
The Sexually Dimorphic Adrenal Cortex: Implications for Adrenal Disease


{kind=link}
Abstract
:1. Introduction
2. Sex Bias in Adrenocortical Diseases
3. Sexual Dimorphism in the Adrenal Cortex
3.1. Tissue Renewal and Homeostasis
3.2. Sexual Dimorphism in Mouse Models of Adrenal Disease
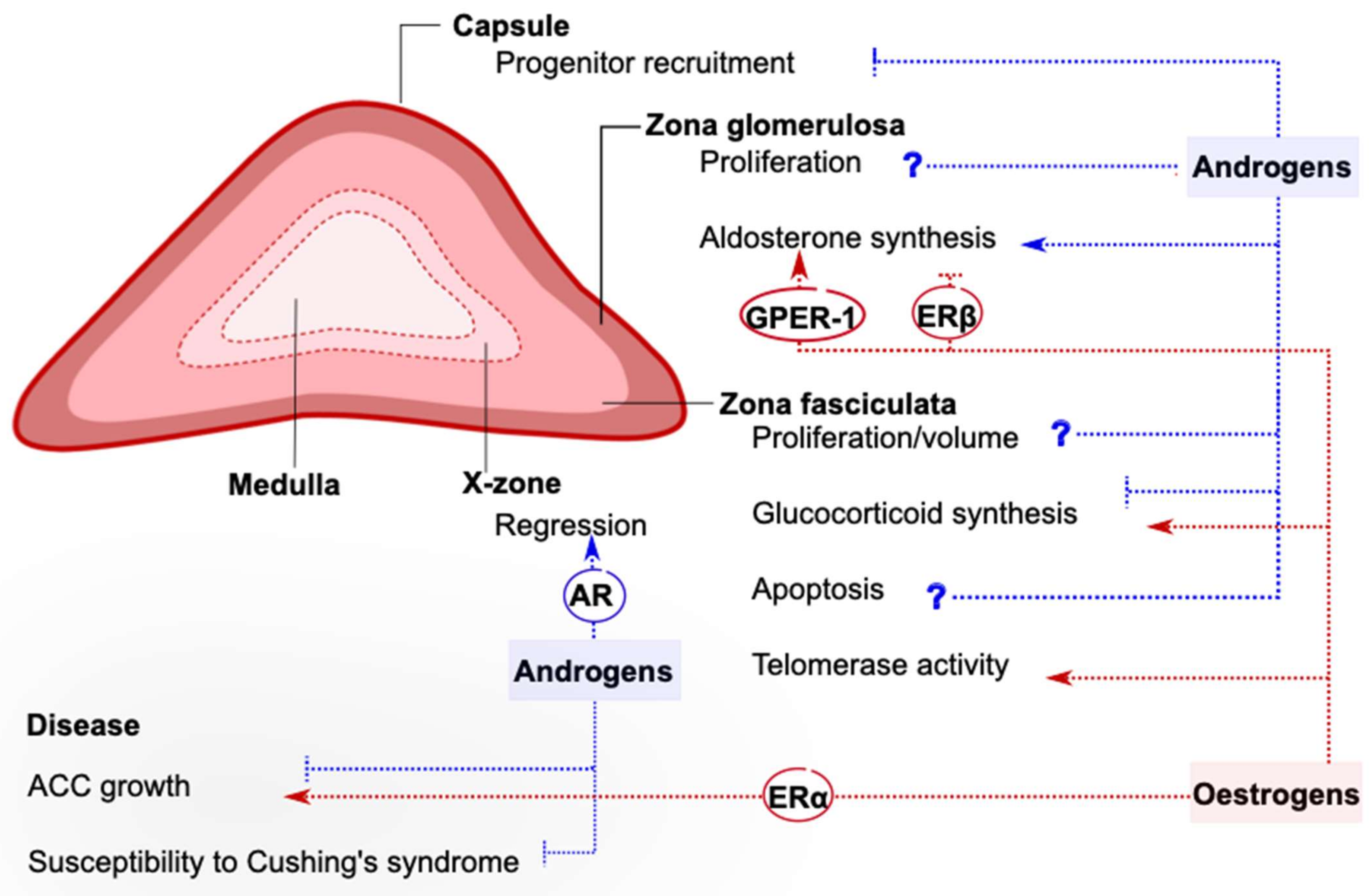
4. Impact of Gonadal Steroid Signaling in the Adrenal Cortex
4.1. Expression of Sex Steroid-Related Genes in the Healthy Adrenal Cortex
4.2. Gonadal Hormones and Adrenal Steroidogenesis
4.3. Effects of Abolishing Androgen Receptor Signaling in the Adrenal Cortex
4.4. Role of Oestrogen Signaling in Telomere Maintenance in the Adrenal Cortex
4.5. Sex Hormones in the Context of Adrenocortical Tumors
5. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Credendino, S.C.; Neumayer, C.; Cantone, I. Genetics and Epigenetics of Sex Bias: Insights from Human Cancer and Autoimmunity. Trends Genet. 2020, 36, 650–663. [Google Scholar] [CrossRef]
- Clocchiatti, A.; Cora, E.; Zhang, Y.; Dotto, G.P. Sexual dimorphism in cancer. Nat. Rev. Cancer 2016, 16, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Karp, N.A.; Mason, J.; Beaudet, A.L.; Benjamini, Y.; Bower, L.; Braun, R.E.; Brown, S.D.M.; Chesler, E.J.; Dickinson, M.E.; Flenniken, A.M.; et al. Prevalence of sexual dimorphism in mammalian phenotypic traits. Nat. Commun. 2017, 8, 15475. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, S.; Xu, Y.; Zhang, Y.; Ma, W.; Liang, C.; Lu, H.; Ji, Y.; Liu, C.; Chen, D.; et al. Androgen Maintains Intestinal Homeostasis by Inhibiting BMP Signaling via Intestinal Stromal Cells. Stem Cell Reports 2020, 15, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Grabek, A.; Dolfi, B.; Klein, B.; Jian-Motamedi, F.; Chaboissier, M.-C.; Schedl, A. The Adult Adrenal Cortex Undergoes Rapid Tissue Renewal in a Sex-Specific Manner. Cell Stem Cell 2019, 25, 290–296.e2. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Mcclusky, R.; Chen, J.; Beaven, S.W.; Tontonoz, P. The Number of X Chromosomes Causes Sex Differences in Adiposity in Mice. PLoS Genet 2012, 8, 1002709. [Google Scholar] [CrossRef] [Green Version]
- Beery, A.K.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2011, 35, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Accounting for sex and gender makes for better science. Nature 2020, 588, 196. [CrossRef] [PubMed]
- Ennen, W.B.; Levay-Young, B.K.; Engeland, W.C. Zone-specific cell proliferation during adrenocortical regeneration after enucleation in rats. Am. J. Physiol. Metab. 2005, 289, E883–E891. [Google Scholar] [CrossRef] [Green Version]
- Finco, I.; Mohan, D.R.; Hammer, G.D.; Lerario, A.M. Regulation of stem and progenitor cells in the adrenal cortex. Curr. Opin. Endocr. Metab. Res. 2019, 8, 66–71. [Google Scholar] [CrossRef]
- Oikonomakos, I.; Weerasinghe Arachchige, L.C.; Schedl, A. Developmental mechanisms of adrenal cortex formation and their links with adult progenitor populations. Mol. Cell. Endocrinol. 2021, 524. [Google Scholar] [CrossRef]
