Anti-Inflammatory Response and Muscarinic Cholinergic Regulation during the Laxative Effect of Asparagus cochinchinensis in Loperamide-Induced Constipation of SD Rats

 and
and 
Abstract
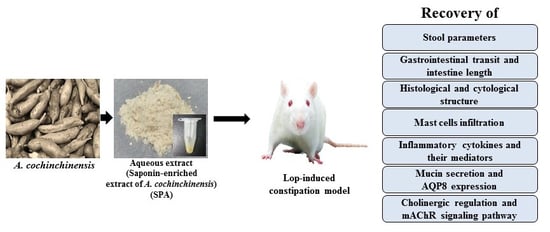
:
1. Introduction
2. Results
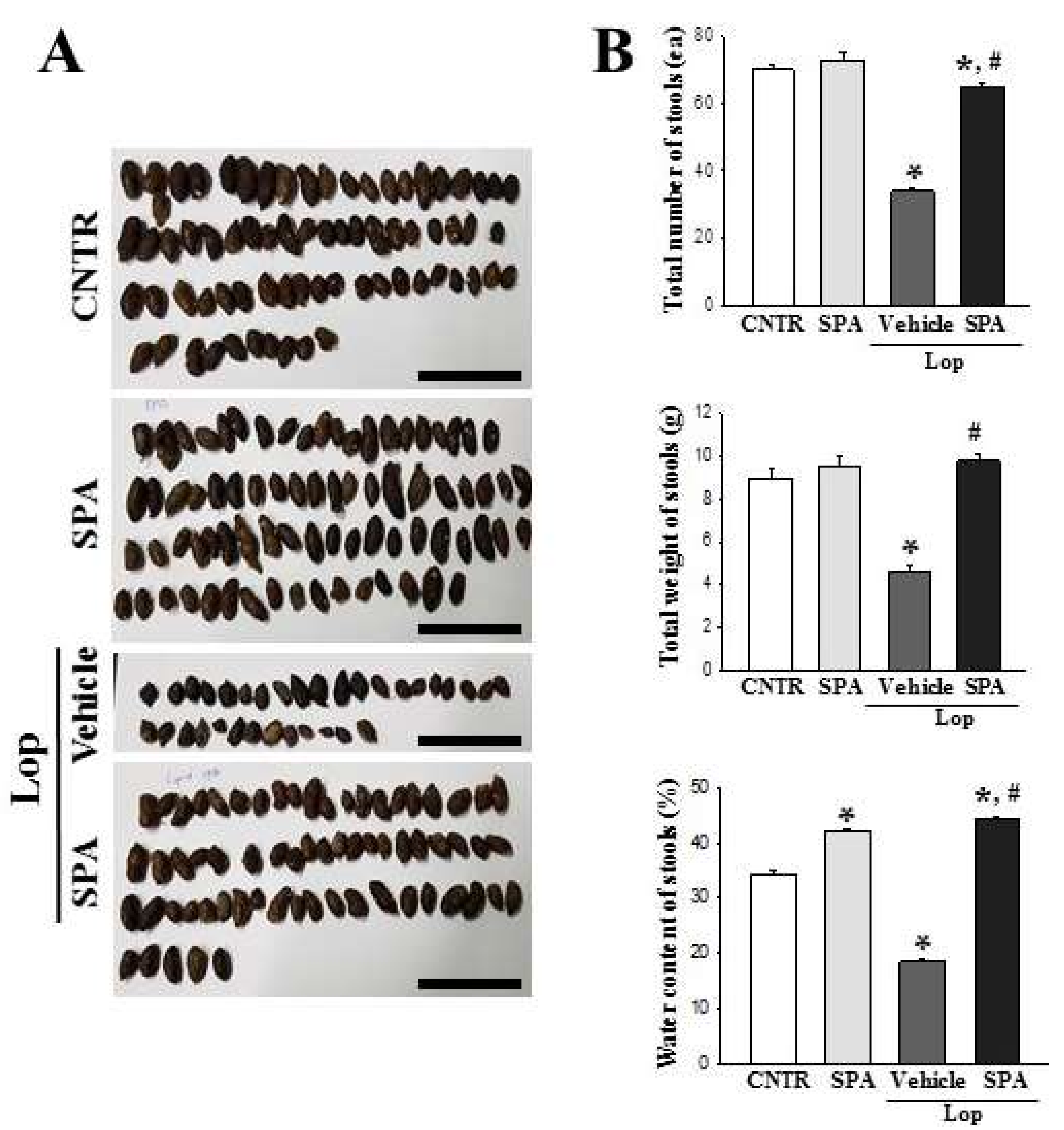
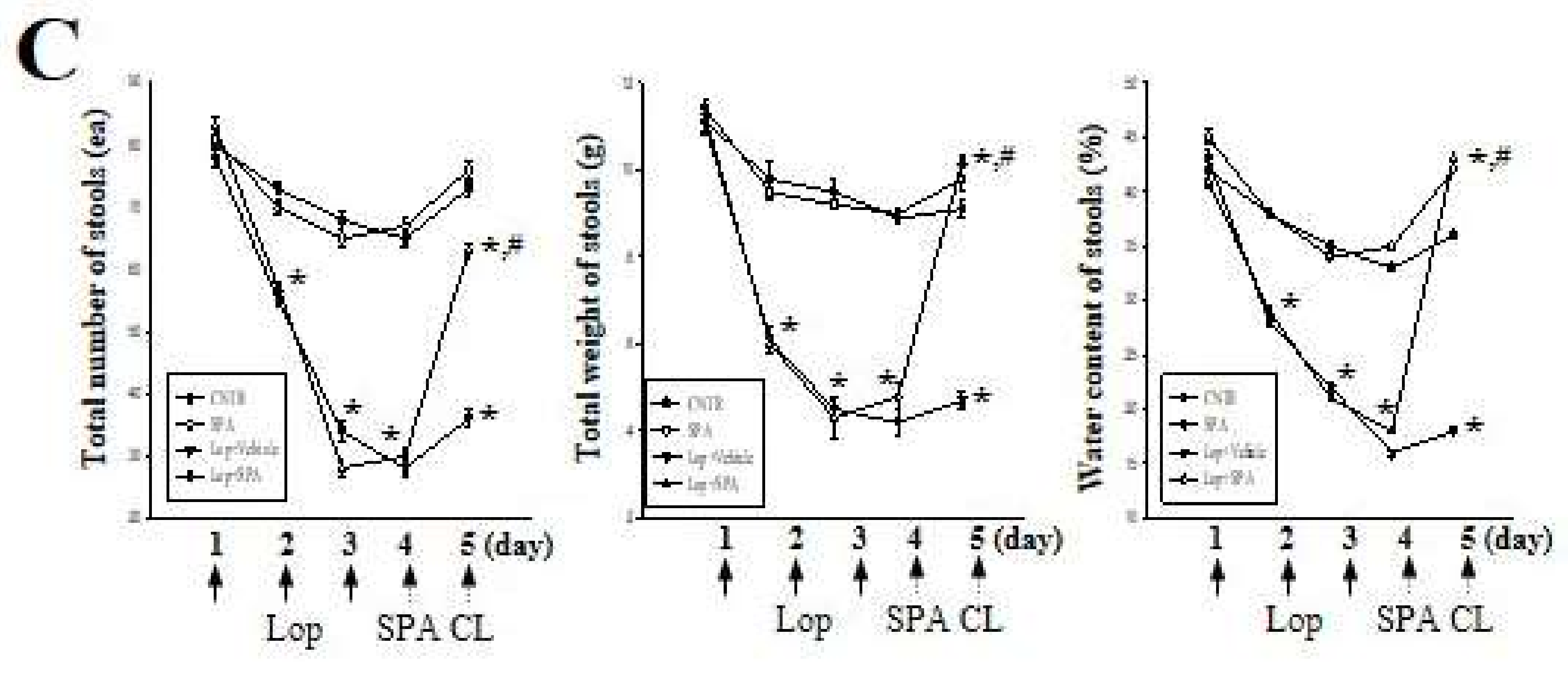
2.1. Effect of SPA Administration on the Feeding Behavior and Excretion Parameters
2.2. Effect of SPA Administration Gastrointestinal Motility
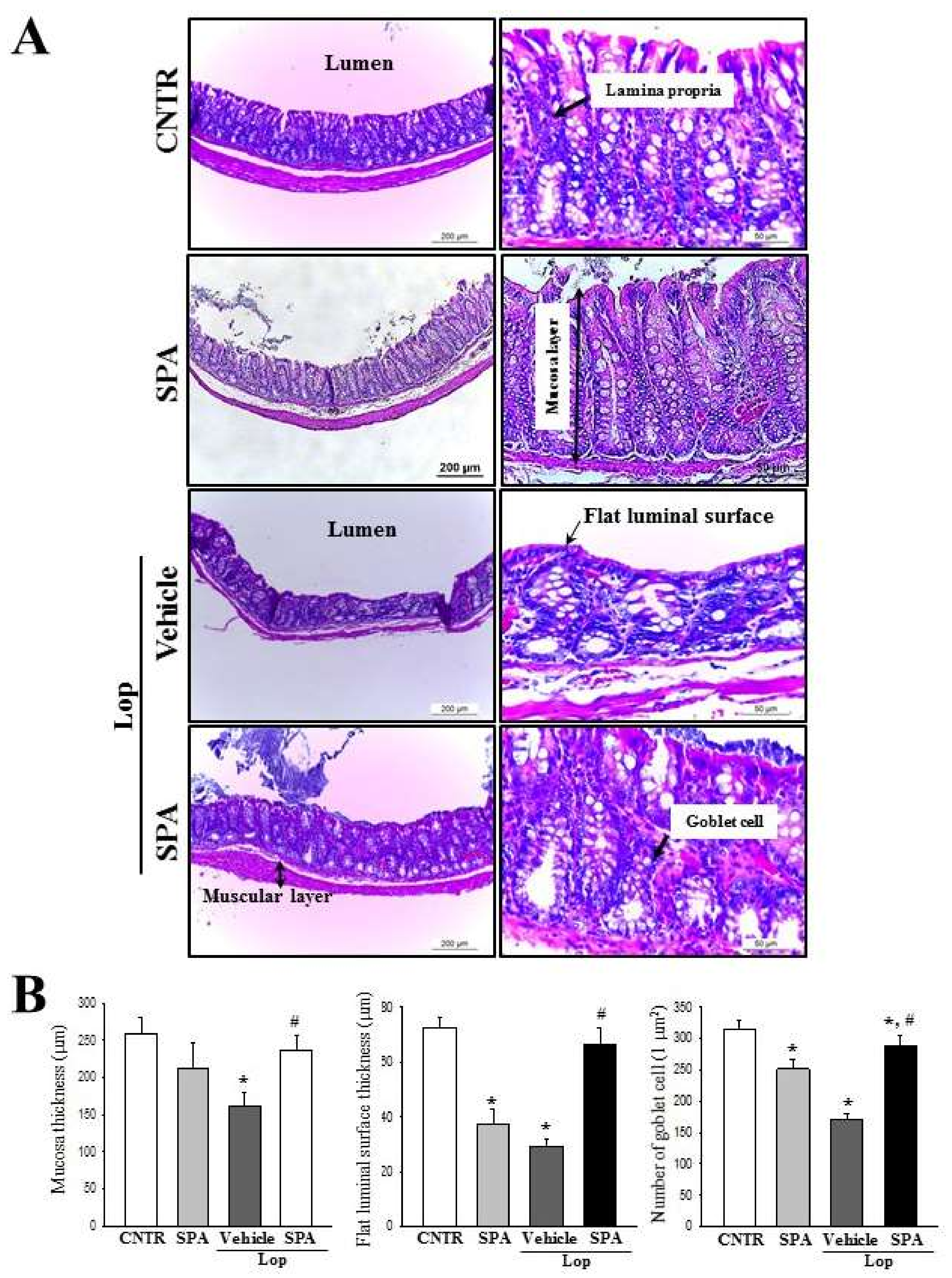
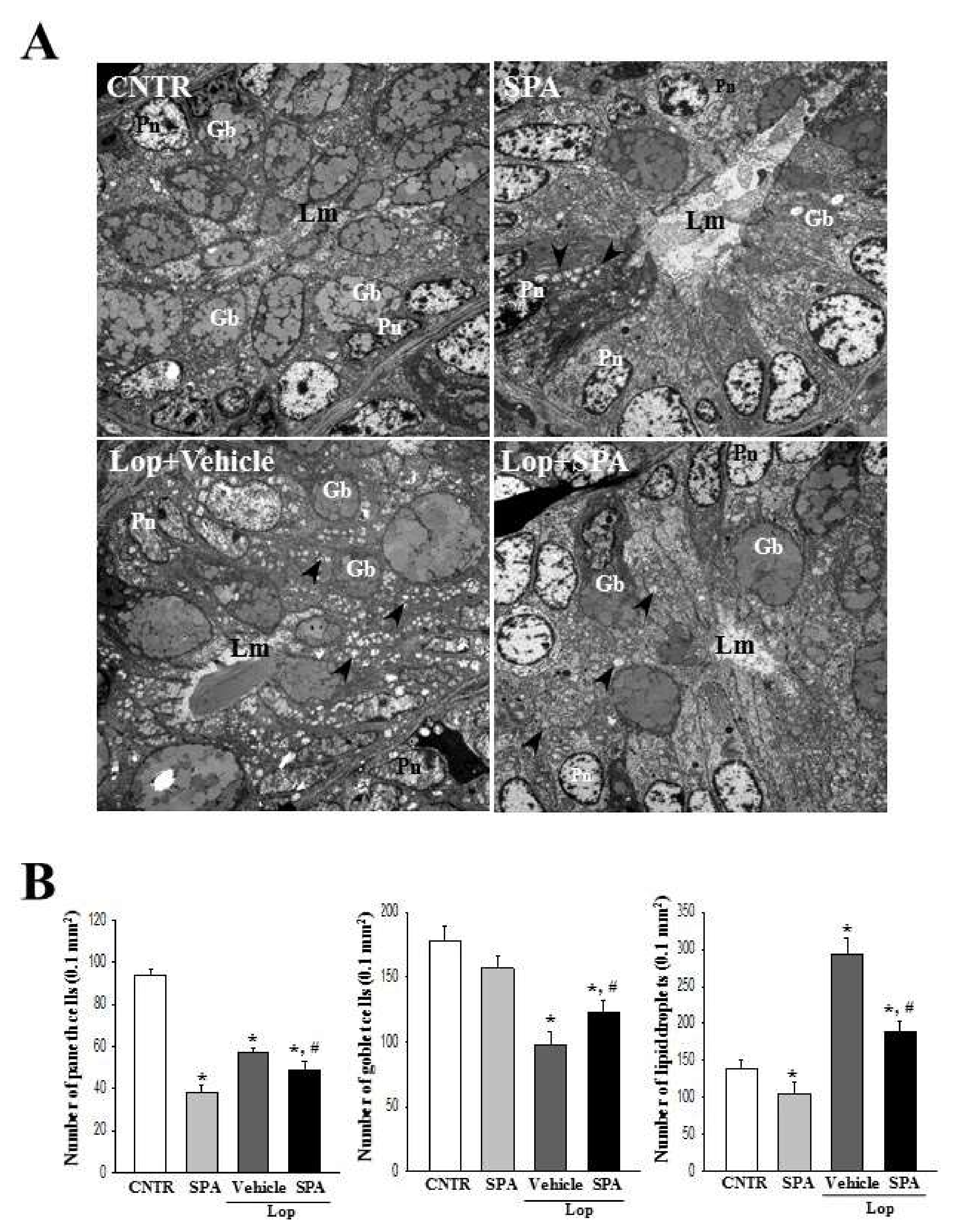
2.3. Effect of SPA Administration on Histological Structure of the Colon
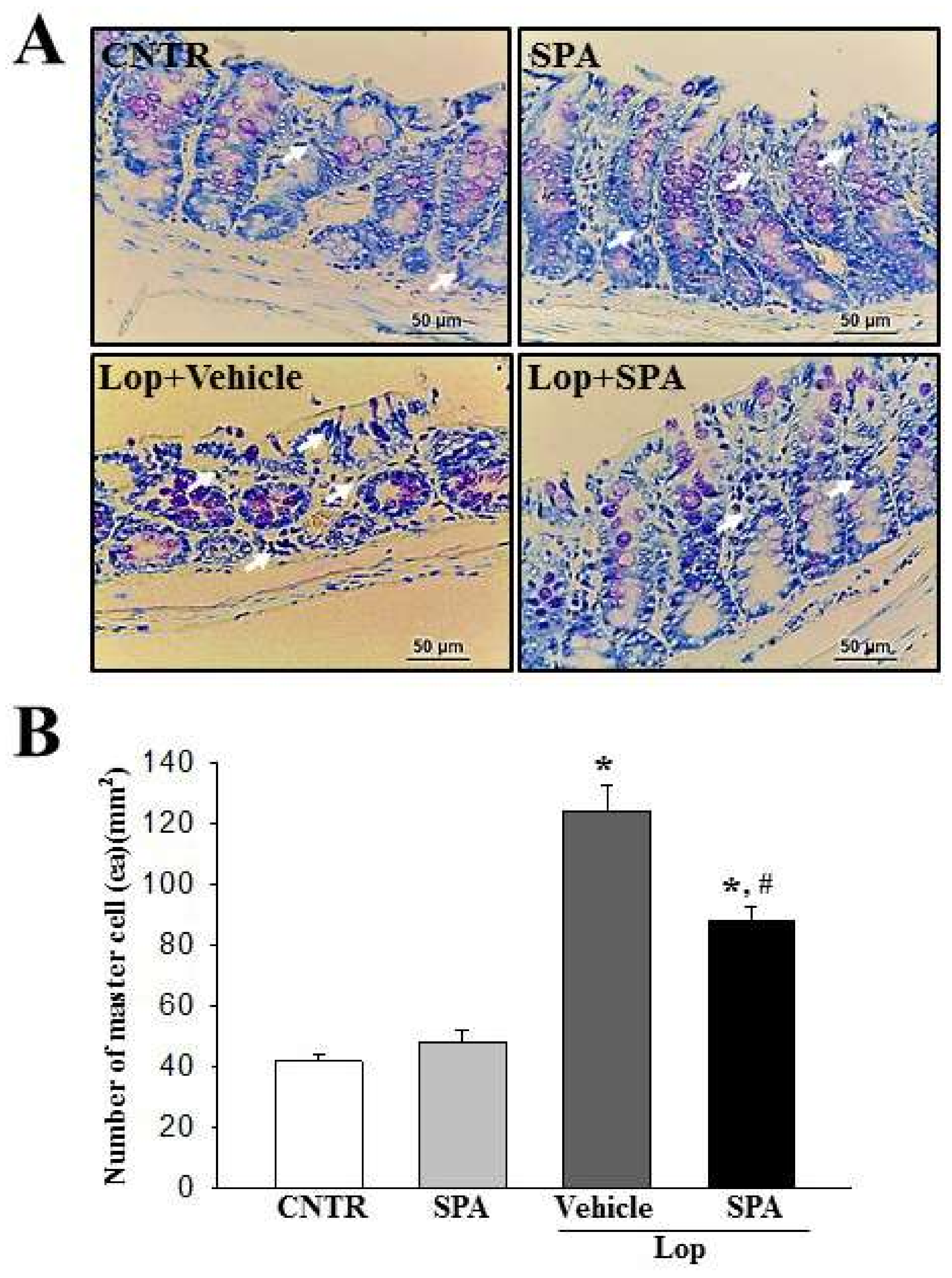
2.4. Effect of SPA Administration on Mast Cells Infiltration
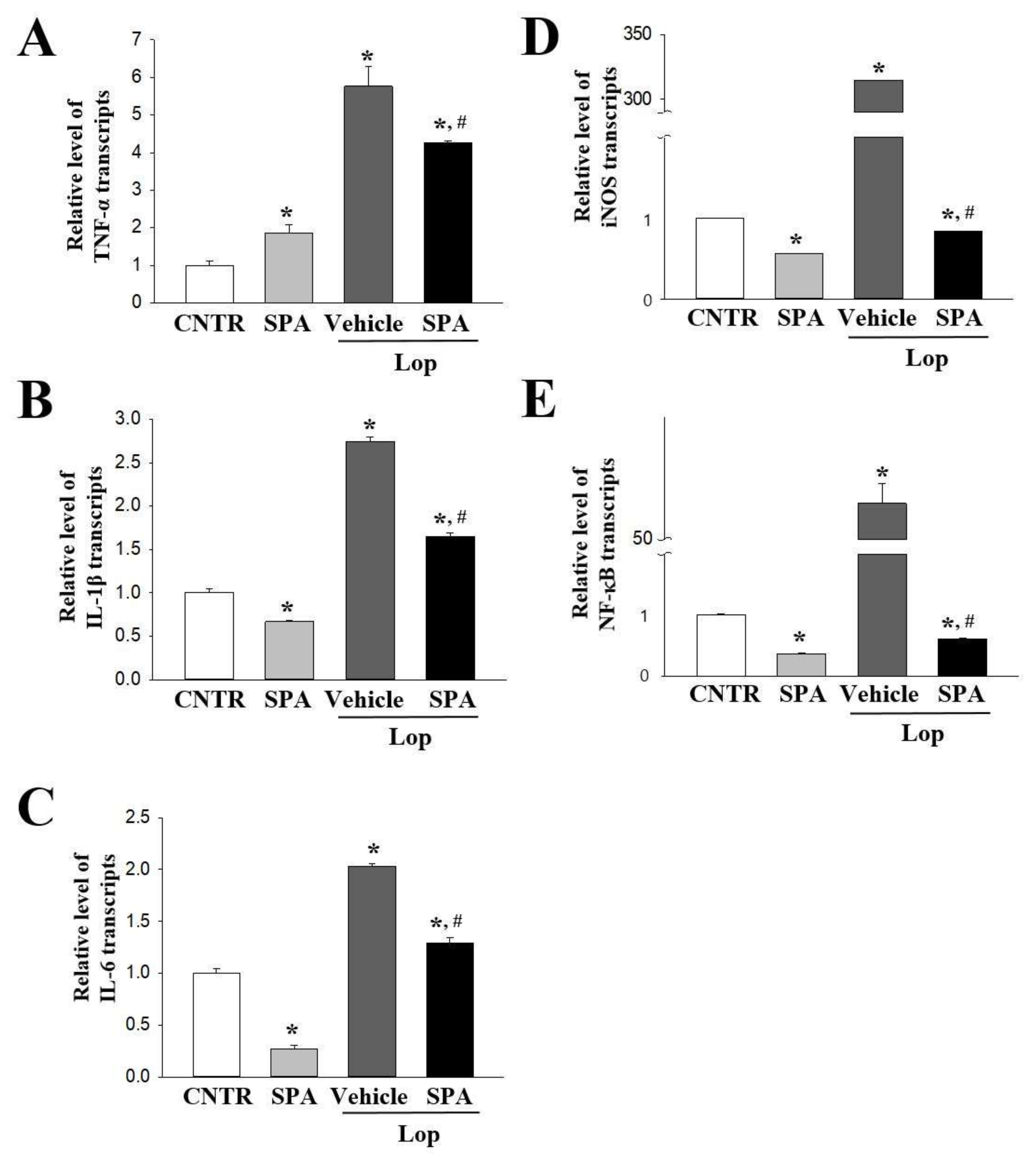
2.5. Effect of SPA Administration on the Expression of Inflammatory Cytokines and their Mediators
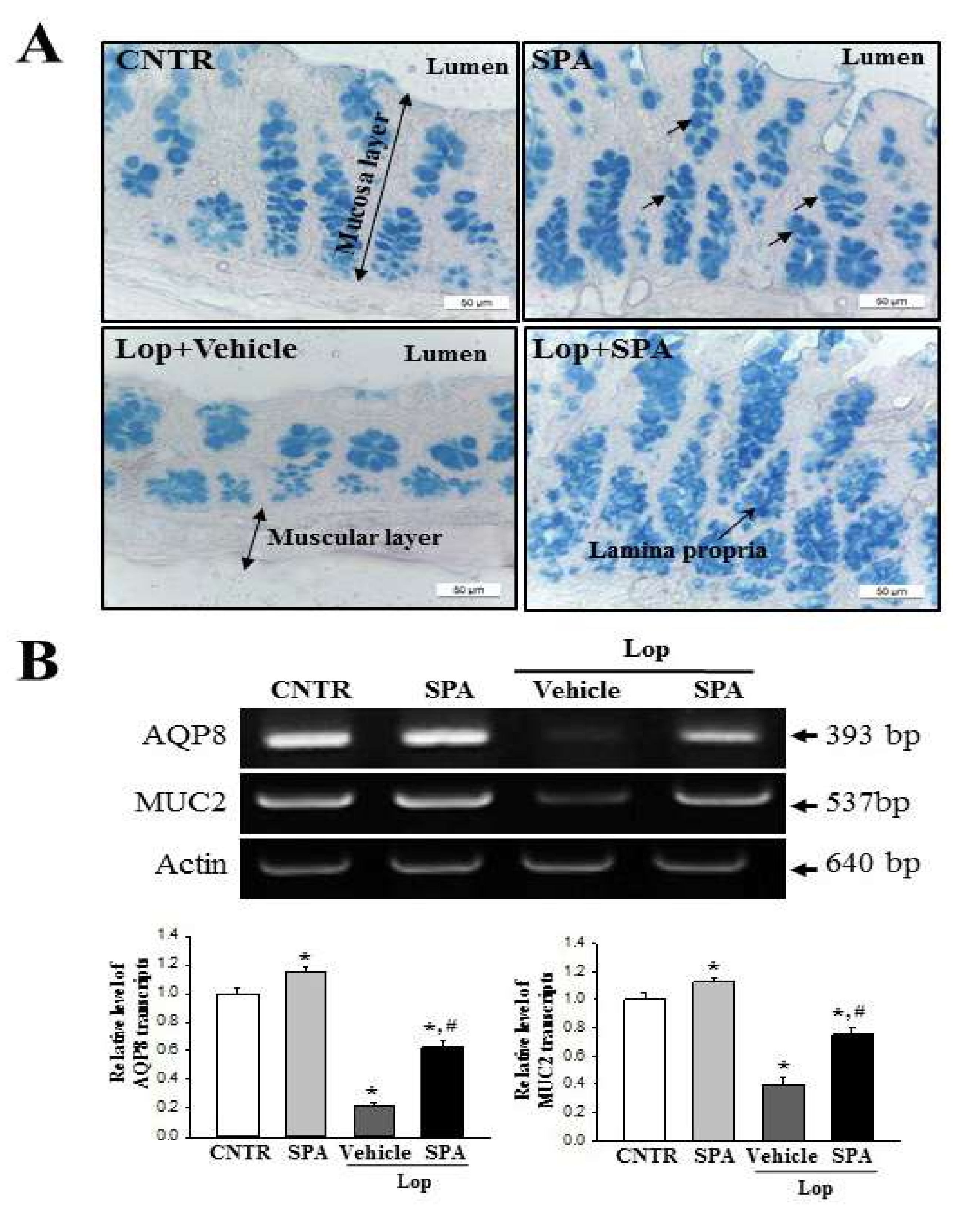
2.6. Effect of SPA Administration on Regulation of Inflammation-Derived Mucin Secretion
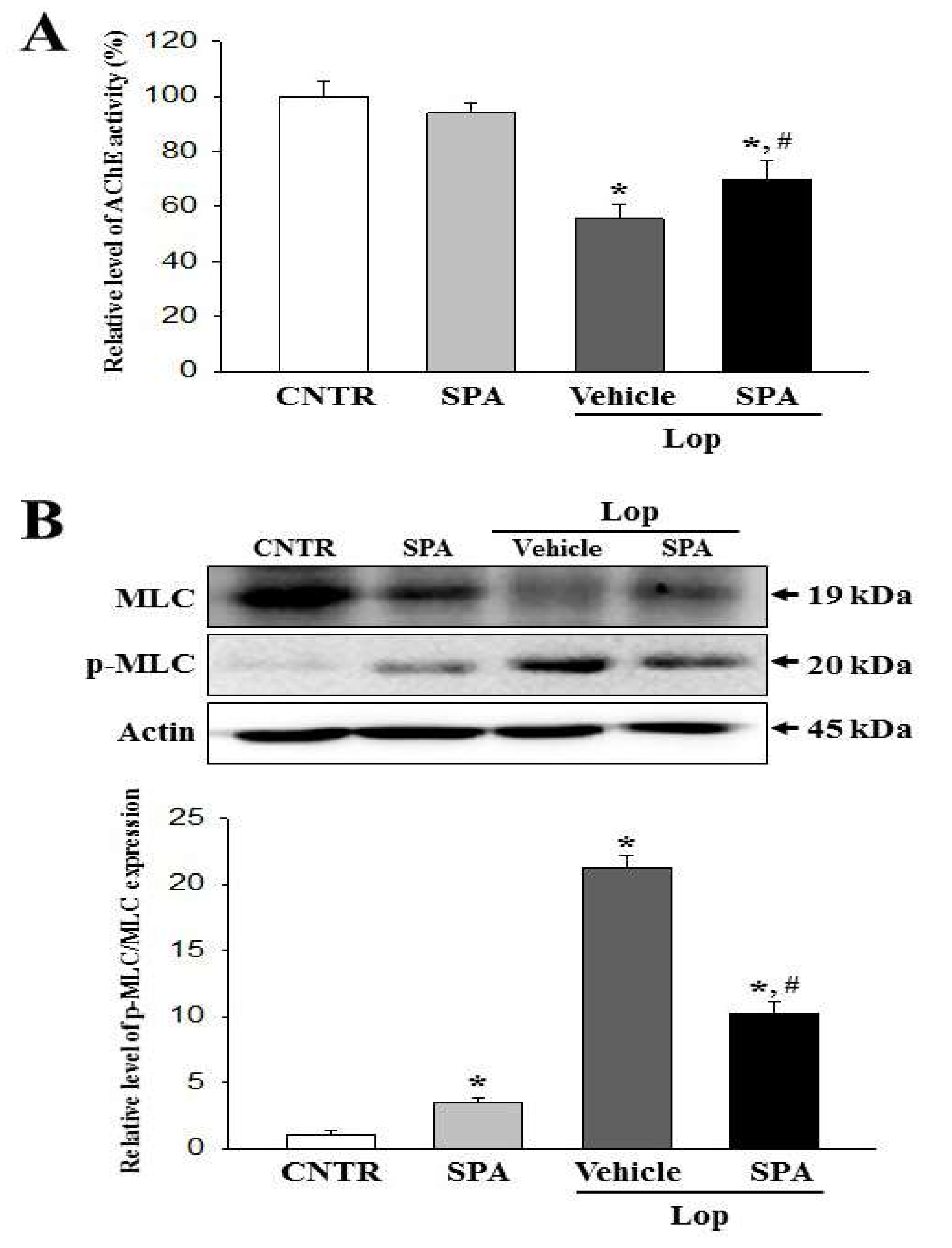
2.7. Mechanism of Action of SPA on the Cholinergic Regulation of Gastrointestinal Mobility
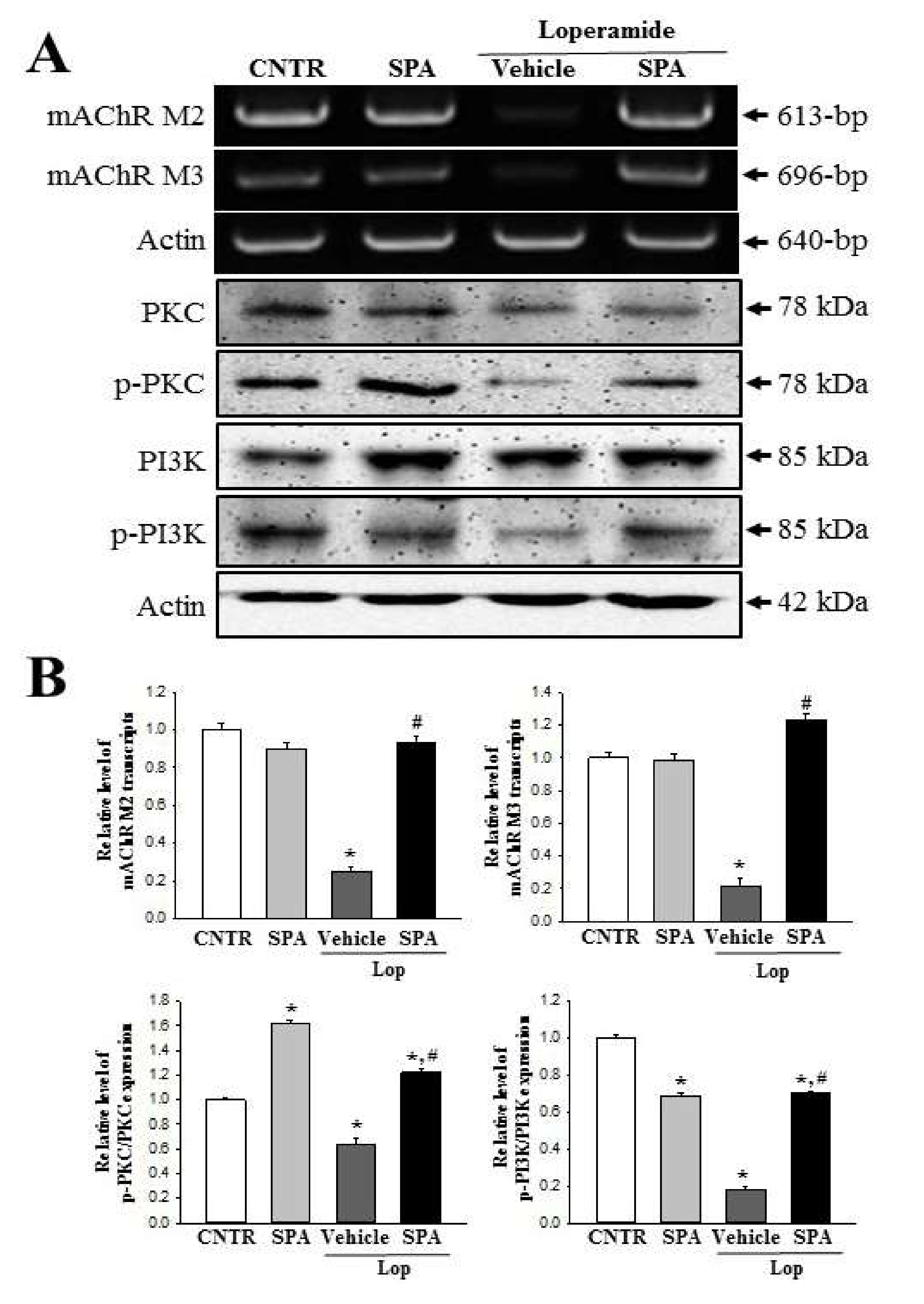
2.8. Molecular Mechanism of the Laxative Effects of SPA on the Downstream Signaling Pathway of mAChRs
3. Discussion
4. Materials and Methods
4.1. Preparation of SPA
4.2. Experimental Design for Animal Study
4.3. Gastrointestinal Transit Ratio Analysis
4.4. Analysis of Food Intake, Water Intake, and Body Weight
4.5. Measurement of Stool Parameters and Urine Volume
4.6. Western Blotting
4.7. Semiquantitative PCR Anlysis
4.8. Quantitative Real-Time PCR Analysis
4.9. Histopathological Analysis
4.10. Ultrastructure Analysis Using Transmission Electron Microscopy (TEM)
4.11. AChE Activity Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SPA | saponin-enriched extracts of Asparagus cochinchinensis |
| Lop | loperamide |
| AChE | acetylcholine esterase |
| MLC | myosin light chain |
| AEtLP | aqueous extract of L. platyphylla |
| RLP | Red L. platyphylla |
| GEGR | gallotanin-enriched galla rhois |
