
Network Pharmacology Analysis and Experimental Validation of Kaempferol in the Treatment of Ischemic Stroke by Inhibiting Apoptosis and Regulating Neuroinflammation Involving Neutrophils


Abstract
:1. Introduction
2. Results
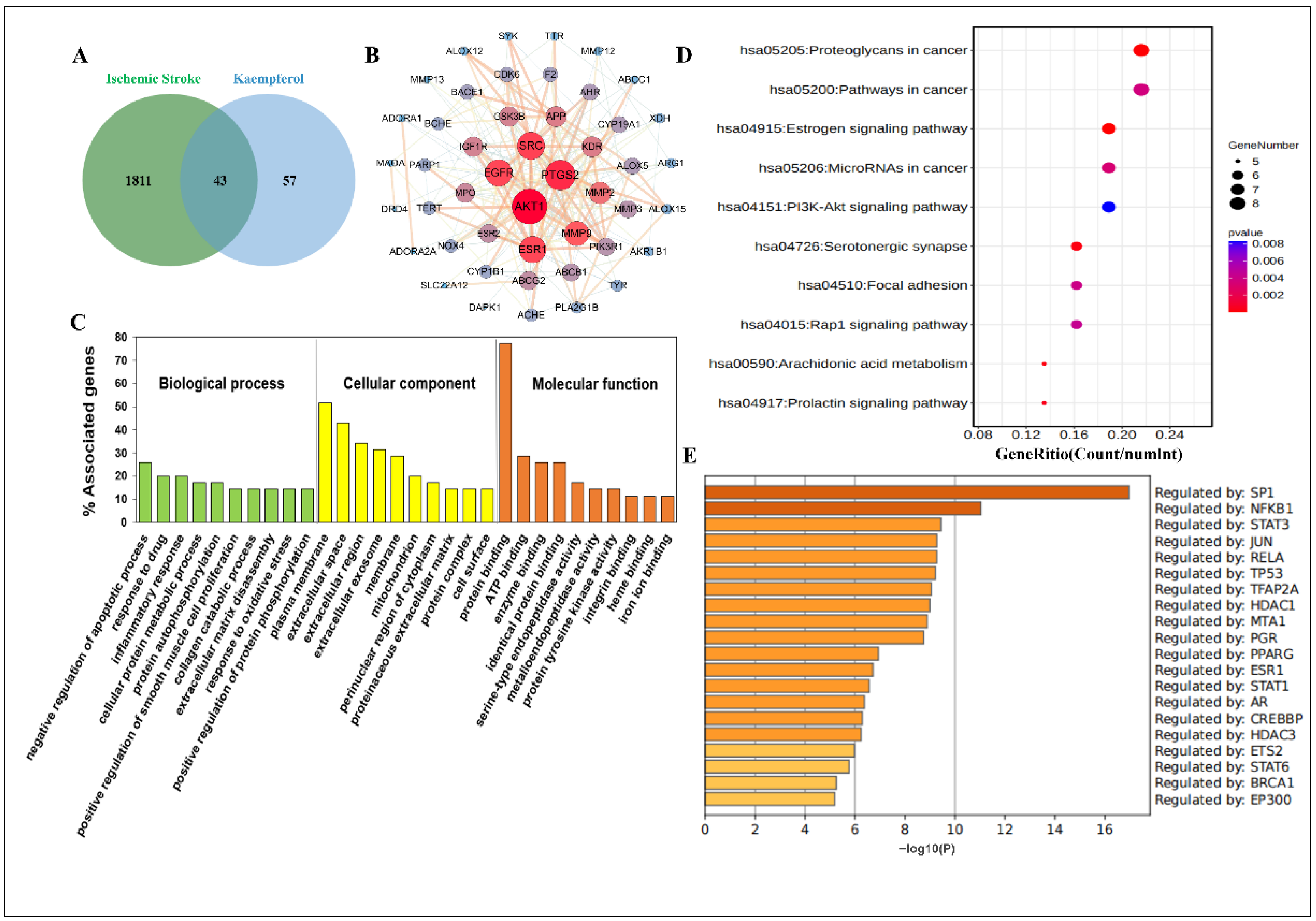
2.1. Network Pharmacology Predicted the