- Lacroix, A.; Feelders, R.A.; Stratakis, C.A.; Nieman, L.K. Cushing’s syndrome. Lancet 2015, 386, 913–927. [Google Scholar] [CrossRef]
- Lindholm, J.; Juul, S.; Jørgensen, J.O.L.; Astrup, J.; Bjerre, P.; Feldt-Rasmussen, U.; Hagen, C.; Jørgensen, J.; Kosteljanetz, M.; Kristensen, L.Ø.; et al. Incidence and Late Prognosis of Cushing’s Syndrome: A Population-Based Study 1. J. Clin. Endocrinol. Metab. 2001, 86, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, C.; Bak, A.M.; Zøylner Rubeck, K.; Jørgensen, J.O.L. Epidemiology of Cushing’s syndrome. Neuroendocrinology 2010, 92 (Suppl. 1), 1–5. [Google Scholar]
- Bertherat, J.; Horvath, A.; Groussin, L.; Grabar, S.; Boikos, S.; Cazabat, L.; Libe, R.; René-Corail, F.; Stergiopoulos, S.; Bourdeau, I.; et al. Mutations in regulatory subunit type 1A of cyclic adenosine 5′-monophosphate-dependent protein kinase (PRKAR1A): Phenotype analysis in 353 patients and 80 different genotypes. J. Clin. Endocrinol. Metab. 2009, 94, 2085–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audenet, F.; Méjean, A.; Chartier-Kastler, E.; Rouprêt, M. Adrenal tumours are more predominant in females regardless of their histological subtype: A review. World J. Urol. 2013, 31, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Crona, J.; Beuschlein, F. Adrenocortical carcinoma—Towards genomics guided clinical care. Nat. Rev. Endocrinol. 2019, 15. [Google Scholar] [CrossRef]
- Fassnacht, M.; Libé, R.; Kroiss, M.; Allolio, B. Adrenocortical carcinoma: A clinician’s update. Nat. Rev. Endocrinol. 2011, 7, 323–335. [Google Scholar] [CrossRef]
- Luton, J.P.; Cerdas, S.; Billaud, L.; Thomas, G.; Guilhaume, B.; Bertagna, X.; Laudat, M.H.; Louvel, A.; Chapuis, Y.; Blondeau, P.; et al. Clinical features of adrenocortical carcinoma, prognostic factors, and the effect of mitotane therapy. N. Engl. J. Med. 1990, 322, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Wooten, M.D.; King, D.K. Adrenal cortical carcinoma. Epidemiology and treatment with mitotane and a review of the literature. Cancer 1993, 72, 3145–3155. [Google Scholar] [CrossRef]
- Ayala-Ramirez, M.; Jasim, S.; Feng, L.; Ejaz, S.; Deniz, F.; Busaidy, N.; Waguespack, S.G.; Naing, A.; Sircar, K.; Wood, C.G.; et al. Adrenocortical carcinoma: Clinical outcomes and prognosis of 330 patients at a tertiary care Center. Eur. J. Endocrinol. 2013, 169, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Scollo, C.; Russo, M.; Trovato, M.A.; Sambataro, D.; Giuffrida, D.; Manusia, M.; Sapuppo, G.; Malandrino, P.; Vigneri, R.; Pellegriti, G. Prognostic factors for adrenocortical carcinoma outcomes. Front. Endocrinol. (Lausanne). 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, L.S.; Kay, R. Adrenocortical carcinoma in children: Review and recent innovations. Urol. Clin. North Am. 2000, 27, 403–421. [Google Scholar] [CrossRef]
