
IRAG2 Interacts with IP3-Receptor Types 1, 2, and 3 and Regulates Intracellular Ca2+ in Murine Pancreatic Acinar Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
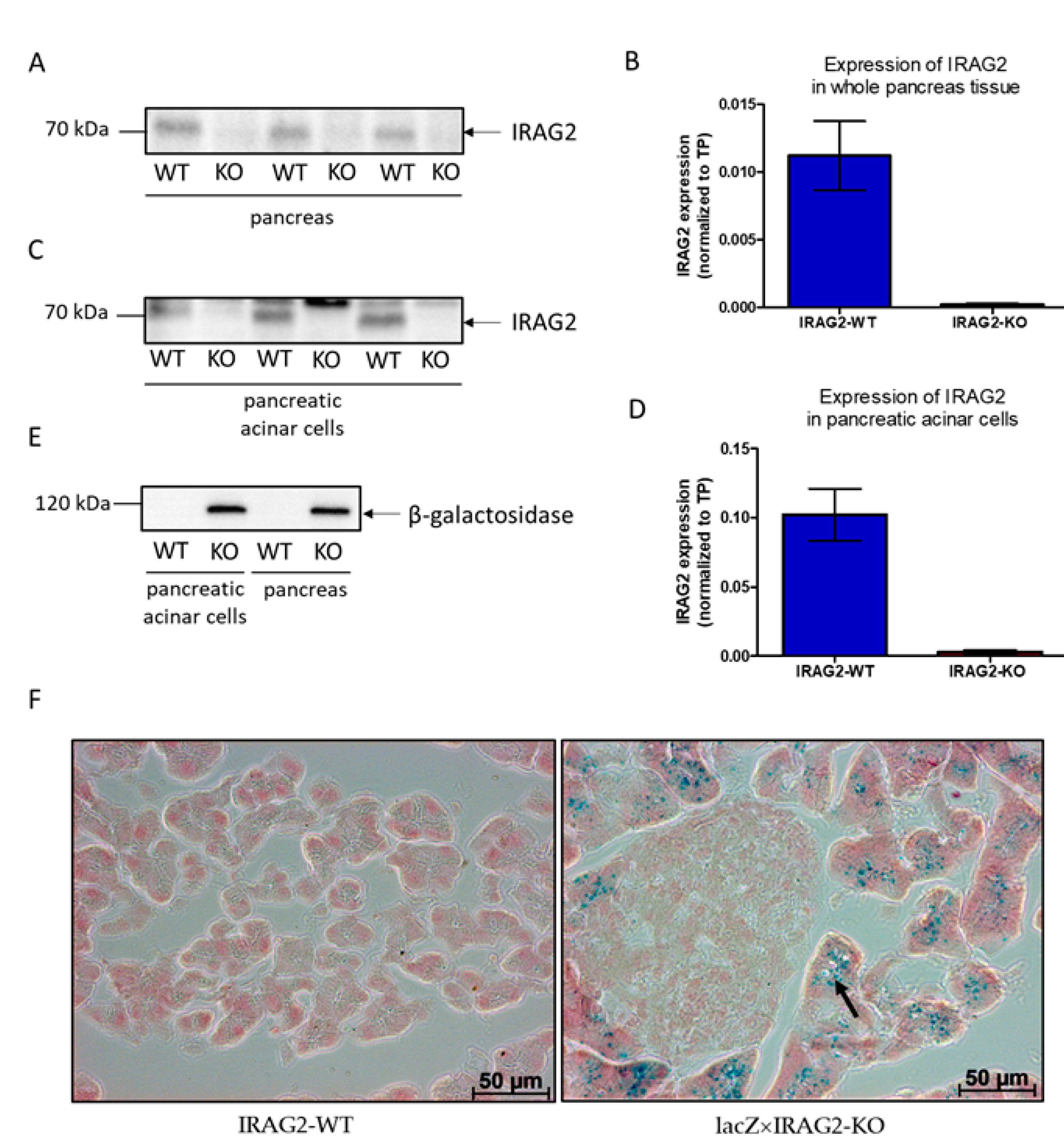