Biological Process and Enrichment Analysis of Kaempferol against Ischemic Stroke
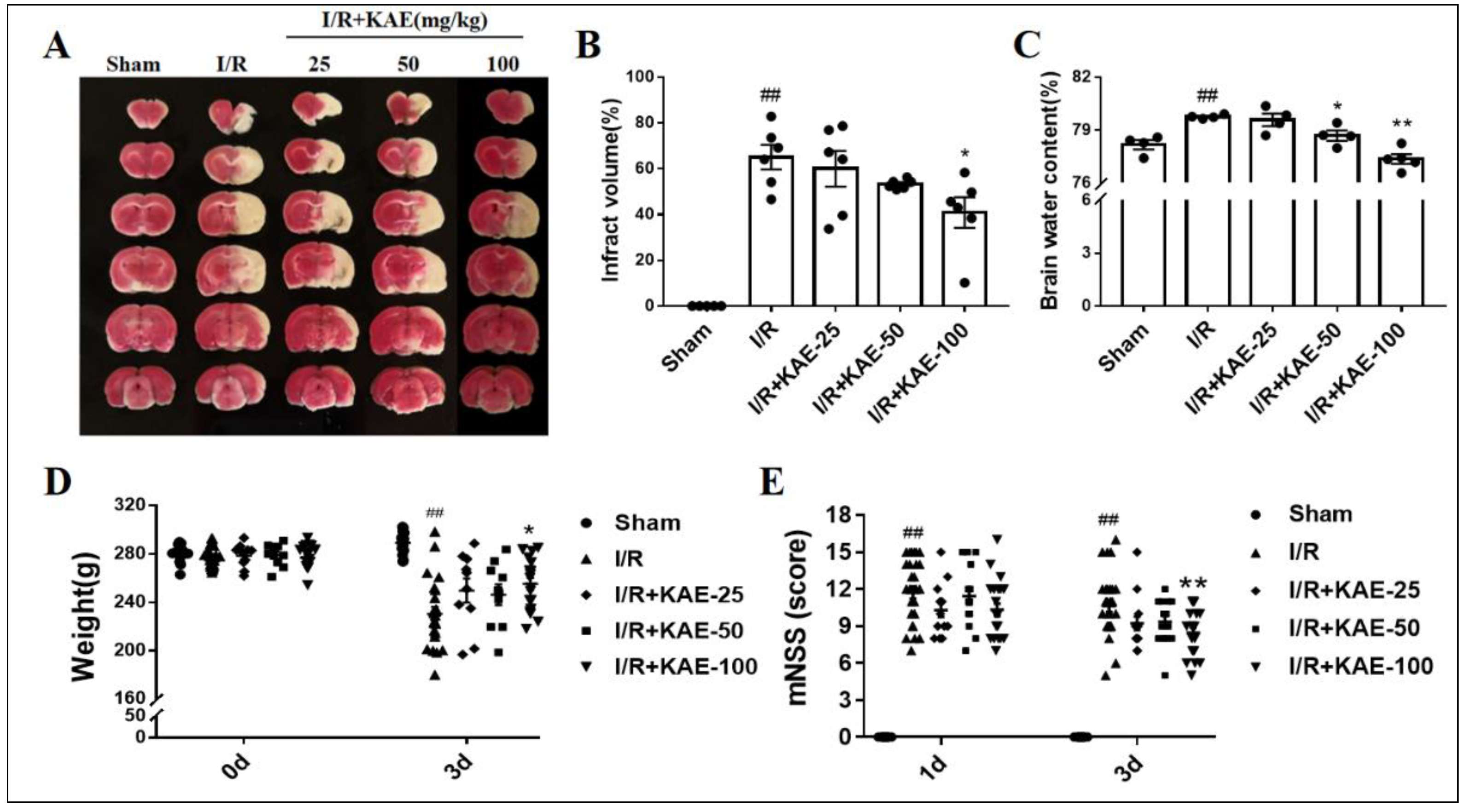
2.2. Effect of Kaempferol against