Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation?


 ,
, 
Abstract
:1. Introduction
2. Platelet Activation and Platelet Microparticle Production
2.1. Introduction to Primary Haemostasis
2.2. Platelet Activation via Other Glycoprotein and G-protein Coupled Receptors
2.3. Downstream Platelet Intracellular Signalling
2.4. Platelet Microparticles (PMPs)
3. Platelet Function Tests
3.1. Aggregometry
3.2. Flow Cytometry
3.3. Other Platelet Function Tests
4. Role of Polyphenols in Modulating Platelet Aggregation and PMP Production
4.1. Polyphenols
4.2. Impact of Polyphenols on Platelet Aggregation
4.3. Impact of Polyphenols on PMP Production
5. Evaluation of Methods Used for Polyphenol Antiplatelet Studies
6. Conclusions and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADP | Adenosine diphosphate |
| ATP | Adenosine triphosphate |
| C3G | Cyanidin-3-glucoside |
| cAMP | Cyclic adenosine monophosphate |
| COX-1 | Cyclooxygenase-1 |
| DAG | Diacylglycerol |
| GSE | Grape Seed Extract |
| NO | Nitric oxide |
| PAR | Protease-activated receptor |
| PIP2 | Phosphatidylinositol 4,5-bisphosphate |
| PI3K | Phosphatidylinositide-3-kinase |
| PKC | Protein kinase C |
| PLC | Phospholipase C |
| PMP | Platelet microparticle |
| PPP | Platelet poor plasma |
| PRP | Platelet rich plasma |
| TRAP | Thrombin receptor-activating peptide |
| TxA2 | Thromboxane A2 |
| vWF | Von-Willebrand factor |
| 5-HT | 5-hydroxytryptamine |
| VASP | Vasodilator-stimulated phosphoprotein |
References
- Waters, A.M.; Trinh, T.; Chau, M.B.; Moon, L. Latest Statistics on Cardiovascular Disease in Australia. Clin. Exp. Pharm. Physiol. 2013, 40, 347–356. [Google Scholar] [CrossRef]
- Organization World Health. Cardiovascular Diseases (Cvds) Fact. Sheet; Organization World Health: Geneva, Switzerland, 2017. [Google Scholar]
- Garraud, O.; Cognasse, F. Are Platelets Cells? And If Yes, Are They Immune Cells? Front. Immunol. 2015, 6, 70. [Google Scholar] [CrossRef] [Green Version]
- Sharda, A.; Flaumenhaft, R. The Life Cycle of Platelet Granules. Research 2018, 7, 236. [Google Scholar] [CrossRef]
- Zaldivia, M.T.K.; McFadyen, J.D.; Lim, B.; Wang, X.; Peter, K. Platelet-Derived Microvesicles in Cardiovascular Diseases. Front. Cardiovasc. Med. 2017, 4, 74. [Google Scholar] [CrossRef] [Green Version]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Gremmel, T.; Koppensteiner, R.; Panze, S. Comparison of Aggregometry with Flow Cytometry for the Assessment of Agonists -Induced Platelet Reactivity in Patients on Dual Antiplatelet Therapy. PLoS ONE 2015, 10, e0129666. [Google Scholar] [CrossRef] [Green Version]
- Sambu, N.; Curzen, N. Monitoring the Effectiveness of Antiplatelet Therapy: Opportunities and Limitations. Br. J. Clin. Pharmacol. 2011, 72, 683–696. [Google Scholar] [CrossRef]
- Damman, P.; Kuijt, R.; Stefan, K.J. P2y12 Platelet Inhibition in Clinical Practice. J. Thromb. Thrombolysis 2012, 33, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Dyszkiewicz-Korpanty, A.; Olteanu, E.P.F.; Sarode, R. Clopidogrel Anti-Platelet Effect: An Evaluation by Optical Aggregometry, Impedance Aggregometry, and the Platelet Function Analyzer (Pfa-100). Platelets 2007, 18, 491–496. [Google Scholar] [CrossRef]
- Santhakumar, B.A.; Bulmer, A.C.; Singh, I. A Review of the Mechanisms and Effectiveness of Dietary Polyphenols in Reducing Oxidative Stress and Thrombotic Risk. J. Hum. Nutr. Diet. 2014, 27, 1–21. [Google Scholar] [CrossRef]
- May, J.A.; Heptinstall, A.T.C.; Hawkey, C.J.P. Responses to Several Agonists and Combinations of Agonists in Whole Blood: A Placebo Controlled Comparison of the Effects of a Once Daily Dose of Plain Aspirin 300 Mg, Plain Aspirin 75 Mg and Enteric Coated Aspirin 300 Mg, in Man. Thromb. Res. 1997, 88, 183–192. [Google Scholar] [CrossRef]
- Espinosa, P.; Enma, V.; John Murad, P.; Fadi Khasawneh, T. Aspirin: Pharmacology and Clinical Applications. Thrombosis 2012, 2012, 173124. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Reddy, V.; Singh, R.S.; Bhatt, G. Aspirin and Clopidogrel Hyporesponsiveness and Nonresponsiveness in Patients with Coronary Artery Stenting. Vasc. Health Risk Manag. 2009, 5, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.H.; Sim, R.Y.; Goh, J.I.P.; Han, J.Y. Platelet Activation: The Mechanisms and Potential Biomarkers. Biomed. Res. Int. 2016, 2016, 9060143. [Google Scholar] [CrossRef] [Green Version]