- Michalkiewicz, E.; Sandrini, R.; Figueiredo, B.; Miranda, E.C.M.; Caran, E.; Oliveira-Filho, A.G.; Marques, R.; Pianovski, M.A.D.; Lacerda, L.; Cristofani, L.M.; et al. Clinical and outcome characteristics of children with adrenocortical tumors: A report from the international pediatric adrenocortical tumor registry. J. Clin. Oncol. 2004, 22, 838–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzon, L.; Sonino, N.; Fallo, F.; Palù, G.; Boscaro, M. Prevalence and natural history of adrenal incidentalomas. Eur. J. Endocrinol. 2003, 149, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Luton, J.P.; Martinez, M.; Coste, J.; Bertherat, J. Outcome in patients with adrenal incidentaloma selected for surgery: An analysis of 88 cases investigated in a single clinical center. Eur. J. Endocrinol. 2000, 143, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.; Scholl, U.I.; Yue, P.; Björklund, P.; Zhao, B.; Nelson-Williams, C.; Ji, W.; Cho, Y.; Patel, A.; Men, C.J.; et al. K+ channel mutations in adrenal aldosterone-producing adenomas and hereditary hypertension. Science 2011, 331, 768–772. [Google Scholar] [CrossRef] [Green Version]
- Fernandes-Rosa, F.L.; Williams, T.A.; Riester, A.; Steichen, O.; Beuschlein, F.; Boulkroun, S.; Strom, T.M.; Monticone, S.; Amar, L.; Meatchi, T.; et al. Genetic spectrum and clinical correlates of somatic mutations in aldosterone-producing adenoma. Hypertension 2014, 64, 354–361. [Google Scholar] [CrossRef]
- Boulkroun, S.; Beuschlein, F.; Rossi, G.P.; Golib-Dzib, J.F.; Fischer, E.; Amar, L.; Mulatero, P.; Samson-Couterie, B.; Hahner, S.; Quinkler, M.; et al. Prevalence, clinical, and molecular correlates of KCNJ5 mutations in primary aldosteronism. Hypertension 2012, 59, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Lenzini, L.; Rossitto, G.; Maiolino, G.; Letizia, C.; Funder, J.W.; Rossi, G.P. A meta-analysis of somatic KCNJ5 K+ channel mutations in 1636 patients with an aldosterone-producing adenoma. J. Clin. Endocrinol. Metab. 2015, 100, E1089–E1095. [Google Scholar] [CrossRef] [Green Version]
- Heitzmann, D.; Derand, R.; Jungbauer, S.; Bandulik, S.; Sterner, C.; Schweda, F.; El Wakil, A.; Lalli, E.; Guy, N.; Mengual, R.; et al. Invalidation of TASK1 potassium channels disrupts adrenal gland zonation and mineralocorticoid homeostasis. EMBO J. 2008, 27, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Kong, M.-F.; Jeffcoate, W. Eighty-six cases of Addison’s disease. Clin. Endocrinol. (Oxf). 1994, 41, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Myhre, A.G.; Undlien, D.E.; Løvås, K.; Uhlving, S.; Nedrebø, B.G.; Fougner, K.J.; Trovik, T.; Sørheim, J.I.; Husebye, E.S. Autoimmune Adrenocortical Failure in Norway Autoantibodies and Human Leukocyte Antigen Class II Associations Related to Clinical Features. J. Clin. Endocrinol. Metab. 2002, 87, 618–623. [Google Scholar] [CrossRef]