Transient Focal Cerebral I/R Injury
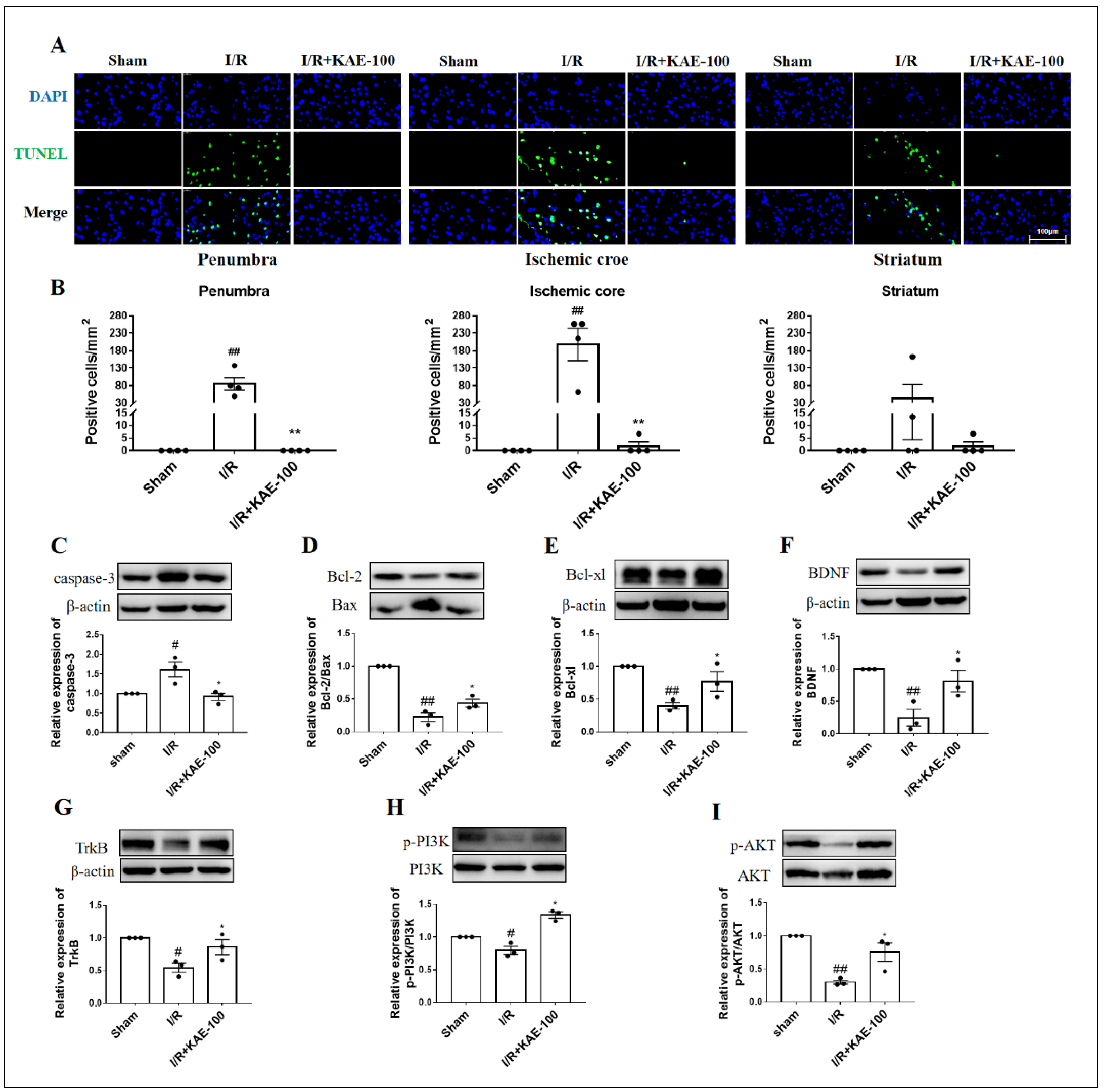
2.3. Kaempferol Attenuated Cell Apoptosis in Cerebral I/R Rats