- Poulter, N.S.; Pollitt, D.M.; Owen, E.E.; Gardiner, R.K.; Andrews, H.; Shimizu, D.; Ishikawa, D.; Bihan, R.W.; Farndale, M.; Moroi, S.P.W.; et al. Clustering of Glycoprotein vi (Gpvi) Dimers Upon Adhesion to Collagen as a Mechanism to Regulate Gpvi Signaling in Platelets. J. Thromb. Haemost. 2017, 15, 549–564. [Google Scholar] [CrossRef] [Green Version]
- Feghhi, S.; Munday, W.; Tooley, S.R.; Fura, J.; Kulman, J.; Nathan, J.S. Glycoprotein Ib-Ix-V Complex Transmits Cytoskeletal Forces That Enhance Platelet Adhesion. Biophys. J. 2016, 111, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Hechler, B.; Gachet, C. P2 Receptors and Platelet Function. Purinergic Signal. 2011, 7, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Nieman, M.T. Protease-Activated Receptors in Hemostasis. Blood 2016, 128, 169–177. [Google Scholar] [CrossRef] [Green Version]
- De Candia, E. Mechanisms of Platelet Activation by Thrombin: A Short History. Thromb. Res. 2012, 129, 250–256. [Google Scholar] [CrossRef]
- Saboor, M.; Ayub, Q.; Samina Ilyas, M. Platelet Receptors; an Instrumental of Platelet Physiology. Pak. J. Med. Sci. 2013, 29, 891–896. [Google Scholar] [CrossRef]
- Rivera, J.; Lozano, M.L.; Navarro-Núñez, L.; Vicente, V. Platelet Receptors and Signaling in the Dynamics of Thrombus Formation. Haematologica 2009, 94, 700–711. [Google Scholar] [CrossRef]
- Bye, P.A.; Unsworth, A.J.; Gibbins, J.M. Platelet Signaling: A Complex Interplay between Inhibitory and Activatory Networks. J. Thromb. Haemost. 2016, 14, 918–930. [Google Scholar] [CrossRef] [Green Version]
- Sangkuhl, K.; Shuldiner, T.; Russ, B.A. Platelet Aggregation Pathway. Pharm. Genom. 2011, 21, 516–521. [Google Scholar] [CrossRef] [Green Version]
- Bledzka, K.; Susan, S.S.; Edward, P.F. Integrin Aiibβ3: From Discovery to Efficacious Therapeutic Target. Circ. Res. 2013, 112, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Badimon, L.R.; Suades, E.; Fuentes, I.; Palomo, T.P. Role of Platelet-Derived Microvesicles as Crosstalk Mediators in Atherothrombosis and Future Pharmacology Targets: A Link between Inflammation, Atherosclerosis, and Thrombosis. Front. Pharm. 2016, 7, 293. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Ghassan, K.S. Role of Shear Stress and Stretch in Vascular Mechanobiology. J. R. Soc. Interface 2011, 8, 1379–1385. [Google Scholar] [CrossRef] [Green Version]
- Dean William, L.; Menq, J.; Lee, T.; Cummins, D.; David, W.P. Proteomic and Functional Characterisation of Platelet Microparticle Size Classes. Thromb. Haemost. 2009, 102, 711–718. [Google Scholar] [CrossRef]
- Mumford, A.D.; Gachet, P.; Gresele, P.; Noris, P.H.; Mezza, D. A Review of Platelet Secretion Assays for the Diagnosis of Inherited Platelet Secretion Disorders. Thromb. Haemost. 2015, 114, 14–25. [Google Scholar] [CrossRef]
- Paniccia, R.; Priora, A.A.L.; Abbate, R. Platelet Function Tests: A Comparative Review. Vasc. Health Risk Manag. 2015, 11, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Pai, M.; Moffat, Y.L.; Hayward, C. Diagnostic Usefulness of a Lumi-Aggregometer Adenosine Triphosphate Release Assay for the Assessment of Platelet Function Disorders. Am. J. Clin. Pathol. 2011, 136, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Koltai, K.; Kesmarky, G.; Feher, G.; Tibold, A.; Toth, K. Platelet Aggregometry Testing: Molecular Mechanisms, Techniques and Clinical Implications. Int. J. Mol. Sci. 2017, 18, 1803. [Google Scholar] [CrossRef] [PubMed]
- Dubois, C.; Gainor, B.; Furie, B. Thrombin-Initiated Platelet Activation in Vivo Is Vwf Independent During Thrombus Formation in a Laser Injury Model. J. Clin. Investig. 2007, 117, 953–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Mogami, Y.; Murakami, T.; Nakamura, N.; Kanayama, H.K. UraReal-Time Analysis of Platelet Aggregation and Procoagulant Activity During Thrombus Formation in Vivo. Pflug. Arch. 2008, 456, 1239–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, C.; Emerson, M. Assessment of Platelet Aggregation Responses in Vivo in the Mouse. Methods Mol. Biol. 2012, 788, 21–28. [Google Scholar]
- Brown, M.; Wittwer, C. Flow Cytometry: Principles and Clinical Applications in Hematology. Clin. Chem. 2000, 46, 1221. [Google Scholar]