- Bielohuby, M.; Herbach, N.; Wanke, R.; Maser-Gluth, C.; Beuschlein, F.; Wolf, E.; Hoeflich, A. Growth analysis of the mouse adrenal gland from weaning to adulthood: Time- and gender-dependent alterations of cell size and number in the cortical compartment. Am. J. Physiol. - Endocrinol. Metab. 2007, 293. [Google Scholar] [CrossRef]
- Huang, C.-C.J.; Kang, Y. The transient cortical zone in the adrenal gland: The mystery of the adrenal X-zone. J. Endocrinol. 2019, 241, R51–R63. [Google Scholar] [CrossRef]
- Chang, S.P.; Morrison, H.D.; Nilsson, F.; Kenyon, C.J.; West, J.D.; Morley, S.D. Cell proliferation, movement and differentiation during maintenance of the adult mouse adrenal cortex. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- King, P.; Paul, A.; Laufer, E. Shh signaling regulates adrenocortical development and identifies progenitors of steroidogenic lineages. Proc. Natl. Acad. Sci. USA 2009, 106, 21185–21190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-C.J.; Miyagawa, S.; Matsumaru, D.; Parker, K.L.; Yao, H.H.-C. Progenitor Cell Expansion and Organ Size of Mouse Adrenal Is Regulated by Sonic Hedgehog. Endocrinology 2010, 151, 1119–1128. [Google Scholar] [CrossRef] [Green Version]
- Bandiera, R.; Vidal, V.P.I.; Motamedi, F.J.; Clarkson, M.; Sahut-Barnola, I.; von Gise, A.; Pu, W.T.; Hohenstein, P.; Martinez, A.; Schedl, A. WT1 maintains adrenal-gonadal primordium identity and marks a population of AGP-like progenitors within the adrenal gland. Dev. Cell 2013, 27, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Steenblock, C.; Rubin de Celis, M.F.; Delgadillo Silva, L.F.; Pawolski, V.; Brennand, A.; Werdermann, M.; Berger, I.; Santambrogio, A.; Peitzsch, M.; Andoniadou, C.L.; et al. Isolation and characterization of adrenocortical progenitors involved in the adaptation to stress. Proc. Natl. Acad. Sci. USA 2018, 115, 12997–13002. [Google Scholar] [CrossRef] [Green Version]
- Mitani, F. Functional zonation of the rat adrenal cortex: The development and maintenance. Proc. Japan Acad. Ser. B Phys. Biol. Sci. 2014, 90, 163–183. [Google Scholar] [CrossRef] [Green Version]
- Yaglova, N.V.; Obernikhin, S.S.; Nazimova, S.V.; Yaglov, V.V. Role of Transcription Factor Oct4 in Postnatal Development and Function of the Adrenal Cortex. Bull. Exp. Biol. Med. 2019, 167, 568–573. [Google Scholar] [CrossRef]
- Dumontet, T.; Sahut-Barnola, I.; Septier, A.; Montanier, N.; Plotton, I.; Roucher-Boulez, F.; Ducros, V.; Lefrançois-Martinez, A.-M.; Pointud, J.-C.; Zubair, M.; et al. PKA signaling drives reticularis differentiation and sexually dimorphic adrenal cortex renewal. JCI insight 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Sahut-Barnola, I.; de Joussineau, C.; Val, P.; Lambert-Langlais, S.; Damon, C.; Lefrançois-Martinez, A.M.; Pointud, J.C.; Marceau, G.; Sapin, V.; Tissier, F.; et al. Cushing’s syndrome and fetal features resurgence in adrenal cortex-specific Prkar1a knockout mice. PLoS Genet. 