dysmorphogenesis. Dev. Dyn. 1998, 211, 11–25. [Google Scholar] [CrossRef]
- Gao, L.; Liu, Y.; Wen, Y.; Wu, W. LncRNA H19-mediated mouse cleft palate induced by all-trans retinoic acid. Hum. Exp. Toxicol. 2017, 36, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Riley, B.M.; Murray, J.C. Sequence evaluation of FGF and FGFR gene conserved non-coding elements in non-syndromic cleft lip and palate cases. Am. J. Med. Genet. A 2007, 143A, 3228–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, B.M.; Mansilla, M.A.; Ma, J.; Daack-Hirsch, S.; Maher, B.S.; Raffensperger, L.M.; Russo, E.T.; Vieira, A.R.; Dodé, C.; Mohammadi, M.; et al. Impaired FGF signaling contributes to cleft lip and palate. Proc. Natl. Acad. Sci. USA 2007, 104, 4512–4517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, B.D.; Buckalew, A.R.; Leffler, K.E. Effects of epidermal growth factor (EGF), transforming growth factor-alpha (TGFalpha), and 2,3,7,8-tetrachlorodibenzo-p-dioxin on fusion of embryonic palates in serum-free organ culture using wild-type, EGF knockout, and TGFalpha knockout mouse strains. Birth Defects Res. A Clin. Mol. Teratol. 2005, 73, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lan, Y.; Krumlauf, R.; Jiang, R. Modulating Wnt signaling rescues palate morphogenesis in Pax9 mutant mice. J. Dent. Res. 2017, 96, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Peters, H.; Neubüser, A.; Kratochwil, K.; Balling, R. Pax9-deficient mice lack pharyngeal pouch derivatives and teeth and exhibit craniofacial and limb abnormalities. Genes Dev. 1998, 12, 2735–2747. [Google Scholar] [CrossRef]
- Richardson, R.; Mitchell, K.; Hammond, N.L.; Mollo, M.R.; Kouwenhoven, E.N.; Wyatt, N.D.; Donaldson, I.J.; Zeef, L.; Burgis, T.; Blance, R.; et al. p63 exerts spatio-temporal control of palatal epithelial cell fate to prevent cleft palate. PLoS Genet. 2017, 13, e1006828. [Google Scholar] [CrossRef]
- Ferone, G.; Thomason, H.A.; Antonini, D.; De Rosa, L.; Hu, B.; Gemei, M.; Zhou, H.; Ambrosio, R.; Rice, D.P.; Acampora, D.; et al. Mutant p63 causes defective expansion of ectodermal progenitor cells and impaired FGF signalling in AEC syndrome. EMBO Mol. Med. 2012, 4, 192–205. [Google Scholar] [CrossRef]
- Aylward, A.; Cai, Y.; Lee, A.; Blue, E.; Rabinowitz, D.; Haddad, J., Jr. University of Washington Center for Mendelian Genomics. Using whole exome sequencing to identify candidate genes with rare variants in nonsyndromic cleft lip and palate. Genet. Epidemiol. 2016, 40, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zuo, X.; He, M.; Gao, J.; Fu, Y.; Qin, C.; Meng, L.; Wang, W.; Song, Y.; Cheng, Y.; et al. Genome-wide analyses of non-syndromic cleft lip with palate identify 14 novel loci and genetic heterogeneity. Nat. Commun. 2017, 8, 14364. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.C.; Nidey, N.L.; Butali, A.; Buxo, C.J.; Christensen, K.; Deleyiannis, F.W.; Hecht, J.T.; Field, L.L.; Moreno-Uribe, L.M.; Orioli, I.M.; et al. Genome-wide interaction studies identify sex-specific risk alleles for nonsyndromic orofacial clefts. Genet. Epidemiol. 2018, 42, 664–672. [Google Scholar] [CrossRef]
- Seelan, R.S.; Mukhopadhyay, P.; Pisano, M.M.; Greene, R.M. Developmental epigenetics of the murine secondary palate. ILAR J. 2012, 53, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Alvarez, C.; Tudela, C.; Pérez-Miguelsanz, J.; O’Kane, S.; Puerta, J.; Ferguson, M.W. Medial edge epithelial cell fate during palatal fusion. Dev. Biol. 2000, 220, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Tian, Z.; Miao, C.; Wang, J.; Wang, S.; Du, J. The histological study of miniature pig early tongue development. Beijing J. Stomatol. 2017, 24, 301–304. (In Chinese) [Google Scholar]
- Okano, J.; Udagawa, J.; Shiota, K. Roles of retinoic acid signaling in normal and abnormal development of the palate and tongue. Congenit. Anom. (Kyoto) 2014, 54, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaffran, S.; Odelin, G.; Stefanovic, S.; Lescroart, F.; Etchevers, H.C. Ectopic expression of Hoxb1 induces cardiac and craniofacial malformations. Genesis 2018, 56, e23221. [Google Scholar] [CrossRef] [Green Version]