- Saboor, M.; Moinuddin, M.; Ilyas, S. New Horizons in Platelets Flow Cytometry. Malays. J. Med. Sci. 2013, 20, 62–66. [Google Scholar]
- Lu, Q.; Richard, M.A. Comparison of Two Platelet Activation Markers Using Flow Cytometry after in Vitro Shear Stress Exposure of Whole Human Blood. Artif. Organs 2011, 35, 137–144. [Google Scholar] [CrossRef]
- Kailashiya, J. Platelet-Derived Microparticles Analysis: Techniques, Challenges and Recommendations. Anal. Biochem. 2018, 546, 78–85. [Google Scholar] [CrossRef]
- Siljander, P.R.M. Platelet-Derived Microparticles—An Updated Perspective. Thromb. Res. 2011, 127, S30–S33. [Google Scholar] [CrossRef]
- Erdbrugger, U.C.K.; Rudy, M.E.; Etter, K.A.; Dryden, M.; Yeager, A.L.; Klibanov, J.L. Imaging Flow Cytometry Elucidates Limitations of Microparticle Analysis by Conventional Flow Cytometry. Cytom. A 2014, 85, 756–770. [Google Scholar] [CrossRef]
- Rywaniak, J.; Luzak, A.; Podsedek, D.; Dudzinska, M.R.; Watala, C. Comparison of Cytotoxic and Anti-Platelet Activities of Polyphenolic Extracts from Arnica Montana Flowers and Juglans Regia Husks. Platelets 2015, 26, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Bijak, M.; Sut, A.; Kosiorek, A.; Saluk-Bijak, J.; Golanski, J. Dual Anticoagulant/Antiplatelet Activity of Polyphenolic Grape Seeds Extract. Nutrients 2019, 11, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijak, M.; Bijak, M.; Szelenberger, R.; Dziedzic, A.; Saluk-Bijak, J. Inhibitory Effect of Flavonolignans on the P2y12 Pathway in Blood Platelets. Molecules 2018, 23, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.H.; Rice, A.S.; Garrett, K.; Stein, S.F. Gender, Race and Diet Affect Platelet Function Tests in Normal Subjects, Contributing to a High Rate of Abnormal Results. Br. J. Haematol. 2014, 165, 842–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludovici, V.; Barthelmes, M.P.; Nagele, A.J.F.; Suda, I. Polyphenols: Anti-Platelet Nutraceutical? Curr. Pharm. Des. 2018, 24, 146–157. [Google Scholar] [CrossRef]
- Santhakumar, B.; Battino, A.M.; Alvarez-Suarez, J.M. Dietary Polyphenols: Structures, Bioavailability and Protective Effects against Atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef]
- Bowey, E.; Adlercreutz, H.; Rowland, I. Metabolism of Isoflavones and Lignans by the Gut Microflora: A Study in Germ-Free and Human Flora Associated Rats. Food Chem. Toxicol. 2003, 41, 631–636. [Google Scholar] [CrossRef]
- Aura, A.M.; Williamson, K.M.O.; Santos-Buelga, C. In Vitro Metabolism of Anthocyanins by Human Gut Microflora. Eur. J. Nutr. 2005, 44, 133–142. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Richard, F.A. Characterization of Protein–Polyphenol Interactions. Trends Food Sci. Technol. 2004, 15, 186–190. [Google Scholar] [CrossRef]
- Goszcz, K.G.G.; Duthie, D.; Stewart, S.J.; Leslie, I.L.M. Bioactive Polyphenols and Cardiovascular Disease: Chemical Antagonists, Pharmacological Agents or Xenobiotics That Drive an Adaptive Response? Br. J. Pharm. 2017, 174, 1209–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arts, I.C.; Hollman, P.C. Polyphenols and Disease Risk in Epidemiologic Studies. Am. J. Clin. Nutr. 2005, 81, 317s–325s. [Google Scholar] [CrossRef] [Green Version]
- Wright, B.; Moraes, C.; Kemp, W.M.; Jonathan, M.G. A Structural Basis for the Inhibition of Collagen-Stimulated Platelet Function by Quercetin and Structurally Related Flavonoids. Br. J. Pharmacol. 2010, 159, 1312–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.; Jawad, M.; Kamal, M.A.; Baldi, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Evidence and Prospective of Plant Derived Flavonoids as Antiplatelet Agents: Strong Candidates to Be Drugs of Future. Food Chem. Toxicol. 2018, 119, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.; Quinn, H.M.; Mok, R.J.; Southgate, A.H.; Turner, D.; Li, A.J.; Sinclair, J.A.H. The Effect of Exercise and Training Status on Platelet Activation: Do Cocoa Polyphenols Play a Role? Platelets 2006, 17, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Rimbach, G.; Mona, M.; Jennifer, M.; Anika, E.W. Polyphenols from Cocoa and Vascular Health—A Critical Review. Int. J. Mol. Sci. 2009, 10, 4290–4309. [Google Scholar] [CrossRef]