2010, 6, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Berthon, A.; Sahut-Barnola, I.; Lambert-Langlais, S.; de Joussineau, C.; Damon-Soubeyrand, C.; Louiset, E.; Taketo, M.M.; Tissier, F.; Bertherat, J.; Lefrançois-Martinez, A.-M.; et al. Constitutive β-catenin activation induces adrenal hyperplasia and promotes adrenal cancer development. Hum. Mol. Genet. 2010, 19, 1561–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levasseur, A.; St-Jean, G.; Paquet, M.; Boerboom, D.; Boyer, A. Targeted disruption of YAP and TAZ impairs the maintenance of the adrenal cortex. Endocrinology 2017, 158, 3738–3753. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Drelon, C.; Rodriguez, S.; Tabbal, H.; Septier, A.; Damon-Soubeyrand, C.; Dumontet, T.; Berthon, A.; Sahut-Barnola, I.; Djari, C.; et al. Steroidogenic differentiation and PKA signaling are programmed by histone methyltransferase EZH2 in the adrenal cortex. Proc. Natl. Acad. Sci. USA 2018, 115, E12265–E12274. [Google Scholar] [CrossRef] [Green Version]
- Borges, K.S.; Pignatti, E.; Leng, S.; Kariyawasam, D.; Ruiz-Babot, G.; Ramalho, F.S.; Taketo, M.M.; Carlone, D.L.; Breault, D.T. Wnt/β-catenin activation cooperates with loss of p53 to cause adrenocortical carcinoma in mice. Oncogene 2020, 39, 5282–5291. [Google Scholar] [CrossRef]
- Heaton, J.H.; Wood, M.A.; Kim, A.C.; Lima, L.O.; Barlaskar, F.M.; Almeida, M.Q.; Fragoso, M.C.B.V.; Kuick, R.; Lerario, A.M.; Simon, D.P.; et al. Progression to adrenocortical tumorigenesis in mice and humans through insulin-like growth factor 2 and β-catenin. Am. J. Pathol. 2012, 181, 1017–1033. [Google Scholar] [CrossRef]
- Trejter, M.; Jopek, K.; Celichowski, P.; Tyczewska, M.; Malendowicz, L.K.; Rucinski, M. Expression of estrogen, estrogen related and androgen receptors in adrenal cortex of intact adult male and female rats. Folia Histochem. Cytobiol. 2015, 53, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gannon, A.-L.; O’Hara, L.; Mason, J.I.; Jørgensen, A.; Frederiksen, H.; Milne, L.; Smith, S.; Mitchell, R.T.; Smith, L.B. Androgen receptor signalling in the male adrenal facilitates X-zone regression, cell turnover and protects against adrenal degeneration during ageing. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Bentvelsen, F.M.; Mcphaul, M.J.; Wilson, C.M.; Wilson, J.D.; George, F.W. Regulation of immunoreactive androgen receptor in the adrenal gland of the adult rat. Endocrinology 1996, 137, 2659–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutson, D.D.; Gurrala, R.; Ogola, B.O.; Zimmerman, M.A.; Mostany, R.; Satou, R.; Lindsey, S.H. Estrogen receptor profiles across tissues from male and female Rattus norvegicus. Biol. Sex Differ. 2019, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Maric, C.; Roesch, D.M.; Zheng, W.; Verbalis, J.G.; Sandberg, K. Estrogen regulates adrenal angiotensin AT1 receptors by modulating AT1 receptor translation. Endocrinology 2003, 144, 3251–3261. [Google Scholar] [CrossRef] [Green Version]
