2.1. Effects of Abiotic Atresses on the Physiological Properties of GB
The first and basic physiological response of plants to drought stress is decreased in photosynthetic efficiency as a result of the stomata closing. This process is initiated within 1–2 min after the occurrence of the causative agent, and completed within 5 min. Maintaining the balance between CO
2 exchange and transpiration is necessary to maximize CO
2 assimilation in photosynthesis and, at the same time, reduce water loss [
20].
There were significant differences in the photosynthesis rate (parameter A) between the varieties, within the light intensity and under the influence of drought (
Table 1). The highest photosynthesis intensity expressed by CO
2 assimilation was found in control plants at a light intensity of PAR = 800 µmol m
−2 s
−1, with the
Nokia variety having higher photosynthetic activity than
KWS Olof. Control plants growing at PAR = 400 µmol m
−2 s
−1 light intensity showed significantly lower photosynthesis activity for the
Nokia variety by 24.1%, while for the
KWS Olof variety by 20.3%. Similarly, at PAR = 800 µmol m
−2 s
−1, plants of the
Nokia variety were characterized by higher intensity of CO
2 assimilation compared to the
KWS Olof variety.
Abiotic stress in the form of water deficiency in both varieties caused a significant decrease in CO2 assimilation, however at a different level. In the Nokia variety, the decrease in CO2 assimilation due to drought was 91.6% at PAR = 800 µmol m−2 s−1 and 87.8% at PAR = 400 µmol m−2 s−1, while in KWS Olof by 67.6% and 18.4%, respectively. In the KWS Olof variety, a much smaller decrease in CO2 assimilation as a result of drought was observed, while stress was intensified by increasing the intensity of PAR. The KWS Olof variety was characterized by lower CO2 assimilation under control conditions, but retained higher in drought stress. Smaller, but significant differences between varieties were noted in the transpiration level (parameter E). The highest water evaporation was observed in control plants at PAR = 800 µmol m−2 s−1, while the reduction of transpiration as a result of lowering PAR was 18.3% for the KWS Olof variety, and 26.1% for Nokia. Similarly to the intensity of CO2 assimilation, in the KWS Olof variety the decrease in transpiration as a result of drought was smaller (at PAR = 800 µmol m−2 s−1 by 78.3%, at PAR = 400 µmol m−2 s−1 by 35.3%) than in Nokia (at PAR = 800 µmol m−2 s−1 by 90, 6%, at PAR = 400 µmol m−2 s−1 by 84.2%).
Stress-reduced CO
2 assimilation can lead to an imbalance between the supply and demand of ATP and NADPH assimilation strength in photosynthesis. In such a situation, or if the antenna complexes provide too much energy in drought conditions, there may be an increased generation of reactive oxygen species, and consequently irreversible degradation of the components of the photosynthetic apparatus [
21]. The resulting situation forces the plant to various processes of scattering the excess energy absorbed by chlorophyll, and one of them is increased fluorescence. The fluorimetric method allows to estimate the efficiency of the energy conversion process of PAR in the reactions of the light phase of photosynthesis [
22].
There were no significant differences in the quantum photochemical reaction efficiency in PSII (Y) andETR between cultivars in control plants at PAR = 800 µmol m−2 s−1, while at PAR = 400 µmol m−2 s−1 the differences were small. However, differences were observed in plants as a result of drought stress. In the KWS Olof variety, the decrease in Y and ETR parameters as a result of drought was 52% and 40.9% at PAR = 800 µmol m−2 s−1 and 39% and 37.2% at PAR = 400 µmol m−2 s−1, while in the Nokia variety 34.6% and 37.5%, respectively, at PAR = 800 µmol m−2 s−1 and 28.6% and 25.2% at PAR = 400 µmol m−2 s−1. The Nokia variety was, therefore, characterized by a smaller decrease in Y and ETR, as well as higher values of both parameters in plants grown under control conditions.
In drought conditions, the
KWS Olof variety showed a smaller decrease in CO
2 assimilation and transpiration and higher values of these parameters at both light intensities (
Table 2). Only the
KWS Olof variety increased stress as a result of increased light intensity. It can be concluded that the
KWS Olof variety is more resistant to drought stress because it has retained a greater ability to assimilate CO
2 in conditions of water scarcity. This may be due to the greater ability of this variety to regulate heat dissipation (NPQ). The simultaneous reduction of Y and ETR could have been a plant defence response to drought stress. During drought stress, the stomata are closed and the efficiency of CO
2 assimilation decreases, which reduces the consumption of ATP and NADPH and, as a consequence, slows the transport of electrons through photocircuits [
23] and the Yied parameter. According to Zlatev [
24], a significant decrease in the Y parameter, can be considered an indicator of the physiological regulation of electron transport by increasing the quenching process of excitation energy in PSII antennas and indicates the occurrence of photoinhibition. There was also a significant increase in non-photochemical quenching (NPQ) associated with thermal losses. In plants of the
KWS Olof variety, NPQ increase due to drought at PAR = 800 µmol m
−2 s
−1 was 37%, while in the
Nokia variety 79%. However, at PAR = 400 µmol m
−2 s
−1, a decrease in NPQ due to drought was noted in both varieties. Additionally, in the research of Oukarroum et al. [
25], differences between spring barley cultivars in the values of chlorophyll A fluorescence parameters in response to drought stress were noted.