- Ostertag, L.M.; O’Kennedy, N.; Kroon, P.A.; Duthie, G.G.; de Roos, B. Impact of Dietary Polyphenols on Human Platelet Function—A Critical Review of Controlled Dietary Intervention Studies. Mol. Nutr. Food Res. 2010, 54, 60–81. [Google Scholar] [CrossRef]
- Singh, I.; Mok, M.; Christensen, A.M.; Turner, A.H.; Hawley, J.A. The Effects of Polyphenols in Olive Leaves on Platelet Function. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Vogel, P.; Kasper Machado, I.; Garavaglia, J.; Zani, V.T.; de Souza, D.; Morelo Dal Bosco, S. Polyphenols Benefits of Olive Leaf (Olea europaea L) to Human Health. Nutr. Hosp. 2014, 31, 1427–1433. [Google Scholar]
- Fuentes, E.; Julio, C.; Marcelo, A.; Armando, R.; Iván, P. Chlorogenic Acid Inhibits Human Platelet Activation and Thrombus Formation. PLoS ONE 2014, 9, e90699. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Chen, W.F.; Chen, K.H.; Hsieh, C.Y.; Chou, D.S.; Lin, L.J.; Sheu, J.R.; Chang, C.C. Novel Bioactivity of Ellagic Acid in Inhibiting Human Platelet Activation. Evid. Based Complement. Altern. Med. 2013, 2013, 595128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.H.; Deng, X.J.; Chen, Y.Q.; Ya, F.L.; Zhang, X.D.; Song, F.; Li, D.; Yang, Y. Anthocyanin Cyanidin-3-Glucoside Attenuates Platelet Granule Release in Mice Fed High-Fat Diets. J. Nutr. Sci. Vitam. 2017, 63, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Chen, Y.; Adili, R.; McKeown, T.; Chen, P.; Zhu, G.; Li, D.; Ling, W.; Ni, H.; Yang, Y. Plant-Based Food Cyanidin-3-Glucoside Modulates Human Platelet Glycoprotein Vi Signaling and Inhibits Platelet Activation and Thrombus Formation. J. Nutr. 2017, 147, 1917–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, W.H.; Cheng, Q.X.; Ma, J.; Wang, T. Red and Black Rice Decrease Atherosclerotic Plaque Formation and Increase Antioxidant Status in Rabbits. J. Nutr. 2001, 131, 1421–1426. [Google Scholar] [CrossRef]
- Callcott, E.T.; Thompson, K.; Oli, P.; Blanchard, C.L.; Santhakumar, A.B. Coloured Rice-Derived Polyphenols Reduce Lipid Peroxidation and Pro-Inflammatory Cytokines Ex Vivo. Food Funct. 2018, 9, 5169–5175. [Google Scholar] [CrossRef]
- Callcott, E.T.; Blanchard, C.L.; Oli, P.; Santhakumar, A.B. Pigmented Rice-Derived Phenolic Compounds Reduce Biomarkers of Oxidative Stress and Inflammation in Human Umbilical Vein Endothelial Cells. Mol. Nutr. Food Res. 2018, 62, e1800840. [Google Scholar] [CrossRef]
- De la Torre, R. Bioavailability of Olive Oil Phenolic Compounds in Humans. Inflammopharmacology 2008, 16, 245–247. [Google Scholar] [CrossRef]
- Correa, J.A.; López-Villodres, J.A.; Asensi, R.; Espartero, J.L.; Rodríguez-Gutiérez, G.; De La Cruz, J.P. Virgin Olive Oil Polyphenol Hydroxytyrosol Acetate Inhibits in Vitro Platelet Aggregation in Human Whole Blood: Comparison with Hydroxytyrosol and Acetylsalicylic Acid. Br. J. Nutr. 2009, 101, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, D.; Mazumder, S.; Sinha, A.K. The Role of Inhibition of Nitric Oxide Synthesis in the Aggregation of Platelets Due to the Stimulated Production of Thromboxane A2. Blood Coagul. Fibrinolysis 2014, 25, 585–591. [Google Scholar] [CrossRef]
- Bojic, M.; Antolic, A.; Tomicic, M.; Tomicic, Z.; Males, Z. Propolis Ethanolic Extracts Reduce Adenosine Diphosphate Induced Platelet Aggregation Determined on Whole Blood. Nutr. J. 2018, 17, 52. [Google Scholar] [CrossRef] [Green Version]
- Faggio, C.; Sureda, A.; Morabito, S.; Sanches-Silva, A.; Mocan, A.; Nabavi, S.F.; Nabavi, S.M. Flavonoids and Platelet Aggregation: A Brief Review. Eur. J. Pharmacol. 2017, 807, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Bojić, M.; Debeljak, Z.; Tomičić, M.; Medić-Šarić, M.; Tomić, S. Evaluation of Antiaggregatory Activity of Flavonoid Aglycone Series. Nutr. J. 2011, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; McGeoch, S.C.; Johnstone, A.M.; Holtrop, G.; Sneddon, A.A.; MacRury, S.M.; Megson, I.L.; Pearson, D.W.; Abraham, P.; De Roos, B.; et al. Platelet-Derived Microparticle Count and Surface Molecule Expression Differ between Subjects with and without Type 2 Diabetes, Independently of Obesity Status. J. Thromb. Thrombolysis 2014, 37, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; McGeoch, S.C.; Megson, I.L.; MacRury, S.M.; Johnstone, A.M.; Abraham, P.; Pearson, D.W.; de Roos, B.; Holtrop, G.; O’Kennedy, N.; et al. Oat-Enriched Diet Reduces Inflammatory Status Assessed by Circulating Cell-Derived Microparticle Concentrations in Type 2 Diabetes. Mol. Nutr. Food Res. 2014, 58, 1322–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, L.; Thondre, P.S.; Henry, C.J.K. Oat-Based Breakfast Cereals Are a Rich Source of Polyphenols and High in Antioxidant Potential. J. Food Compos. Anal. 2011, 24, 929–934. [Google Scholar] [CrossRef]