- Baquedano, M.S.; Saraco, N.; Berensztein, E.; Pepe, C.; Bianchini, M.; Levy, E.; Goñi, J.; Rivarola, M.A.; Belgorosky, A. Identification and developmental changes of aromatase and estrogen receptor expression in prepubertal and pubertal human adrenal tissues. J. Clin. Endocrinol. Metab. 2007, 92, 2215–2222. [Google Scholar] [CrossRef]
- Caroccia, B.; Seccia, T.M.; Campos, A.G.; Gioco, F.; Kuppusamy, M.; Ceolotto, G.; Guerzoni, E.; Simonato, F.; Mareso, S.; Lenzini, L.; et al. GPER-1 and estrogen receptor-β ligands modulate aldosterone synthesis. Endocrinology 2014, 155, 4296–4304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzon, L.; Masi, G.; Pacenti, M.; Trevisan, M.; Fallo, F.; Remo, A.; Martignoni, G.; Montanaro, D.; Pezzi, V.; Palù, G. Expression of aromatase and estrogen receptors in human adrenocortical tumors. Virchows Arch. 2008, 452, 181–191. [Google Scholar] [CrossRef]
- De Cremoux, P.; Rosenberg, D.; Goussard, J.; Brémont-Weil, C.; Tissier, F.; Tran-Perennou, C.; Groussin, L.; Bertagna, X.; Bertherat, J.; Raffin-Sanson, M.L. Expression of progesterone and estradiol receptors in normal adrenal cortex, adrenocortical tumors, and primary pigmented nodular adrenocortical disease. Endocr. Relat. Cancer 2008, 15, 465–474. [Google Scholar] [CrossRef]
- Colafella, K.M.M.; Denton, K.M. Sex-specific differences in hypertension and associated cardiovascular disease. Nat. Rev. Nephrol. 2018, 14, 185. [Google Scholar] [CrossRef] [PubMed]
- Roesch, D.M.; Tian, Y.; Zheng, W.; Shi, M.; Verbalis, J.G.; Sandberg, K. Estradiol attenuates angiotensin-induced aldosterone secretion in ovariectomized rats. Endocrinology 2000, 141, 4629–4636. [Google Scholar] [CrossRef] [PubMed]
- Macova, M.; Armando, I.; Zhou, J.; Baiardi, G.; Tyurmin, D.; Larrayoz-Roldan, I.M.; Saavedra, J.M. Estrogen reduces aldosterone, upregulates adrenal angiotensin II AT 2 receptors and normalizes adrenomedullary Fra-2 in ovariectomized rats. Neuroendocrinology 2008, 88, 276–286. [Google Scholar] [CrossRef] [Green Version]
- Yanes, L.L.; Romero, D.G. Dihydrotestosterone stimulates aldosterone secretion by H295R human adrenocortical cells. Mol. Cell. Endocrinol. 2009, 303, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heck, A.L.; Handa, R.J. Sex differences in the hypothalamic–pituitary–adrenal axis’ response to stress: An important role for gonadal hormones. Neuropsychopharmacology 2019, 44, 45–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, J.V.; Wood, S.A.; Atkinson, H.C.; Bate, E.; Lightman, S.L.; Ingram, C.D.; Jessop, D.S.; Harbuz, M.S. Gonadectomy reverses the sexually diergic patterns of circadian and stress-induced hypothalamic-pituitary-adrenal axis activity in male and female rats. J. Neuroendocrinol. 2004, 16, 516–524. [Google Scholar] [CrossRef]
- Seale, J.V.; Wood, S.A.; Atkinson, H.C.; Harbuz, M.S.; Lightman, S.L. Gonadal steroid replacement reverses gonadectomy-induced changes in the corticosterone pulse profile and stress-induced hypothalamic-pituitary-adrenal axis activity of male and female rats. J. Neuroendocrinol. 2004, 16, 989–998. [Google Scholar] [CrossRef]