Chlorophyll A fluorescence parameters after dark plant adaptation (F0, Fm and Fv/Fm) did not differ between the tested plants. It can, therefore, be concluded that both light intensity and drought stress did not cause significant damage to Photosystem II.
2.2. Antioxidant Activity
Antioxidant activity was tested by three different methods (
Table 3)—DPPH, FRAP and ABTS. Results from the DPPH method in the
KWS Olof variety ranged from 3.43 to 3.87 mg TEAC per g and from 0.95 to 2.00 mg TEAC per g in the
Nokia variety. While the period of watering, as well as photosynthesis active radiation, had no significant influence on the antioxidant activity by this method in
KWS Olof, in
Nokia these factors had influence, especially under the drought condition, which caused an increase of antioxidant activity. The obtained results from the FRAP method ranged from 12.34 to 19.10 mmol TEAC per g in
KWS Olof and from 11.71 to 18.58 mmol TEAC per g in
Nokia. In both tested varieties drought as an abiotic stress caused an increase of activity in comparison to control samples. The same tendency was also observed in intensive irradiation, in which PAR = 800 µmol m
−2 s
−1 caused increasing antioxidant activity in both varieties. This increasing can be explained by antioxidant enzyme activity, such as superoxide dismutase, catalase and peroxidase, which generally increased significantly in plants under stress [
26]. Results from the ABTS method in
KWS Olof ranged from 64.43 to 66.66 mg TEAC per g and from 63.76 to 65.58 mg TEAC per g.
Table 4 shows that, for the antioxidant activity tested by the ABTS method, the variety and abiotic stress, like drought and intensive irradiation, have no significant influence in comparison to DPPH and FRAP methods. Differences between the antioxidant activity was also observed in study Pérez-Labrada et al. [
27] which observed antioxidant activity of tomato plants under abiotic stress (salinity). DPPH and ABTS behaved differently in the plants: antioxidant activity was decreased in those exposed to abiotic stress (−1.2%) while the activity by ABTS increased (4.7%). Ahmed et al. [
28] reported the increment of antioxidant capacity (FRAP) with increasing abiotic stress (salinity) in barley. In contrast, Neffati et al. [
29] found a decrement in antioxidant activity (DPPH) with increasing abiotic stress (NaCl concentrations) in coriander.
Total phenolic content (
Table 4) in tested samples of variety
KWS Olof ranged from 5.00 to 8.16 mg GAE per g and in variety
Nokia from 5.09 to 6.38 mg GAE per g. While drought conditions caused a decrease of total phenolic content in
KWS Olof, radiation, especially PAR 800 µmol m
−2 s
−1, caused an increase of these compounds in comparison to PAR = 400 µmol m
−2 s
−1. In
Nokia the period of watering, as well as photosynthesis active radiation, had no the significant influence. In the study by Sarker et al. [
30] abiotic stress (salinity) in
Amaranthus tricolor leaves observed a higher amount of total phenolic content in comparison to control variant without stress. Total phenolic acid content in tested samples of
KWS Olof ranged from 0.60 to 0.84 mg CAE per g and in
Nokia from 0.75 to 1.65 mg CAE per g. While in
KWS Olof significant differences were not observed, in
Nokia, radiation PAR = 400 µmol m
−2 s
−1 caused an increase of total phenolic acid content in comparison to PAR = 800 µmol m
−2 s
−1 in control and drought cultivation conditions. From our findings it is noticeable that the biological activity of the tested samples is very strongly influenced not only by the cultivation conditions but also with the variety.