- Sur, R.; Nigam, A.; Grote, D.; Liebel, F.; Southall, M.D. Avenanthramides, Polyphenols from Oats, Exhibit Anti-Inflammatory and Anti-Itch Activity. Arch. Dermatol. Res. 2008, 300, 569–574. [Google Scholar] [CrossRef]
- Tripathi, V.; Mohd, A.S.; Ashraf, T. Avenanthramides of Oats: Medicinal Importance and Future Perspectives. Pharmacogn. Rev. 2018, 12, 66–71. [Google Scholar] [CrossRef]
- Hosseini, B.; Saedisomeolia, A.; Wood, L.G.; Yaseri, M.; Tavasoli, S. Effects of Pomegranate Extract Supplementation on Inflammation in Overweight and Obese Individuals: A Randomized Controlled Clinical Trial. Complement. Ther. Clin. Pract. 2016, 22, 44–50. [Google Scholar] [CrossRef]
- Chang, S.-S.; Viola, S.Y.; Lee, Y.T.; Li, C. Gallic Acid Attenuates Platelet Activation and Platelet-Leukocyte Aggregation: Involving Pathways of Akt and Gsk3β. Evid. Based Complement. Altern. Med. 2012, 2012, 683872. [Google Scholar] [CrossRef] [Green Version]
- Olas, B.; Wachowicz, B.; Stochmal, A.; Oleszek, W. The Polyphenol-Rich Extract from Grape Seeds Inhibits Platelet Signaling Pathways Triggered by Both Proteolytic and Non-Proteolytic Agonists. Platelets 2012, 23, 282–289. [Google Scholar] [CrossRef]
- Veluri, R.; Singh, R.P.; Liu, Z.; Thompson, J.A.; Agarwal, R.; Agarwal, C. Fractionation of Grape Seed Extract and Identification of Gallic Acid as One of the Major Active Constituents Causing Growth Inhibition and Apoptotic Death of Du145 Human Prostate Carcinoma Cells. Carcinogenesis 2006, 27, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.J.; Yu, J.E.; Pohorly, J.E.; Kakuda, Y. Polyphenolics in Grape Seeds-Biochemistry and Functionality. J. Med. Food 2003, 6, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Ravishankar, D.; Albadawi, D.A.I.; Chaggar, V.; Patra, P.H.; Williams, H.F.; Salamah, M.; Vaiyapuri, R.; Dash, P.R.; Patel, K.; Watson, K.A.; et al. Isorhapontigenin, a Resveratrol Analogue Selectively Inhibits Adp-Stimulated Platelet Activation. Eur. J. Pharmacol. 2019, 27, 1726. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Huang, Y.; Zou, J.; Cao, K.; Xu, Y.; Wu, J.M. Effects of red wine and wine polyphenol resveratrol on platelet aggregation in vivo and in vitro. Int. J. Mol. Med. 2002, 9, 77–79. [Google Scholar] [CrossRef] [PubMed]
- López Andrés, N.; Thornton, C.L.; Andriantsitohaina, R. Increased Microparticle Production and Impaired Microvascular Endothelial Function in Aldosterone-Salt-Treated Rats: Protective Effects of Polyphenols. PLoS ONE 2012, 7, e39235. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.-Y.; Xu, R.; Zhang, S.; Qiao, Q.; Shen, L.; Li, M.; Xu, D.; Wang, Z. Alteration of Circulatory Platelet Microparticles and Endothelial Microparticles in Patients with Chronic Kidney Disease. Int. J. Clin. Exp. Med. 2015, 8, 16704–16708. [Google Scholar]
- Giacomazzi, A.; Degan, M.; Calabria, S.; Meneguzzi, A.; Minuz, P. Antiplatelet Agents Inhibit the Generation of Platelet-Derived Microparticles. Front. Pharmacol. 2016, 7, 314. [Google Scholar] [CrossRef] [Green Version]
- Michal, B.; Angela, D.; Saluk-Bijak, J. Flavonolignans Reduce the Response of Blood Platelet to Collagen. Int. J. Biol. Macromol. 2018, 106, 878–884. [Google Scholar]
- Bijak, M.; Saluk-Bijak, J. Flavonolignans Inhibit the Arachidonic Acid Pathway in Blood Platelets. BMC Complement. Altern. Med. 2017, 17, 396. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, E.; Vigh, E.; Molnár, V.; Kenyeres, P.; Fehér, G.; Késmárky, G.; Tóth, K.; Garai, J. Thrombosis Preventive Potential of Chicory Coffee Consumption: A Clinical Study. Phytother. Res. 2011, 25, 744–748. [Google Scholar] [CrossRef] [Green Version]