2.1. Expression of IRAG2 in Pancreatic Acinar Cells
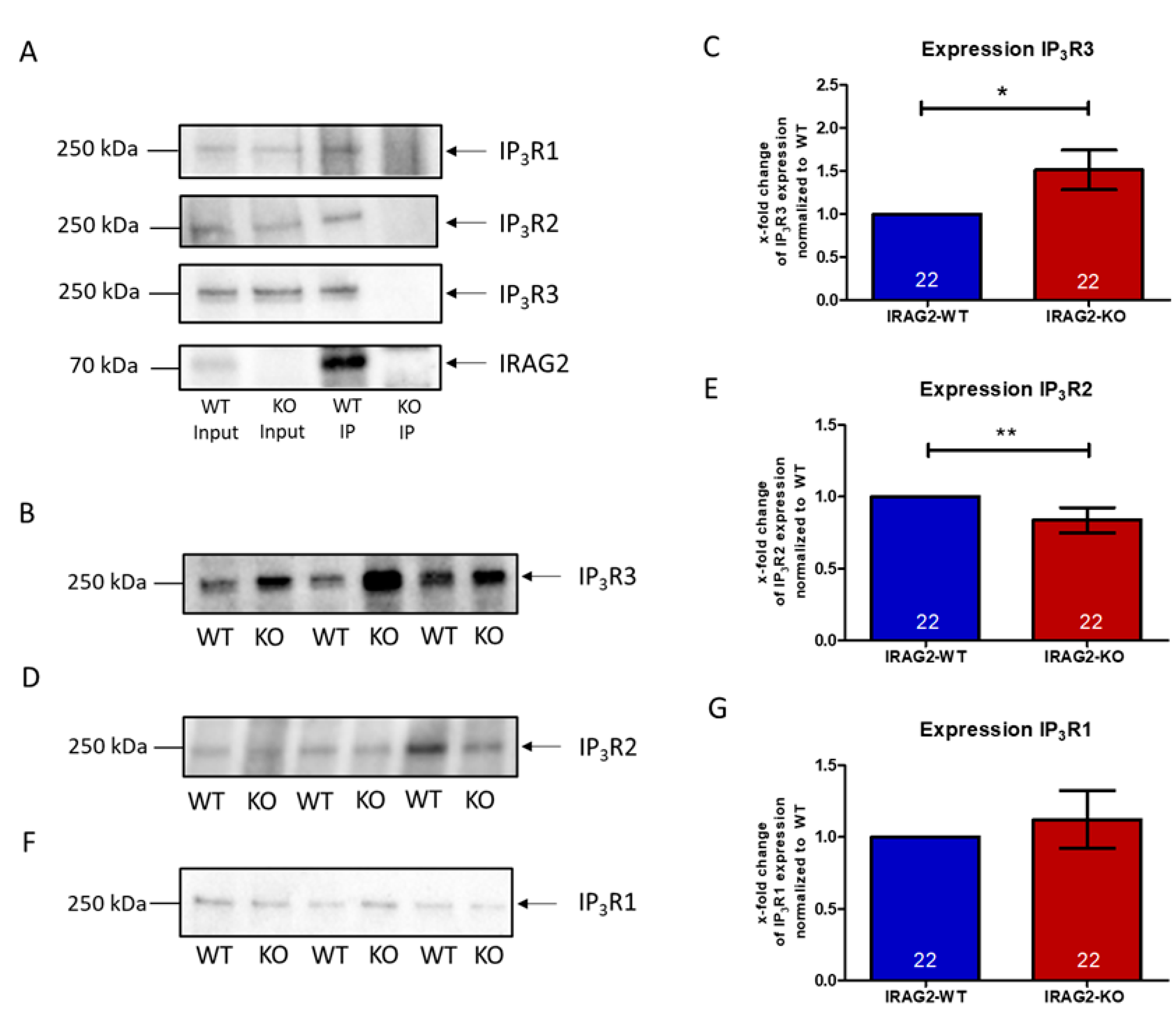
2.2. Interaction of IRAG2 with IP3R Types 1, 2, and 3 and Regulation of IP3R Expression
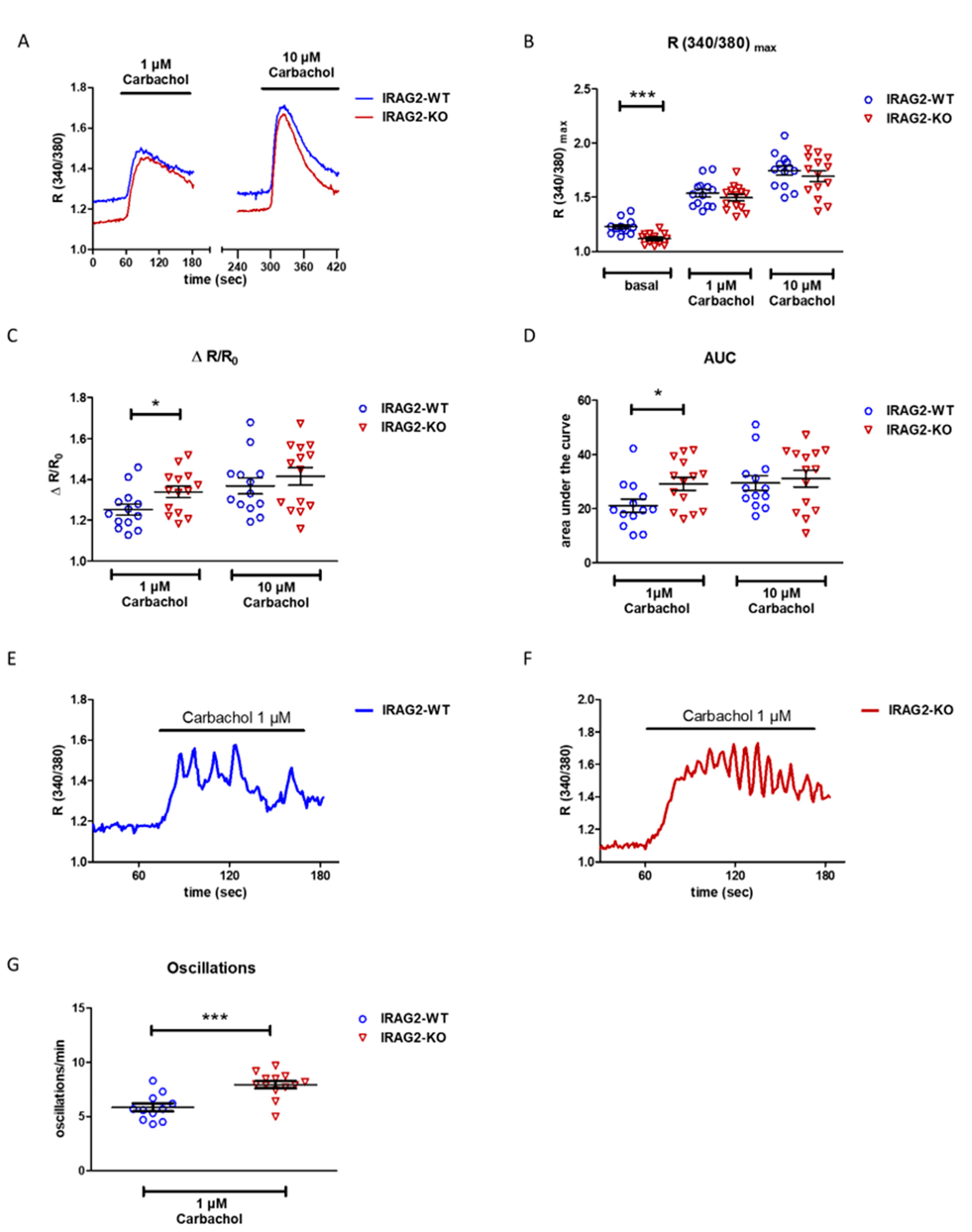