- Figueiredo, H.F.; Ulrich-Lai, Y.M.; Choi, D.C.; Herman, J.P. Estrogen potentiates adrenocortical responses to stress in female rats. Am. J. Physiol. - Endocrinol. Metab. 2007, 292. [Google Scholar] [CrossRef]
- Nowak, K.W.; Neri, G.; Nussdorfer, G.G.; Malendowicz, L.K. Effects of sex hormones on the steroidogenic activity of dispersed adrenocortical cells of the rat adrenal cortex. Life Sci. 1995, 57, 833–837. [Google Scholar] [CrossRef]
- Benmouloud, A.; Amirat, Z.; Khammar, F.; Patchev, A.V.; Exbrayat, J.M.; Almeida, O.F.X. Androgen receptor-mediated regulation of adrenocortical activity in the sand rat, Psammomys obesus. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2014, 184, 1055–1063. [Google Scholar] [CrossRef]
- Young, E.A.; Altemus, M.; Parkison, V.; Shastry, S. Effects of estrogen antagonists and agonists on the ACTH response to restraint stress in female rats. Neuropsychopharmacology 2001, 25, 881–891. [Google Scholar] [CrossRef]
- van Keulen, B.J.; Dolan, C.V.; van der Voorn, B.; Andrew, R.; Walker, B.R.; Pol, H.H.; Boomsma, D.I.; Rotteveel, J.; Finken, M.J.J. Sexual dimorphism in cortisol metabolism throughout pubertal development: A longitudinal study. Endocr. Connect. 2020, 9, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Wudy, S.A.; Hartmann, M.F.; Remer, T. Sexual dimorphism in cortisol secretion starts after age 10 in healthy children: Urinary cortisol metabolite excretion rates during growth. Am. J. Physiol. Metab. 2007, 293, E970–E976. [Google Scholar] [CrossRef] [Green Version]
- Rubinow, D.R.; Roca, C.A.; Schmidt, P.J.; Danaceau, M.A.; Putnam, K.; Cizza, G.; Chrousos, G.; Nieman, L. Testosterone suppression of CRH-stimulated cortisol in men. Neuropsychopharmacology 2005, 30, 1906–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, M.A.C.; Mahon, P.B.; McCaul, M.E.; Wand, G.S. Hypothalamic-pituitary-adrenal axis response to acute psychosocial stress: Effects of biological sex and circulating sex hormones. Psychoneuroendocrinology 2016, 66, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, J.; Matsumoto, T.; Shiina, H.; Inoue, K.; Takada, I.; Ito, S.; Itoh, J.; Minematsu, T.; Sato, T.; Yanase, T.; et al. The Pituitary Function of Androgen Receptor Constitutes a Glucocorticoid Production Circuit. Mol. Cell. Biol. 2007, 27, 4807–4814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubair, M.; Parker, K.L.; Morohashi, K. Developmental Links between the Fetal and Adult Zones of the Adrenal Cortex Revealed by Lineage Tracing. Mol. Cell. Biol. 2008, 28, 7030–7040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasco, M.A. Telomere length, stem cells and aging. Nat. Chem. Biol. 2007, 3, 640–649. [Google Scholar] [CrossRef]