- Machon, O.; Masek, J.; Machonova, O.; Krauss, S.; Kozmik, Z. Meis2 is essential for cranial and cardiac neural crest development. BMC Dev. Biol. 2015, 15, 40. [Google Scholar] [CrossRef]
- León, M.; Ferreira, C.R.; Eberlin, L.S.; Jarmusch, A.K.; Pirro, V.; Rodrigues, A.C.B.; Favaron, P.O.; Miglino, M.A.; Cooks, R.G. Metabolites and lipids associated with fetal swine anatomy via desorption electrospray ionization—Mass spectrometry imaging. Sci. Rep. 2019, 9, 7247. [Google Scholar] [CrossRef]
- Yuan, X.; Yang, S. Primary Cilia and Intraflagellar Transport Proteins in Bone and Cartilage. J. Dent. Res. 2016, 95, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Schock, E.N.; Struve, J.N.; Chang, C.F.; Williams, T.J.; Snedeker, J.; Attia, A.C.; Stottmann, R.W.; Brugmann, S.A. A tissue-specific role for intraflagellar transport genes during craniofacial development. PLoS ONE 2017, 12, e0174206. [Google Scholar] [CrossRef] [PubMed]
- Laitala, T.; Vaananen, H.K. Inhibition of bone resorption in vitro by antisense RNA and DNA molecules targeted against carbonic anhydrase II or two subunits of vacuolar H (+)-ATPase. J. Clin. Investig. 1994, 93, 2311–2318. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.; Kaartinen, V. Signaling networks in palate development. Wiley Interdiscip. Rev. Syst. Biol. Med. 2014, 6, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Tarr, J.T.; Lambi, A.G.; Bradley, J.P.; Barbe, M.F.; Popoff, S.N. Development of normal and cleft palate: A central role for connective tissue growth factor (CTGF)/CCN2. J. Dev. Biol. 2018, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Fakhouri, W.D.; Rhea, L.; Du, T.; Sweezer, E.; Morrison, H.; Fitzpatrick, D.; Yang, B.; Dunnwald, M.; Schutte, B.C. MCS9.7 enhancer activity is highly, but not completely, associated with expression of Irf6 and p63. Dev. Dyn. 2012, 241, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Gao, Y.; Lan, Y.; Jia, S.; Jiang, R. Pax9 regulates a molecular network involving Bmp4, Fgf10, Shh signaling and the Osr2 transcription factor to control palate morphogenesis. Development 2013, 140, 4709–4718. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Klement, J.F.; Leperi, D.A.; Birk, D.E.; Sasaki, T.; Timpl, R.; Uitto, J.; Pulkkinen, L. Targeted inactivation of murine laminin gamma2-chain gene recapitulates human junctional epidermolysis bullosa. J. Investig. Dermatol. 2003, 121, 720–731. [Google Scholar] [CrossRef]
- Huang, H.Y.; Liu, R.R.; Zhao, G.P.; Li, Q.H.; Zheng, M.Q.; Zhang, J.J.; Li, S.F.; Liang, Z.; Wen, J. Integrated analysis of microRNA and mRNA expression profiles in abdominal adipose tissues in chickens. Sci. Rep. 2015, 5, 16132. [Google Scholar] [CrossRef]
- Du, J.; Ma, Y.; Ma, P.; Wang, S.; Fan, Z. Demethylation of epiregulin gene by histone demethylase FBXL11 and BCL6 corepressor inhibits osteo/dentinogenic differentiation. Stem Cells 2013, 31, 126–136. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Chen, J.; Yuan, D.; Sun, L.; Fan, Z.; Wang, S.; Du, J. Dynamic mRNA Expression Analysis of the Secondary Palatal Morphogenesis in Miniature Pigs. Int. J. Mol. Sci. 2019, 20, 4284. https://doi.org/10.3390/ijms20174284
Liu J, Chen J, Yuan D, Sun L, Fan Z, Wang S, Du J. Dynamic mRNA Expression Analysis of the Secondary Palatal Morphogenesis in Miniature Pigs. International Journal of Molecular Sciences. 2019; 20(17):4284. https://doi.org/10.3390/ijms20174284
Chicago/Turabian StyleLiu, Jia, Jing Chen, Dong Yuan, Lindong Sun, Zhipeng Fan, Songlin Wang, and Juan Du. 2019. "Dynamic mRNA Expression Analysis of the Secondary Palatal Morphogenesis in Miniature Pigs" International Journal of Molecular Sciences 20, no. 17: 4284. https://doi.org/10.3390/ijms20174284
APA StyleLiu, J., Chen, J., Yuan, D., Sun, L., Fan, Z., Wang, S., & Du, J. (2019). Dynamic mRNA Expression Analysis of the Secondary Palatal Morphogenesis in Miniature Pigs. International Journal of Molecular Sciences, 20(17), 4284. https://doi.org/10.3390/ijms20174284