2.3. Intracellular Ca2+ Regulation in IRAG2-KO Pancreatic Acinar Cells
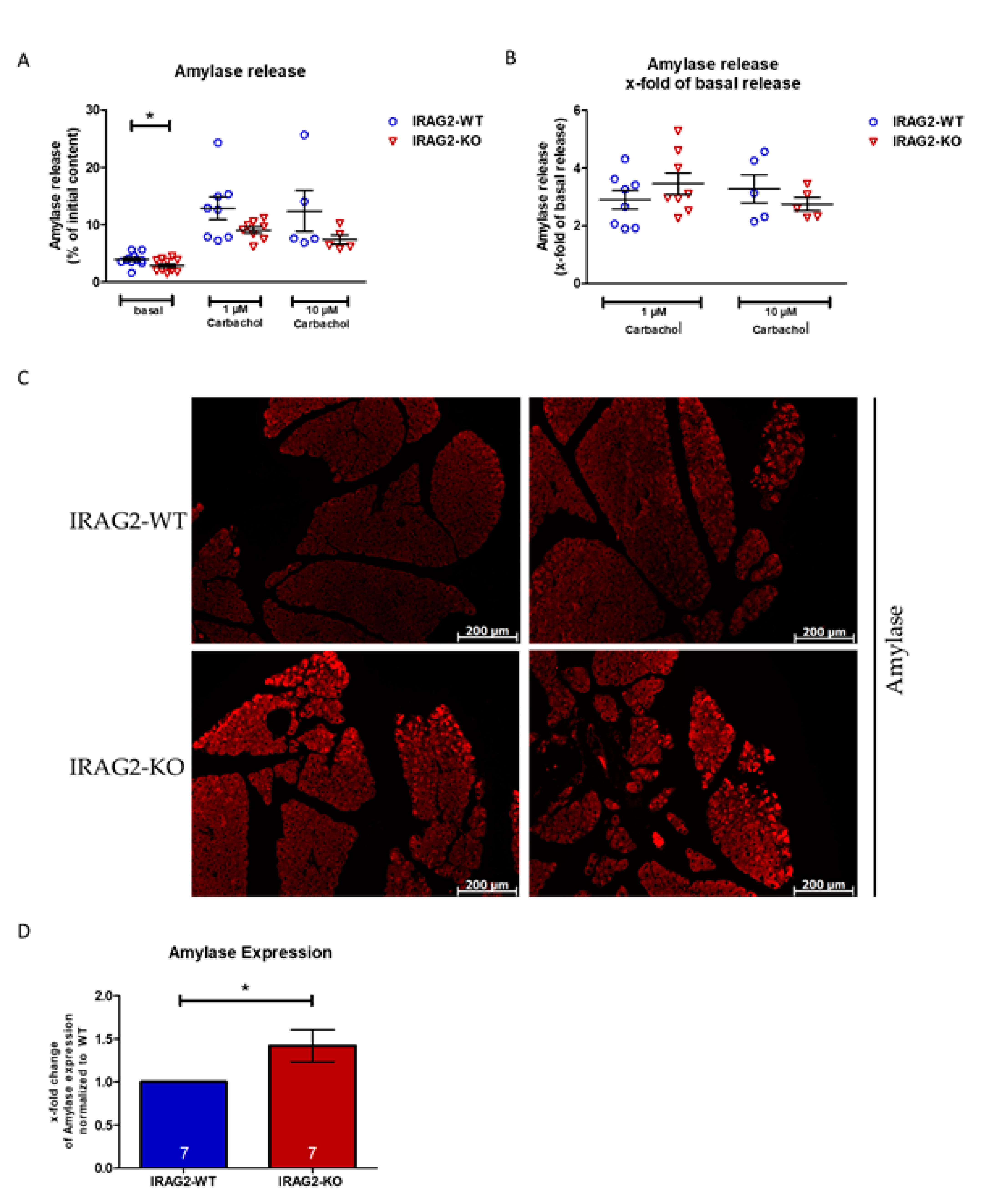
2.4. Amylase Content and Release in IRAG2-WT and IRAG2-KO Pancreatic Acinar Cells
3. Discussion
- IRAG2 is expressed in acinar cells of the murine exocrine pancreas and interacts with IP3 receptor types 1, 2, and 3 in the murine pancreas. Loss of IRAG2 leads to an upregulation of IP3R3 and to downregulation of IP3R2 expression in the pancreas.