2.4. Identification of Bioactive Compounds
The chemical profile of samples was compared using non-target analysis of LC-MS data collected in the negative mode. The bucket table which contains 600 compounds with intensity higher than 10
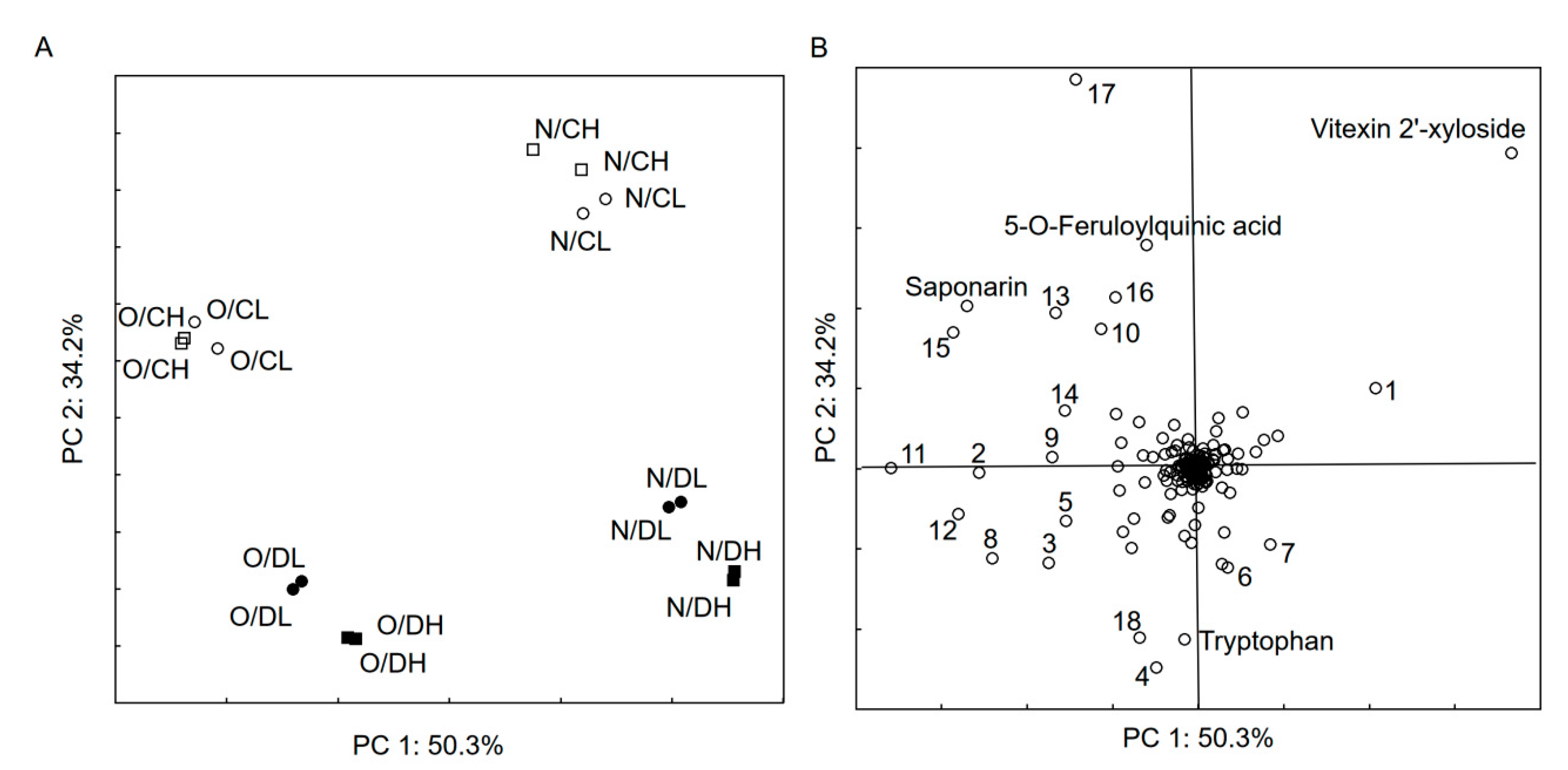
4 cps was generated with MetaboScape 4.0. The PCA results (
Figure 1) show that only a few compounds are relevant and differentiate the sample set. However, it was not possible to identify most of them using applied computational methods (CSI:FingerID and MetFrag). The score plot (
Figure 1A) show clearly that chemical profile of samples not reflect to intensity of radiation, whereas factors, such as barley variety and period of watering, have an influence on the profile of synthesized compounds. Vitexin 2′-xyloside was the most abundant in
Nokia barley, especially control samples. The non-identified compound 568.24 Da (monoisotopic neutral mass), which eluates at 14.5 min, was intensive in the control samples. The D (drought) samples were characterized by a high abundance of tryptophan and two other components (4—304.1 Da_12 min and 18—414.13 Da_15.6 min,
Figure 1B).
In analysed green barleys, many compounds were identified that affect their pro-health value. The details of phenolic compounds which were identified in samples are presented in
Table 6 and the results obtained are consistent with published data [
33]. It has been shown that both irradiation intensity and drought stress have a significant impact on the profile of green barley polyphenolic compounds, with no simple relationship between the effect of stress factors on the content of polyphenols. Literature data indicate that the polyphenolic compounds found in barley not only have antioxidant activity but also hypolipidemic [
9,
34] and anti-ulcerative activity [
35], and some authors also indicate anti-inflammatory or anti-cancer effects [
9,
36].
The phenolic profile of samples and categorical variables, such as period of watering, intensity of radiation and barley variety, were included during k-means clustering with the V-fold cross-validation method. This statistical procedure was performed to show which cultivation factors have an influence on phenolic components. The results are presented in
Table 7 and two clusters were generated. It was noticed that only the period of watering had a discriminative power (
p < 0.05), while other cultivation variables were not relevant. The most discriminative phenolic compounds (the highest F value) were sinapic acid, ferulic acid and sinapoyl-beta-glucose. However, saponarin, vicetin-2 and vitexin 2′-xyloside dominated in samples, but they are not statistically different in clusters (
p > 0.05).

 ,
, 





{kind=link}