| CLP | cecal ligation and puncture |
| DSS | dextran sulfate sodium |
| HPLC | high performance liquid chromatography |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| KFDA | Korea Food and Drug Administration |
| AAALAC | Association for Assessment and Accreditation of Laboratory Animal Care |
| SD | Sprague Dawley |
| SPF | specific pathogen-free |
| RT-PCR | Reverse transcription polymerase chain reaction |
| H&E | hematoxylin and eosin |
| TEM | transmission electron microscopy |
| SOD | superoxide dismutase |
| MDA | malondialdehyde |
| NGF | nerve growth factor |
| MP | methyl protodioscin |
| TPA | 12-O-tetradecanoyl-phorbol-13-acetate |
| LPS | Lipopolysaccharide |
| BAW | butanol extract of A. cochinchinensis roots fermented with Weissella cibaria |
| OVA | ovalbumin |
| FMT | fecal microbiota transplantation |
References
- Walia, R.; Mahajan, L.; Steffen, R. Recent advances in chronic constipation. Curr. Opin. Pediatr. 2009, 21, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Voderholzer, W.A.; Schatke, W.; Muhldorfer, B.E.; Klauser, A.G.; Birkner, B.; Muller-Lissner, S.A. Clinical response to dietary fiber treatment of chronic constipation. Am. J. Gastroenterol. 1997, 92, 95–98. [Google Scholar] [PubMed]
- Lembo, A.; Camilleri, M. Chronic constipation. N. Engl. J. Med. 2013, 349, 1360–1368. [Google Scholar] [CrossRef] [PubMed]
- Busti, A.J.; Murillo, J.R., Jr.; Cryer, B. Tegaserod-induced myocardial infarction: Case report and hypothesis. Parmacotherapy 2004, 24, 526–531. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, Y.J.; Kwak, M.H.; Ko, J.; Hong, J.T.; Hwang, D.Y. Aquous extracts of Liriope platyphylla induced significant laxative effects on loperamide-induced constipation of SD rats. BMC Complement. Altern. Med. 2013, 13, 333. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Xiong, Y.; Chen, D.; Li, Y.; Xu, B.; Lin, Y.; Tang, Z.; Jiang, C.; Wang, L. Ameliorative effects of atractylodin on intestinal inflammation and co-occurring dysmotility in both constipation and diarrhea prominent rats. Korean J. Physiol. Pharmacol. 