- Nickel, T.; Lackermair, K.; Scherr, J.; Calatzis, A.; Vogeser, M.; Hanssen, H.; Waidhauser, G.; Schönermark, U.; Methe, H.; Horster, S. Influence of High Polyphenol Beverage on Stress-Induced Platelet Activation. J. Nutr. Health Aging 2016, 20, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Graaf, Y.; Slettenaar, M.; de Groot, E.; Wright, C. Consumption of a Polyphenol-Rich Grape-Wine Extract Lowers Ambulatory Blood Pressure in Mildly Hypertensive Subjects. Nutrients 2015, 7, 3138–3153. [Google Scholar]
- Ras, R.T.; Zock, P.L.; Zebregs, Y.E.; Johnston, N.R.; Webb, D.J.; Draijer, R. Effect of Polyphenol-Rich Grape Seed Extract on Ambulatory Blood Pressure in Subjects with Pre- and Stage I Hypertension. Br. J. Nutr. 2013, 110, 2234–2241. [Google Scholar] [CrossRef] [PubMed]
- Flammer, A.J.; Sudano, I.; Wolfrum, M.; Thomas, R.; Enseleit, F.; Périat, D.; Kaiser, P.; Hirt, A.; Hermann, M.; Serafini, M. Cardiovascular Effects of Flavanol-Rich Chocolate in Patients with Heart Failure. Eur. Heart J. 2011, 33, 2172–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loo, B.-M.; Erlund, I.; Koli, R.; Puukka, P.; Hellström, J.; Wähälä, K.; Mattila, P.; Jula, A. Consumption of Chokeberry (Aronia mitschurinii) Products Modestly Lowered Blood Pressure and Reduced Low-Grade Inflammation in Patients with Mildly Elevated Blood Pressure. Nutr. Res. 2016, 36, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Bommannan, A.; Kundur, S.A.R.; Fanning, K.; Netzel, M.; Stanley, R.; Singh, I. Consumption of Anthocyanin-Rich Queen Garnet Plum Juice Reduces Platelet Activation Related Thrombogenesis in Healthy Volunteers. J. Funct. Foods 2015, 12, 11–22. [Google Scholar]
- Ostertag, L.M.; O’Kennedy, N.G.W.; Horgan, P.A.; Kroon, G.G.D.; de Roos, B. In Vitro Anti-Platelet Effects of Simple Plant-Derived Phenolic Compounds Are Only Found at High, Non-Physiological Concentrations. Mol. Nutr. Food Res. 2011, 55, 1624–1636. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and Bioefficacy of Polyphenols in Humans. I. Review of 97 Bioavailability Studies. Am. J. Clin. Nutr. 2005, 81, 230s–242s. [Google Scholar] [CrossRef] [Green Version]
- Scholz, S.; Williamson, G. Interactions Affecting the Bioavailability of Dietary Polyphenols in Vivo. Int. J. Vitam. Nutr. Res. 2007, 77, 224–235. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef]
- Santhakumar, B.A.; Stanley, R.; Singh, I. The Ex Vivo Antiplatelet Activation Potential of Fruit Phenolic Metabolite Hippuric Acid. Food Funct. 2015, 6, 2679–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stainer, A.; Bye, A.; Unsworth, L.; Holbrook, M.T.; Jonathan, M.G. The Metabolites of the Dietary Flavonoid Quercetin Possess Potent Antithrombotic Activity, and Interact with Aspirin to Enhance Antiplatelet Effects. TH Open Companion J. Thromb. Haemost. 2019, 3, e244–e258. [Google Scholar] [CrossRef] [PubMed]
- Basheer, L.; Zohar, K. Interactions between Cyp3a4 and Dietary Polyphenols. Oxidative Med. Cell. Longev. 2015, 15, 8540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.H.; Ito, R.; Ohnishi, T. HataInhibitory Effects of Polyphenols on Human Cytochrome P450 3a4 and 2c9 Activity. Food Chem. Toxicol. 2010, 48, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Are Polyphenols Antioxidants or Pro-Oxidants? What Do We Learn from Cell Culture and in Vivo Studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.; Sang, D.S.; Yang, C.S. Possible Controversy over Dietary Polyphenols: Benefits Vs Risks. Chem. Res. Toxicol. 2007, 20, 583–585. [Google Scholar] [CrossRef]
- Nardini, M.; Natella, F.; Scaccini, C. Role of Dietary Polyphenols in Platelet Aggregation. A Review of the Supplementation Studies. Platelets 2007, 18, 224–243. [Google Scholar] [CrossRef]
- Marx, W.; Kelly, S.; Marshall, S.; Nakos, K.C.; Itsiopoulos, C. The Effect of Polyphenol-Rich Interventions on Cardiovascular Risk Factors in Haemodialysis: A Systematic Review and Meta-Analysis. Nutrients 2017, 9, 1345. [Google Scholar] [CrossRef] [Green Version]
- Cointe, S.; Judicone, C.S.; Robert, M.J.; Mooberry, P.; Poncelet, M.; Wauben, R.; Nieuwland, N.S.; Key, F.D.; Lacroix, R. Standardization of Microparticle Enumeration across Different Flow Cytometry Platforms: Results of a Multicenter Collaborative Workshop. J. Thromb. Haemost. 2017, 15, 187–193. [Google Scholar] [CrossRef]
- Recabarren-Leiva, D.; Alarcón, M. Standardization of a Fast and Effective Method for the Generation and Detection of Platelet-Derived Microparticles by a Flow Cytometer. Immunol. Lett. 2018, 79–84. [Google Scholar] [CrossRef]