- Bayne, S.; Jones, M.E.; Li, H.; Pinto, A.R.; Simpson, E.R.; Liu, J.-P. Estrogen deficiency leads to telomerase inhibition, telomere shortening and reduced cell proliferation in the adrenal gland of mice. Cell Res. 2008, 18, 1141–1150. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, K.; Aida, J.; Takubo, K.; Yamazaki, Y.; Gao, X.; Komatsu, A.; Takakuma, S.; Kakizaki, M.; Inoshita, N.; Gomi, F.; et al. Correlation Between Telomere Attrition of Zona Fasciculata and Adrenal Weight Reduction in Older Men. J. Clin. Endocrinol. Metab. 2019, 105, e200–e210. [Google Scholar] [CrossRef]
- Nonaka, K.; Aida, J.; Takubo, K.; Yamazaki, Y.; Takakuma, S.; Kakizaki, M.; Matsuda, Y.; Ishikawa, N.; Ishiwata, T.; Chong, J.M.; et al. Correlation between Differentiation of Adrenocortical Zones and Telomere Lengths Measured by Q-FISH. J. Clin. Endocrinol. Metab. 2019, 104, 5642–5650. [Google Scholar] [CrossRef]
- Rossi, R.; Zatelli, M.C.; Valentini, A.; Cavazzini, P.; Fallo, F.; Del Senno, L.; Uberti, E.C.D. Evidence for androgen receptor gene expression and growth inhibitory effect of dihydrotestosterone on human adrenocortical cells. J. Endocrinol. 1998, 159, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Montanaro, D.; Maggiolini, M.; Recchia, A.G.; Sirianni, R.; Aquila, S.; Barzon, L.; Fallo, F.; Andò, S.; Pezzi, V. Antiestrogens upregulate estrogen receptor β expression and inhibit adrenocortical H295R cell proliferation. J. Mol. Endocrinol. 2005, 35, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Sirianni, R.; Zolea, F.; Chimento, A.; Ruggiero, C.; Cerquetti, L.; Fallo, F.; Pilon, C.; Arnaldi, G.; Carpinelli, G.; Stigliano, A.; et al. Targeting estrogen receptor-α reduces adrenocortical cancer (ACC) cell growth in vitro and in vivo: Potential therapeutic role of selective estrogen receptor modulators (SERMs) for ACC treatment. J. Clin. Endocrinol. Metab. 2012, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, F.; Mittre, H.; Benhaim, A.; Bois, C.; Bertherat, J.; Carreau, S.; Reznik, Y. Aromatase expression in the normal human adult adrenal and in adrenocortical tumors: Biochemical, immunohistochemical, and molecular studies. Eur. J. Endocrinol. 2009, 160, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyraki, R.; Schedl, A. The Sexually Dimorphic Adrenal Cortex: Implications for Adrenal Disease. Int. J. Mol. Sci. 2021, 22, 4889. https://doi.org/10.3390/ijms22094889
Lyraki R, Schedl A. The Sexually Dimorphic Adrenal Cortex: Implications for Adrenal Disease. International Journal of Molecular Sciences. 2021; 22(9):4889. https://doi.org/10.3390/ijms22094889
Chicago/Turabian StyleLyraki, Rodanthi, and Andreas Schedl. 2021. "The Sexually Dimorphic Adrenal Cortex: Implications for Adrenal Disease" International Journal of Molecular Sciences 22, no. 9: 4889. https://doi.org/10.3390/ijms22094889
APA StyleLyraki, R., & Schedl, A. (2021). The Sexually Dimorphic Adrenal Cortex: Implications for Adrenal Disease. International Journal of Molecular Sciences, 22(9), 4889. https://doi.org/10.3390/ijms22094889