2017, 21, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.E.; Go, J.; Koh, E.K.; Song, S.H.; Sung, J.E.; Lee, H.A.; Lee, Y.H.; Hong, J.T.; Hwang, D.Y. Gallotannin-enriched extract isolated from Galla Rhois may be a functional candidate with laxative effects for treatment of loperamide-induced constipation of SD rats. PLoS ONE 2016, 11, e0161144. [Google Scholar] [CrossRef]
- Kim, J.E.; Go, J.; Lee, H.S.; Hong, J.T.; Hwang, D.Y. Spicatoside A in red Liriope platyphylla displays a laxative effect in a constipation rat model via regulating mAChRs and ER stress signaling. Int. J. Mol. Med. 2019, 43, 185–198. [Google Scholar] [CrossRef]
- Hara, H.; Ise, Y.; Morimoto, N.; Shimazawa, M.; Ichihashi, K.; Ohyama, M.; Iinuma, M. Laxative effect of agarwood leaves and its mechanism. Biosci. Biotechnol. Biochem. 2008, 72, 335–345. [Google Scholar] [CrossRef]
- Meite, S.; Bahi, C.; Yeo, D.; Datte, J.Y.; Djaman, J.A.; Nguessan, D.J. Laxativeactivities of Mareya micrantha (Benth.) Müll. Arg. (Euphorbiaceae) leaf aqueous extract in rats. BMC Complement. Altern. Med. 2010, 10, 7–10. [Google Scholar] [CrossRef]
- Guarize, L.; Costa, J.C.; Dutra, L.B.; Mendes, R.F.; Lima, I.V.A.; Scio, E. Anti-inflammatory, laxative and intestinal motility effects of Senna Macranthera leaves. Nat. Prod. Res. 2012, 26, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Wintola, O.A.; Sunmonu, T.O.; Afolayan, A.J. The effect of Aloe ferox Mill. in the treatmentet of loperamide-induced constipation in Wistar rats. BMC Gastroenterol. 2010, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Rehman, N.; Mehmood, M.H.; Al-Rehaily, A.J.; Mothana, R.A.A.; Gilani, A.H. Species and tissue-specificity of prokinetic, laxative and spasmodic effects of Fumaria parviflora. BMC Complement. Altern. Med. 2012, 12, 16. [Google Scholar]