- Huskens, D.; Sang, Y.; Konings, J.; van der Vorm, L.; de Laat, B.; Kelchtermans, H.; Roest, M. Standardization and Reference Ranges for Whole Blood Platelet Function Measurements Using a Flow Cytometric Platelet Activation Test. PLoS ONE 2018, 13, e0192079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikora, J.; Markowicz-Piasecka, M.; Broncel, M.; Mikiciuk-Olasik, E. Extract of Aronia Melanocarpa-Modified Hemostasis: In Vitro Studies. Eur. J. Nutr. 2014, 53, 1493–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]


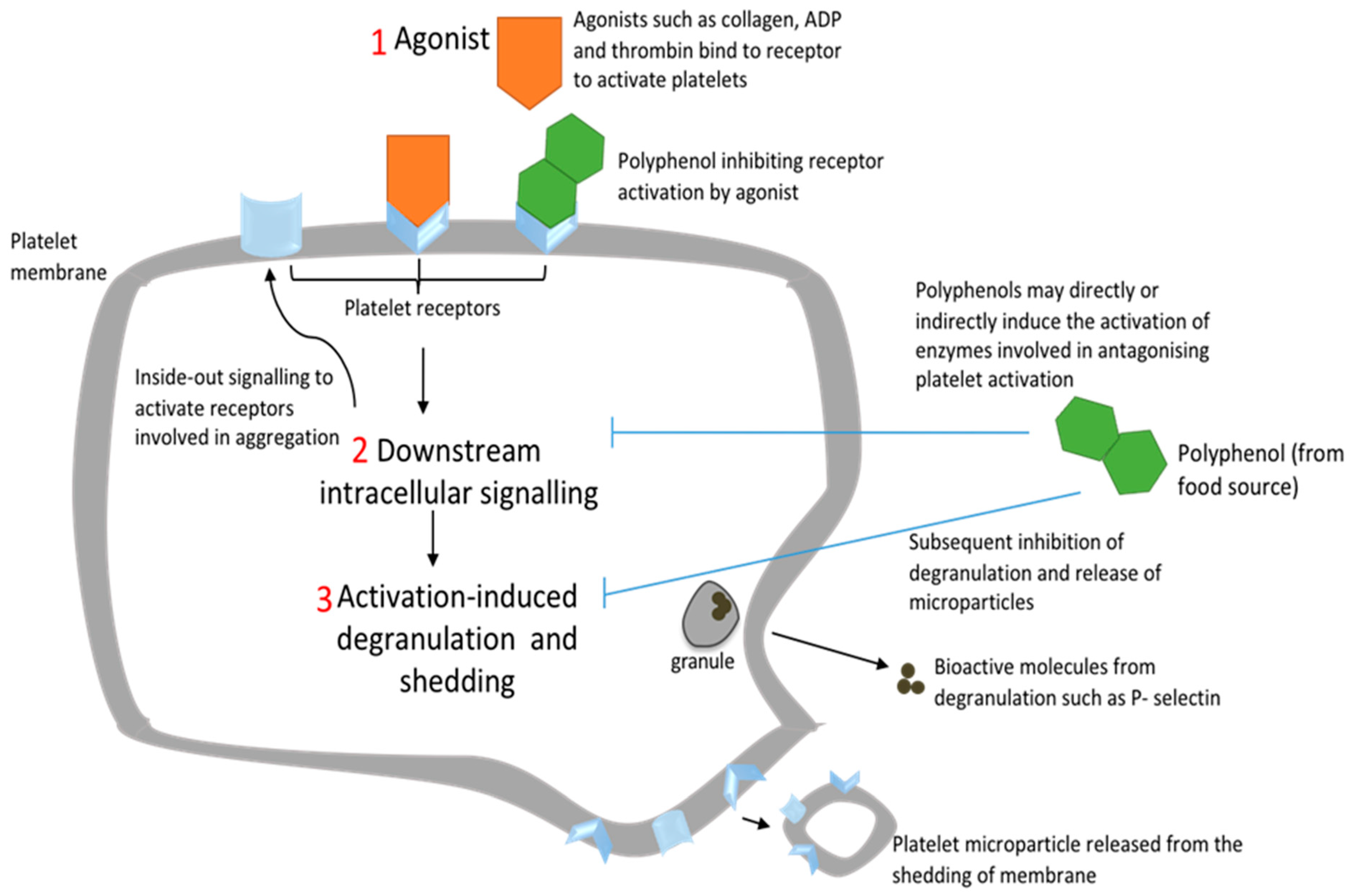{kind=link}
| Design Model | Polyphenol/Source | Population | Study Description | Aggregometry (Agg) / Flow Cytometry (Flow) | Findings | Reference |
|---|---|---|---|---|---|---|
| In vitro | Grape seed extract | 30 healthy subjects | ADP-induced aggregation and VASP phosphorylation to evaluate inhibition of GSE (7.5 µg mL−1 and 15 µg mL−1) | Both | Reduced ADP-induced platelet aggregation and platelet reactivity index (by increasing VASP phosphorylation) | [43] |
| In vitro | Olive leaves | 11 healthy male subjects | Randomized single blind study. Evaluation of inhibition of platelet aggregation and ATP release by olive leaf extract polyphenols | Agg | Significant dose-dependent inhibition of aggregation and ATP release | [59] |
| In vitro | Grape seed extract | 5 healthy subjects | TRAP and thrombin-induced activation of gel filtered platelets to compare antiplatelet property of GSE (1.25–50 µg mL−1) with pure resveratrol | Both | Inhibition of TRAP and thrombin-induced P-selectin expression, PMP formation, platelet aggregation, and superoxide anion radicals generated from platelets in a dose-dependent manner | [81] |
| In vitro | Propolis | 10 healthy subjects | ADP-induced platelet aggregation to evaluate inhibition 21 propolis polyphenolic extracts | Agg | Most potent inhibitor of aggregation contained luteolin, apigenin, and chrysin | [71] |
| In vitro | Extracts of the Arnica montana L. flower head and walnut husks (Juglans regia L) | 55 healthy subjects | Inhibition of A. montana or J. regia polyphenol extracts (7.5 and 15 µg mL−1) using: (a) Arachidonic acid-, collagen-, and ADP-induced aggregation. (b) VASP phosphorylation and platelet surface marker expression. | Both | Significant inhibition of ADP-induced aggregation but not collagen or arachidonic acid. No significant effect on P-selectin or GPIIb/IIIa expression. Significant decrease in VASP phosphorylation | [42] |
| In vitro | Virgin olive oil polyphenols | 6 healthy subjects | Single-blind study. Anti-aggregation effect of hydroxytyrosol acetate (HT-AC) compared with hydroxytyrosol (HT) and aspirin in whole blood and PRP | Agg | HT-AC anti-aggregation effect higher than HT but similar to aspirin | [70] |
| In vitro and in vivo | Chlorogenic acid | 6 healthy subjects (18 mice also used for in vivo assays) | Activation of washed platelets by multiple agonist to evaluate chlorogenic acid (0.1 to 1 mmoL/L) effect on aggregation, secretion, platelet surface marker expression, cAMP levels, and PKA activation. Chlorogenic acid inhibition of thrombus formation assessed in vivo | Both | Dose dependent inhibition of ADP-, collagen-, arachidonic acid-, and TRAP-6-induced aggregation, ATP release P-selectin expression, and GP IIb/IIIa expression. Increased intraplatelet cAMP and PKA activation. Inhibition of in vivo thrombus formation | [61] |
| In vivo | Anthocyanin C3G from purified black rice | 60 male mice | Mice randomly allocated to one of the 3 groups with 20 subjects for each group: control group (normal diet), high-fat diet (HFD group), or an HFD supplemented with C3G | Both | Decreased platelet activation, serum lipid levels, and inhibits platelet ATP release | [63] |
| In vivo | Red wine polyphenols (Provinols™) | 149 (uniephrectomized male Sprague-Dawley rats) | Rats were randomly grouped based on being treated with or without aldosterone-salt, with or without Provinols (20 mg/kg/day) or spironolactone (30 mg/kg/day) for 4 weeks | Flow (including erythroaggregometer using in vitro shear stress-induced platelet activation) | Provinols decreased circulating microparticles independent of shear stress or mineralocorticoid receptor activation | [84] |