- Deletion of IRAG2 leads to lower basal Ca2+ release in murine pancreatic acinar cells, suggesting that IRAG2 leads to activation of IP3R under unstimulated basal conditions. However, IRAG2-KO reveals a higher amount of released Ca2+ normalized to basal release upon stimulation with 1 µM carbachol compared to IRAG2-WT. The frequency of Ca2+ oscillations is higher in IRAG2-KO pancreatic acinar cells compared to IRAG2-WT, maybe due to modulation of IP3 receptors through IRAG2.
- Decreased basal Ca2+ release in IRAG2-KO acinar cells leads to lower basal amylase secretion. Lower release of amylase may cause a higher amount of amylase that remains in the zymogen granules of IRAG2-KO.
4. Materials and Methods
4.1. Animals
4.2. Tissue Preparation for Western Blot Analysis
4.3. Coimmunoprecipitation
4.4. Western Blot Analysis
4.5. Preparation of Dispersed Mouse Pancreatic Acinar Cells
4.6. Measurement of Intracellular Calcium
4.7. Amylase Secretion Assay
4.8. X-Gal-Staining
4.9. Immunohistochemistry
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petersen, O.H. Ca2+ signaling in pancreatic acinar cells: Physiology and pathophysiology. Braz J. Med. Biol Res. 2009, 42, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Hattori, M.; Suzuki, A.Z.; Higo, T.; Miyauchi, H.; Michikawa, T.; Nakamura, T.; Inoue, T.; Mikoshiba, K. Distinct roles of inositol 1,4,5-trisphosphate receptor types 1 and 3 in Ca2+ signaling. J. Biol. Chem. 2004, 279, 11967–11975. [Google Scholar] [CrossRef] [Green Version]
- Futatsugi, A.; Nakamura, T.; Yamada, M.K.; Ebisui, E.; Nakamura, K.; Uchida, K.; Kitaguchi, T.; Takahashi-Iwanaga, H.; Noda, T.; Aruga, J.; et al. IP3 receptor types 2 and 3 mediate exocrine secretion underlying energy metabolism. Science 2005, 309, 2232–2234. [Google Scholar] [CrossRef]
- Yule, D.I.; Ernst, S.A.; Ohnishi, H.; Wojcikiewicz, R.J. Evidence that zymogen granules are not a physiologically relevant calcium pool. Defining the distribution of inositol 1,4,5-trisphosphate receptors in pancreatic acinar cells. J. Biol. Chem. 1997, 272, 9093–9098. [Google Scholar] [CrossRef] [Green Version]
- Streb, H.; Irvine, R.F.; Berridge, M.J.; Schulz, I. Release of Ca2+ from a nonmitochondrial intracellular store in pancreatic acinar cells by inositol-1,4,5-trisphosphate. Nature 1983, 306, 67–69. [Google Scholar] [CrossRef]
- Behrens, T.W.; Jagadeesh, J.; Scherle, P.; Kearns, G.; Yewdell, J.; Staudt, L.M. Jaw1, A lymphoid-restricted membrane protein localized to the endoplasmic reticulum. J. Immunol. 1994, 153, 682–690. [Google Scholar]
- Behrens, T.W.; Kearns, G.M.; Rivard, J.J.; Bernstein, H.D.; Yewdell, J.W.; Staudt, L.M. Carboxyl-terminal Targeting and Novel Post-translational Processing of JAW1, a Lymphoid Protein of the Endoplasmic Reticulum. J. Biol. Chem. 1996, 271, 23528–23534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, C.H.; Myers, M.E.; Juchno, J.; Haimbaugh, C.; Bichraoui, H.; Du, Y.; Bankston, J.R.; Walker, L.A.; Proenza, C. Isoform-specific regulation of HCN4 channels by a family of endoplasmic reticulum proteins. Proc. Natl. Acad. Sci. USA 2020, 117, 18079–18090. [Google Scholar] [CrossRef] [PubMed]