- Abbas, S.; Bashir, S.; Khan, A.; Mehmood, M.H.; Gilani, A.H. Gastrointestinal stimulant effect of Urginea indica Kunth. and involvement of muscarinic receptors. Phytother. Res. 2012, 26, 704–408. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Kim, J.H.; Jeung, H.W.; Lee, C.U.; Kim, D.S.; Li, B.; Lee, G.H.; Sung, M.S.; Ha, K.C.; Back, H.I.; et al. Effects of Ficus carica paste on loperamide-induced constipation in rats. Food Chem. Toxicol. 2012, 50, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.H.; Xu, X.X.; Pan, R.; Li, Y.; Luo, Y.B.; Xia, Y.F.; Murata, K.; Matsuda, H.; Dai, Y. Saponin fraction from Astragalus membranaceus roots protects mice against polymicrobial sepsis induced by cecal ligation and puncture by inhibiting inflammation and upregulating protein C pathway. J. Nat. Med. 2009, 63, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.D.; Kang, S.H.; Bang, K.S.; Chang, Y.N.; Lee, J.H.; Jin, J.S. Glycyrrhetic acid ameliorates dextran sulfate sodium-induced Ulcerative colitis in vivo. Molecules 2016, 21, 523. [Google Scholar] [CrossRef]
- Zou, Y.; Tao, T.; Tian, Y.; Zhu, J.; Cao, L.; Deng, X.; Li, J. Ginsenoside Rg1 improves survival in a murine model of polymicrobial sepsis by suppressing the inflammatory response and apoptosis of lymphocytes. J. Surg. Res. 2013, 183, 760–766. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, D.; Lv, B.; Liu, F.; Yao, Q.; Tang, Z.; Lin, Y. Effects of ginsenoside Re on rat jejunal contractility. J. Nat. Med. 2014, 68, 530–538. [Google Scholar] [CrossRef]
- Chen, D.; Xiong, Y.; Jiang, C.; Lv, B.; Liu, F.; Wang, L.; Lin, Y. Effects of ginsenosides on rat jejunal contractility. Pharm. Biol. 2014, 52, 162–168. [Google Scholar] [CrossRef]
- Negi, J.S.; Singh, P.; Joshi, G.P.; Rawat, M.S.; Bisht, V.K. Chemical constituents of Asparagus. Phcog. Rev. 2010, 4, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Kim, J.E.; Song, S.H.; Sung, J.E.; Jung, M.G.; Kim, D.S.; Hong, J.S.; Lee, C.Y.; Lee, S.H.; Hwang, D.Y. Effects of an aqueous extract of Asparagus cochinchinensis on the regulation of nerve growth factor in neuronal cells. J. Life Sci. 2016, 26, 509–518. [Google Scholar] [CrossRef]