| In vivo | Cocoa | 16 healthy male subjects | Double-blind, crossover study. Placebo-controlled. Eight trained and untrained subjects randomly assigned to receive placebo or cocoa polyphenol supplements (236 mg/day) over a week and then afterwards subjected to one hour of exercise | Both | No change in collagen induced aggregation post exercise. ATP release higher post exercise in both trained and untrained groups. Cocoa supplementation administered over a week did not normalize platelet activity after exercise | [56] |
| In vivo | Chicory coffee | 27 healthy subjects | 300 mL chicory coffee every day for 1 week | Agg | Variable effects on platelet aggregation depending on the agonists used | [91] |
| In vivo | High polyphenol beverage | 103 healthy athletes | Randomized, double-blind study. Group 1 received a polyphenol-rich beverage, Group 2 a placebo. Samples were collected three weeks before, one day before, immediately, as well as 24 h and 72 h, after a marathon run | Agg | Control group demonstrated a 2.2-fold increase in platelet aggregation after marathon completion. No increase in platelet aggregation in polyphenol-rich beverage group | [92] |
| In vivo | Polyphenol-rich grape wine | 60 untreated, mildly hypertensive subjects | Double-blind placebo-controlled crossover study. Grape juice extract; grape and wine extract each for 4 weeks including a 2-week run-in period | Agg | No effect on ADP, collagen, or epinephrine induced platelet aggregation | [93] |
| In vivo | Polyphenol-rich grape seed extract | 35 untreated subjects with pre and stage 1 hypertension | Double-blind, placebo-controlled, randomized, parallel-group intervention with 300 mg/day grape seed extract capsule. Eight-week intervention period. | Agg | Did not affect platelet aggregation | [94] |
| In vivo | Flavanol-rich chocolate | 20 patients with congestive heart failure | Double-blind, randomized placebo-controlled trial. Supplementation – (2 h after ingestion of a chocolate bar) and long term (4 weeks, two chocolate bars/day). | Agg | Platelet adhesion significantly decreased 2 h after flavanol-rich chocolate ingestion. No effect post 2- and 4-week supplementation | [95] |
| In vivo | Chokeberry (Aronia mitschurinii) products | 38 patients with untreated mild hypertension | 16-week single blinded crossover trial. Cold-pressed 100% chokeberry juice (300 mL/d) and oven-dried chokeberry powder (3 g/d), or placebo for 8 weeks without washout. | Agg | No change in platelet aggregation | [96] |
| In vivo | Oats | 22 type 2 diabetes subjects | Randomized crossover involving 8-week intervention with either oat enriched diet (OAT) or a standard dietary (SDA) advice diet. Preintervention habitual (HAB) intakes were used to compare responses. | Flow | Decrease in tissue factor-activated platelets (CD142) after OAT than HAB or SDA. Decrease in tissue factor positive PMPs and fibrinogen-positive PMPs with OAT intervention. | [75] |
| In vivo | Anthocyanin rich beverage | 21 sedentary subjects | Double-blind placebo controlled. Queen garnet plum juice (200 mL/day) were consumed for 28 days | Both | Reduced ADP, collagen, and arachidonic acid-induced platelet aggregation. Reduced P-selectin expression | [97] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ed Nignpense, B.; Chinkwo, K.A.; Blanchard, C.L.; Santhakumar, A.B. Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation? Int. J. Mol. Sci. 2020, 21, 146. https://doi.org/10.3390/ijms21010146
Ed Nignpense B, Chinkwo KA, Blanchard CL, Santhakumar AB. Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation? International Journal of Molecular Sciences. 2020; 21(1):146. https://doi.org/10.3390/ijms21010146
Chicago/Turabian StyleEd Nignpense, Borkwei, Kenneth A. Chinkwo, Christopher L. Blanchard, and Abishek B. Santhakumar. 2020. "Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation?" International Journal of Molecular Sciences 21, no. 1: 146. https://doi.org/10.3390/ijms21010146
APA StyleEd Nignpense, B., Chinkwo, K. A., Blanchard, C. L., & Santhakumar, A. B. (2020). Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation? International Journal of Molecular Sciences, 21(1), 146. https://doi.org/10.3390/ijms21010146