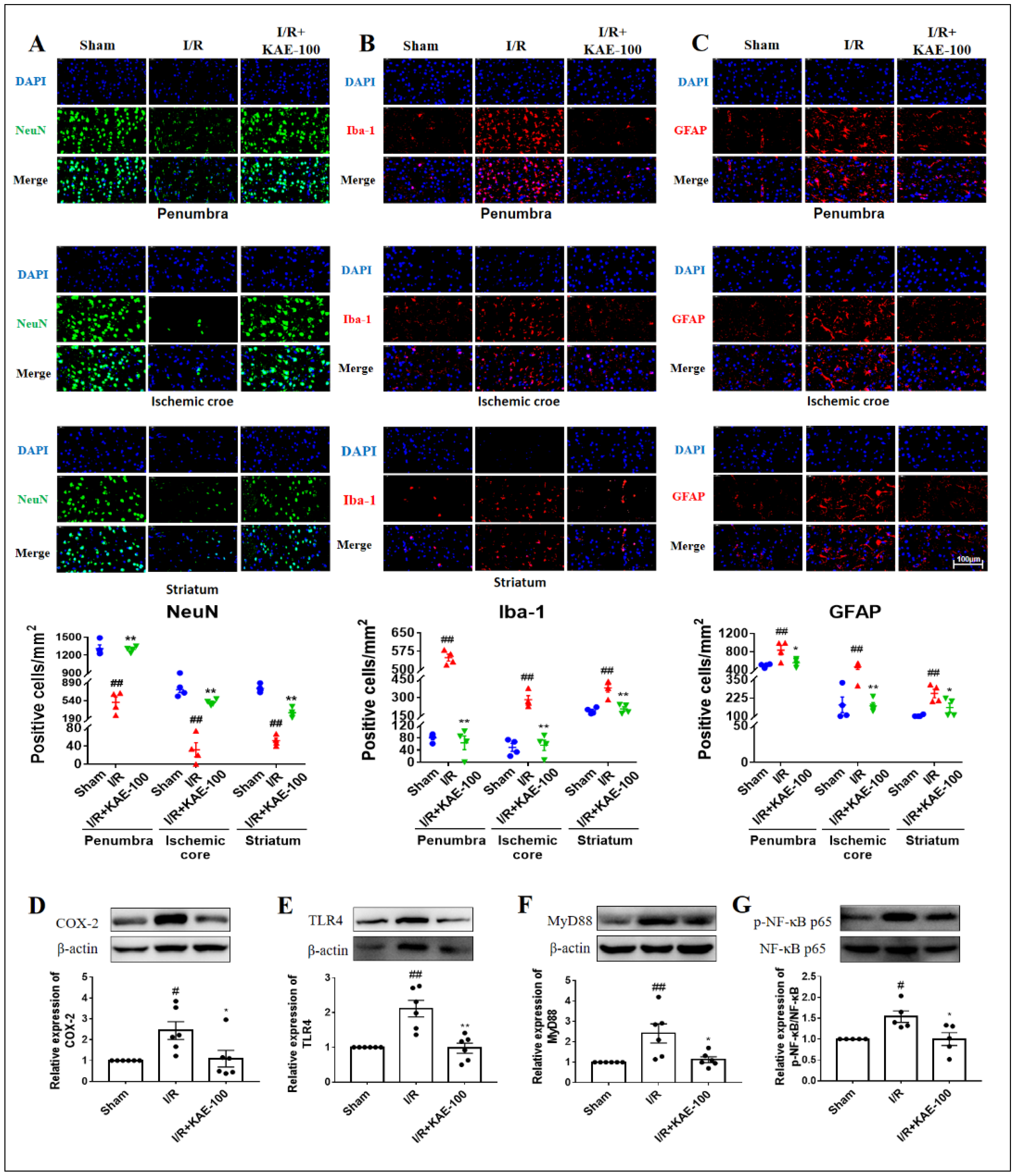
2.4. Kaempferol Reduced Neuron Loss as Well as the Activation of Microglia and Astrocytes in Cerebral I/R Rats
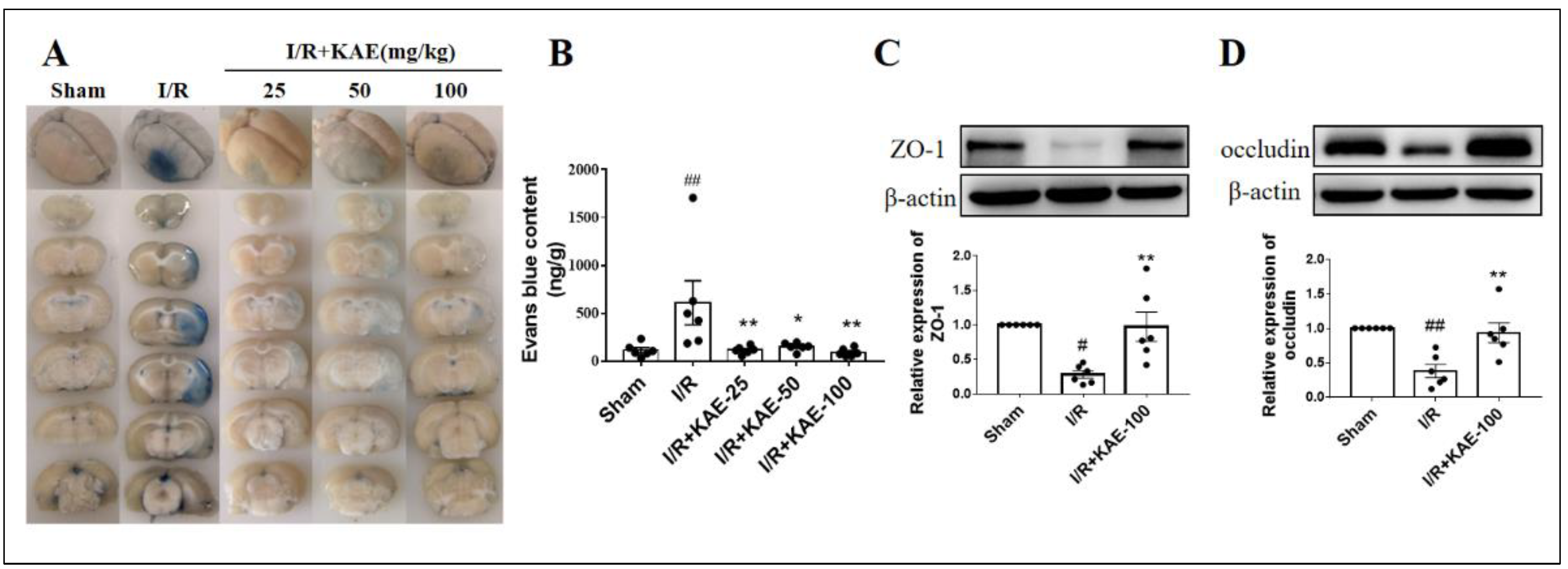