- Shindo, Y.; Kim, M.-R.; Miura, H.; Yuuki, T.; Kanda, T.; Hino, A.; Kusakabe, Y. Lrmp/Jaw1 is Expressed in Sweet, Bitter, and Umami Receptor–Expressing Cells. Chem. Sens. 2010, 35, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Shaughnessy, J.D.; Largaespada, D.A.; Tian, E.; Fletcher, C.F.; Cho, B.C.; Vyas, P.; Jenkins, N.A.; Copeland, N.G. Mrvi1, a common MRV integration site in BXH2 myeloid leukemias, encodes a protein with homology to a lymphoid-restricted membrane protein Jaw1. Oncogene 1999, 18, 2069–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiselhöringer, A.; Werner, M.; Sigl, K.; Smital, P.; Wörner, R.; Acheo, L.; Stieber, J.; Weinmeister, P.; Feil, R.; Feil, S.; et al. IRAG is essential for relaxation of receptor-triggered smooth muscle contraction by cGMP kinase. EMBO J. 2004, 23, 4222–4231. [Google Scholar] [CrossRef] [Green Version]
- Schlossmann, J.; Ammendola, A.; Ashman, K.; Zong, X.; Huber, A.; Neubauer, G.; Wang, G.X.; Allescher, H.D.; Korth, M.; Wilm, M.; et al. Regulation of intracellular calcium by a signalling complex of IRAG, IP3 receptor and cGMP kinase Ibeta. Nature 2000, 404, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-Y.; Wang, J.; Zhao, Y.; Liu, J.; Yang, X.; Yue, X.; Wang, H.; Zhou, F.; Inclan-Rico, J.M.; Ponessa, J.J.; et al. Tumor suppressor p53 regulates intestinal type 2 immunity. Nat. Commun. 2021, 12, 3371. [Google Scholar] [CrossRef]
- Tedoldi, S.; Paterson, J.C.; Cordell, J.; Tan, S.-Y.; Jones, M.; Manek, S.; Dei Tos, A.P.; Roberton, H.; Masir, N.; Natkunam, Y.; et al. Jaw1/LRMP, a germinal centre-associated marker for the immunohistological study of B-cell lymphomas. J. Pathol. 2006, 209, 454–463. [Google Scholar] [CrossRef]
- Geiselhöringer, A.; Gaisa, M.; Hofmann, F.; Schlossmann, J. Distribution of IRAG and cGKI-isoforms in murine tissues. FEBS Lett. 2004, 575, 19–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yule, D.I.; Gallacher, D.V. Oscillations of cytosolic calcium in single pancreatic acinar cells stimulated by acetylcholine. FEBS Lett. 1988, 239, 358–362. [Google Scholar] [CrossRef] [Green Version]
- Tsunoda, Y.; Stuenkel, E.L.; Williams, J.A. Oscillatory mode of calcium signaling in rat pancreatic acinar cells. Am. J. Physiol. 1990, 258, C147–C155. [Google Scholar] [CrossRef]
- Herbert, Y.G.; Dolai, S.; Takahashi, T. Pancreapedia: Exocrine Pancreas Knowledge Base. Pancreapedia Exocrine Pancreas Knowl. Base 2020. [Google Scholar] [CrossRef]