- Xiong, D.; Yu, L.X.; Yan, X.; Guo, C.; Xiong, Y. Effects of root and stem extracts of Asparagus cochinchinensis on biochemical indicators related to aging in the brain and liver of mice. Am. J. Chin. Med. 2011, 39, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jin, L.H. Asparagus cochinchinensis extract alleviates metal ion-induced gut injury in drosophila: An in silico analysis of potential active constituents. Evid. Based Complement. Altern. Med. 2016, 7603746. [Google Scholar] [CrossRef]
- Le, V.T.; Nguyen, G.C. Selected Medicinal Plants in Vietnam; Science and Technology Publishing House: Hanoi, Vietnam, 1999; Volume 1, p. 439. [Google Scholar]
- Zeng, G.P.; Pan, Z.P.; Huang, L.X.; Zhao, Z. Quality of Asparagus cochinchinensis from different producing areas in Guizhou. Agric. Sci. 2011, 12, 45–47. [Google Scholar]
- Li, Z.X.; Huang, C.G.; Cai, Y.J.; Chen, X.M.; Wang, F.; Chen, Y.Z. The chemical structure and antioxidative activity of polysaccharide from Asparagus cochinchinensis. Acta Pharm. Sin. 2000, 5, 358–362. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in burkina fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Hassan, S.M.; Al Aqil, A.A.; Attimarad, M. Determination of crude saponin and total flavonoids content in guar meal. Adv. Med. Plant Res. 2013, 1, 24–28. [Google Scholar]
- Lee, D.Y.; Choo, B.K.; Yoon, T.; Cheon, M.S.; Lee, H.W.; Lee, A.Y.; Kim, H.K. Anti-inflammatory effects of Asparagus cochinchinensis extract in acute and chronic cutaneous inflammation. J. Ethnopharmacol. 2009, 121, 28–34. [Google Scholar] [CrossRef]
- Lee, J.H.; Lim, H.J.; Lee, C.W.; Son, K.H.; Son, J.K.; Lee, S.K.; Kim, H.P. Methyl protodioscin from the roots of Asparagus cochinchinensis attenuates airway inflammation by inhibiting cytokine production. Evid. Based Complement. Altern. Med. 2015, 640846. [Google Scholar] [CrossRef]
- Sung, J.E.; Lee, H.A.; Kim, J.E.; Yun, W.B.; An, B.S.; Yang, S.Y.; Kim, D.S.; Lee, C.Y.; Lee, H.S.; Bae, C.J.; et al. Saponin-enriched extract of Asparagus cochinchinensis alleviates airway inflammation and remodeling in ovalbumin-induced asthma model. Int. J. Mol. Med. 2017, 40, 1365–1376. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Kim, J.E.; Park, J.J.; Lee, M.R.; Song, B.R.; Park, J.W.; Kang, M.J.; Lee, H.S.; Son, H.J.; Hong, J.T.; et al. The anti-inflammatory effects of fermented herbal roots of Asparagus cochinchinensis in an ovalbumin-induced asthma model. J. Clin. Med. 2018, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhu, Q.; Qu, X.; Qin, H. Microbial treatment in chronic constipation. Sci. China Life Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Suo, H.Y.; Qian, Y.; Li, G.J.; Liu, Z.H.; Li, J. Therapeutic effects of Lactobacillus casei Qian treatment in activated carbon-induced constipated mice. Mol. Med. Rep. 2015, 12, 3191–3199. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, I.; Robinson, L.; Verhelst, A.; Marzorati, M.; Winkens, B.; Abbeele, P.V.; Possemiers, S.A. Yeast fermentate improves gastrointestinal discomfort and constipation by modulation of the gut microbiome: Results from a randomized double-blind placebo-controlled pilot trial. BMC Complement. Altern. Med. 2017, 17, 441. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Zhao, Y.; Yu, Y.B. Intestinal microbiota and chronic constipation. Springerplus 2016, 5, 1130. [Google Scholar] [CrossRef]
- Tottey, W.; Feria-Gervasio, D.; Gaci, N.; Laillet, B.; Pujos, E.; Martin, J.F.; Sebedio, J.L.; Sion, B.; Jarrige, J.F.; Alric, M.; et al. Colonic Transit Time Is a Driven Force of the Gut Microbiota Composition and Metabolism: In Vitro Evidence. J. Neurogastroenterol. Motil. 2017, 23, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Verbeke, K.A.; Boobis, A.R.; Chiodini, A.; Edwards, C.A.; Franck, A.; Kleerebezem, M.; Nauta, A.; Raes, J.; van Tol, E.A.; Tuohy, K.M. Towards microbial fermentation metabolites as markers for health benefits of prebiotics. Nutr. Res. Rev. 2015, 28, 42–66. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.H.; Yu, H.J.; Pan, A.; Du, J.Y.; Ruan, Y.C.; Ko, W.H.; Chan, H.C.; Zhou, W.L. Cellular mechanisms underlying the laxative effect of flavonol naringenin on rat constipation model. PLoS ONE 2008, 3, 3348. [Google Scholar] [CrossRef] [PubMed]