- Matthews, E.K.; Petersen, O.H.; Williams, J.A. Pancreatic acinar cells: Acetylcholine-induced membrane depolarization, calcium efflux and amylase release. J. Physiol. 1973, 234, 689–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Matsumoto, M.; Nakagawa, T.; Inoue, T.; Nagata, E.; Tanaka, K.; Takano, H.; Minowa, O.; Kuno, J.; Sakakibara, S.; Yamada, M.; et al. Ataxia and epileptic seizures in mice lacking type 1 inositol 1,4,5-trisphosphate receptor. Nature 1996, 379, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Kato, K.; Kohda, K.; Mikoshiba, K. Type 1 Inositol 1,4,5-Trisphosphate Receptor Is Required for Induction of Long-Term Depression in Cerebellar Purkinje Neurons. J. Neurosci. 1998, 18, 5366–5373. [Google Scholar] [CrossRef]
- Nishiyama, M.; Hong, K.; Mikoshiba, K.; Poo, M.M.; Kato, K. Calcium stores regulate the polarity and input specificity of synaptic modification. Nature 2000, 408, 584–588. [Google Scholar] [CrossRef]
- Fujii, S.; Matsumoto, M.; Igarashi, K.; Kato, H.; Mikoshiba, K. Synaptic plasticity in hippocampal CA1 neurons of mice lacking type 1 inositol-1,4,5-trisphosphate receptors. Learn. Mem. 2000, 7, 312–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojcikiewicz, R.J. Type I, II, and III inositol 1,4,5-trisphosphate receptors are unequally susceptible to down-regulation and are expressed in markedly different proportions in different cell types. J. Biol. Chem. 1995, 270, 11678–11683. [Google Scholar] [CrossRef] [Green Version]
- Iino, M. Biphasic Ca2+ dependence of inositol 1,4,5-trisphosphate-induced Ca release in smooth muscle cells of the guinea pig taenia caeci. J. Gen. Physiol. 1990, 95, 1103–1122. [Google Scholar] [CrossRef] [Green Version]
- Finch, E.A.; Turner, T.J.; Goldin, S.M. Calcium as a coagonist of inositol 1,4,5-trisphosphate-induced calcium release. Science 1991, 252, 443–446. [Google Scholar] [CrossRef]
- Bezprozvanny, I.; Watras, J.; Ehrlich, B.E. Bell-shaped calcium-response curves of Ins(1,4,5)P3- and calcium-gated channels from endoplasmic reticulum of cerebellum. Nature 1991, 351, 751–754. [Google Scholar] [CrossRef]
- Miyakawa, T.; Maeda, A.; Yamazawa, T.; Hirose, K.; Kurosaki, T.; Iino, M. Encoding of Ca2+ signals by differential expression of IP3 receptor subtypes. EMBO J. 1999, 18, 1303–1308. [Google Scholar] [CrossRef]
- Giovannucci, D.R.; Groblewski, G.E.; Sneyd, J.; Yule, D.I. Targeted phosphorylation of inositol 1,4,5-trisphosphate receptors selectively inhibits localized Ca2+ release and shapes oscillatory Ca2+ signals. J. Biol. Chem. 2000, 275, 33704–33711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straub, S.V.; Giovannucci, D.R.; Bruce, J.I.E.; Yule, D.I. A role for phosphorylation of inositol 1,4,5-trisphosphate receptors in defining calcium signals induced by Peptide agonists in pancreatic acinar cells. J. Biol. Chem. 2002, 277, 31949–31956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, K.; Petersen, O.H.; Tepikin, A.V. Dual sensitivity of sarcoplasmic/endoplasmic Ca2+-ATPase to cytosolic and endoplasmic reticulum Ca2+ as a mechanism of modulating cytosolic Ca2+ oscillations. Biochem. J. 2004, 383, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.A. Isolation of rodent pancreatic acinar cells and acini by collagenase digestion. Pancreapedia Exocrine Pancreas Knowl. Base 2010. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.; Williams, J.A. A role for Rho and Rac in secretagogue-induced amylase release by pancreatic acini. Am. J. Physiol. Cell Physiol. 2005, 289, C22–C32. [Google Scholar] [CrossRef]
- Straubinger, J.; Boldt, K.; Kuret, A.; Deng, L.; Krattenmacher, D.; Bork, N.; Desch, M.; Feil, R.; Feil, S.; Nemer, M.; et al. Amplified pathogenic actions of angiotensin II in cysteine-rich LIM-only protein 4-negative mouse hearts. FASEB J. 2017, 31, 1620–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, D.B.; Pasumarthi, R.K.; Baridon, B.; Djan, E.; Trainor, A.; Griffey, S.M.; Engelhard, E.K.; Rapp, J.; Li, B.; de Jong, P.J.; et al. A lacZ reporter gene expression atlas for 313 adult KOMP mutant mouse lines. Genome Res. 2015, 25, 598–607. [Google Scholar] [CrossRef] [Green Version]
- Kühbandner, S.; Brummer, S.; Metzger, D.; Chambon, P.; Hofmann, F.; Feil, R. Temporally controlled somatic mutagenesis in smooth muscle. Genesis 2000, 28, 15–22. [Google Scholar] [CrossRef]
- Schelter, F.; Gerg, M.; Halbgewachs, B.; Schaten, S.; Görlach, A.; Schrötzlmair, F.; Krüger, A. Identification of a survival-independent metastasis-enhancing role of hypoxia-inducible factor-1alpha with a hypoxia-tolerant tumor cell line. J. Biol. Chem. 2010, 285, 26182–26189. [Google Scholar] [CrossRef] [Green Version]
- Schinner, E.; Schramm, A.; Kees, F.; Hofmann, F.; Schlossmann, J. The cyclic GMP-dependent protein kinase Iα suppresses kidney fibrosis. Kidney Int. 2013, 84, 1198–1206. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prüschenk, S.; Majer, M.; Schreiber, R.; Schlossmann, J. IRAG2 Interacts with IP3-Receptor Types 1, 2, and 3 and Regulates Intracellular Ca2+ in Murine Pancreatic Acinar Cells. Int. J. Mol. Sci. 2021, 22, 13409. https://doi.org/10.3390/ijms222413409
Prüschenk S, Majer M, Schreiber R, Schlossmann J. IRAG2 Interacts with IP3-Receptor Types 1, 2, and 3 and Regulates Intracellular Ca2+ in Murine Pancreatic Acinar Cells. International Journal of Molecular Sciences. 2021; 22(24):13409. https://doi.org/10.3390/ijms222413409
Chicago/Turabian StylePrüschenk, Sally, Michael Majer, Rainer Schreiber, and Jens Schlossmann. 2021. "IRAG2 Interacts with IP3-Receptor Types 1, 2, and 3 and Regulates Intracellular Ca2+ in Murine Pancreatic Acinar Cells" International Journal of Molecular Sciences 22, no. 24: 13409. https://doi.org/10.3390/ijms222413409
APA StylePrüschenk, S., Majer, M., Schreiber, R., & Schlossmann, J. (2021). IRAG2 Interacts with IP3-Receptor Types 1, 2, and 3 and Regulates Intracellular Ca2+ in Murine Pancreatic Acinar Cells. International Journal of Molecular Sciences, 22(24), 13409. https://doi.org/10.3390/ijms222413409