- Bustos, D.; Ogawa, K.; Pons, S.; Soriano, E.; Bandi, J.C.; Bustos, F.L. Effect of loperamide and bisacodyl on intestinal transit time, fecal weight and short chain fatty acid excretion in the rat. Acta Gastroenterol. Latinoam. 1991, 21, 3–9. [Google Scholar] [PubMed]
- Hughes, S.; Higgs, N.B.; Turnberg, L.A. Loperamide has antisecretory activity in the human jejunum in vivo. Gut 1984, 25, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Sohji, Y.; Kawashima, K.; Shimizu, M. Pharmacological studies of loperamide, an anti-diarrheal agent. II. Effects on peristalsis of the small intestine and colon in guinea pigs (author’s transl). Nihon Yakurigaku Zasshi 1978, 74, 155–163. [Google Scholar] [CrossRef]
- Yamada, K.; Onoda, Y. Comparison of the effects of T-1815, yohimbine and naloxone on mouse colonic propulsion. J. Smooth Muscle Res. 1993, 29, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Niemegeers, C.J.E.; Colpaert, F.C.; Awouters, F.H.L. Pharmacology and antidiarrheal effect of loperamide. Drug Dev. Res. 1981, 1, 1–20. [Google Scholar] [CrossRef]
- Altaparmakov, I.; Wienbeck, M. Local inhibition of myoelectrical activity of human colon by loperamide. Dig. Dis. Sci. 1984, 29, 232–238. [Google Scholar] [CrossRef]
- Hansson, G.C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 2012, 15, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Antoni, L.; Nuding, S.; Wehkamp, J.; Stange, E.F. Intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1165–1179. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, J.W.; Cho, H.R.; Kim, K.Y.; Lee, J.K.; Sohn, J.H.; Ku, S.K. Laxative effects of fermented rice extract in rats with loperamide-induced constipation. Exp. Ther. Med. 2014, 8, 1847–1854. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Lee, M.R.; Park, J.J.; Choi, J.Y.; Song, B.R.; Son, H.J.; Choi, Y.W.; Kim, K.M.; Hong, J.T.; Hwang, D.Y. Quercetin promotes gastrointestinal motility and mucin secretion in loperamide-induced constipation of SD rats through regulation of the mAChRs downstream signal. Pharm. Biol. 2018, 56, 309–317. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contents | NCTR | SPA | Lop | ||
|---|---|---|---|---|---|
| Vehicle | SPA | ||||
| Body weight (g) | 289 ± 17 | 288 ± 7 | 279 ± 11 | 283 ± 12 | |
| Feeding behavior | Food intake (g/day) | 18.7 ± 3.6 | 23.0 ± 1.2 | 22.0 ± 3.28 | 23.8 ± 1.8 |
| Water consumption (ml/day) | 29.4 ± 3.4 | 27.7 ± 3.4 | 26.1 ± 4.1 | 25.4 ± 3.6 | |
| Urine volume (mL/day) | 11.6 ± 5.6 | 18.3 ± 2.1 * | 21.6 ± 2.8 * | 16.0 ± 1.6 # | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.E.; Park, J.W.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Choi, Y.S.; Lee, Y.J.; Lee, H.S.; Hong, J.T.; Hwang, D.Y. Anti-Inflammatory Response and Muscarinic Cholinergic Regulation during the Laxative Effect of Asparagus cochinchinensis in Loperamide-Induced Constipation of SD Rats. Int. J. Mol. Sci. 2019, 20, 946. https://doi.org/10.3390/ijms20040946
Kim JE, Park JW, Kang MJ, Choi HJ, Bae SJ, Choi YS, Lee YJ, Lee HS, Hong JT, Hwang DY. Anti-Inflammatory Response and Muscarinic Cholinergic Regulation during the Laxative Effect of Asparagus cochinchinensis in Loperamide-Induced Constipation of SD Rats. International Journal of Molecular Sciences. 2019; 20(4):946. https://doi.org/10.3390/ijms20040946
Chicago/Turabian StyleKim, Ji Eun, Ji Won Park, Mi Ju Kang, Hyeon Jun Choi, Su Ji Bae, You Sang Choi, Young Ju Lee, Hee Seob Lee, Jin Tae Hong, and Dae Youn Hwang. 2019. "Anti-Inflammatory Response and Muscarinic Cholinergic Regulation during the Laxative Effect of Asparagus cochinchinensis in Loperamide-Induced Constipation of SD Rats" International Journal of Molecular Sciences 20, no. 4: 946. https://doi.org/10.3390/ijms20040946
APA StyleKim, J. E., Park, J. W., Kang, M. J., Choi, H. J., Bae, S. J., Choi, Y. S., Lee, Y. J., Lee, H. S., Hong, J. T., & Hwang, D. Y. (2019). Anti-Inflammatory Response and Muscarinic Cholinergic Regulation during the Laxative Effect of Asparagus cochinchinensis in Loperamide-Induced Constipation of SD Rats. International Journal of Molecular Sciences, 20(4), 946. https://doi.org/10.3390/ijms20040946