2.5. Kaempferol Alleviated Blood Brain Barrier Disruption in Cerebral I/R Rats
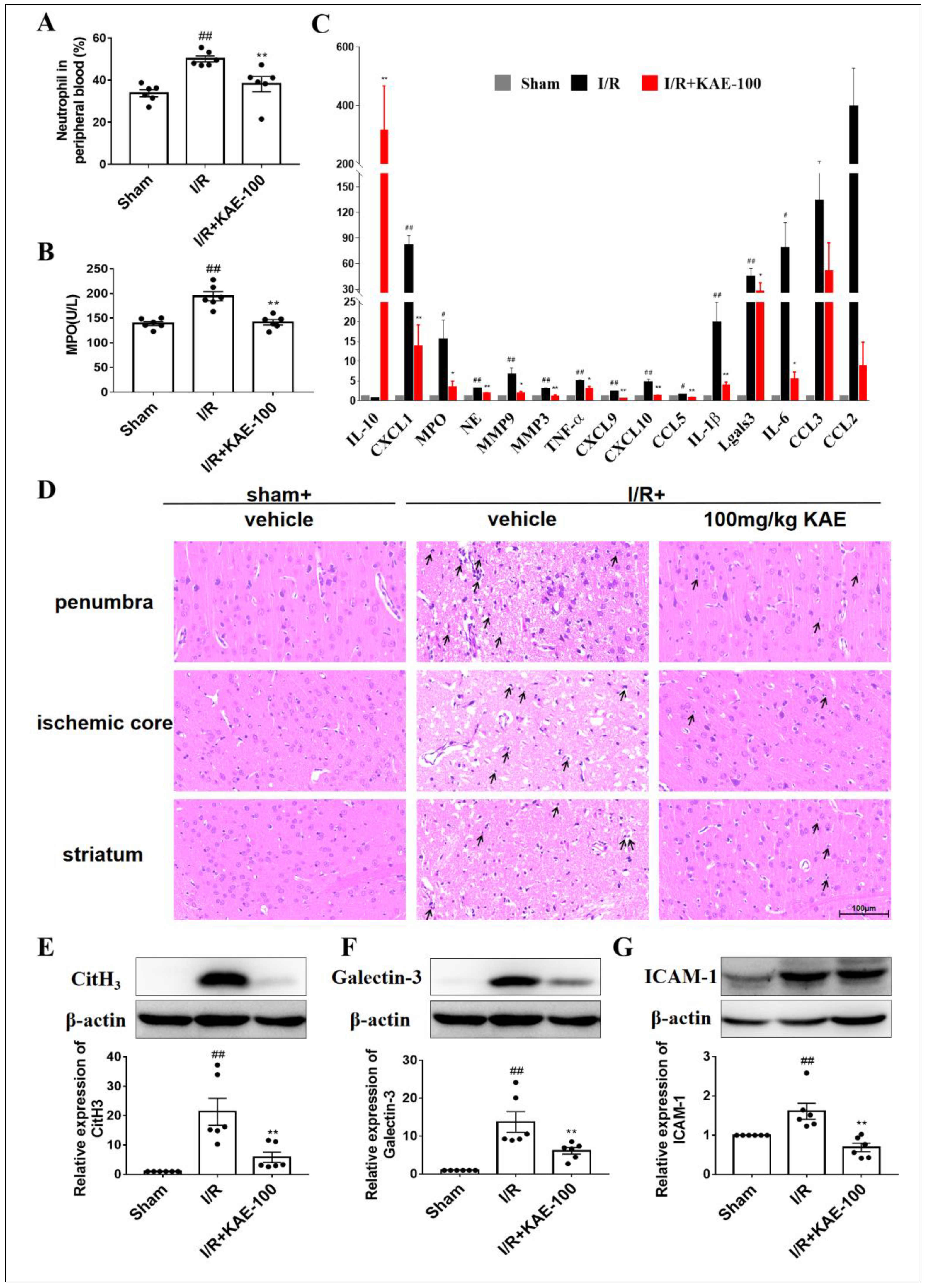
2.6. Kaempferol Inhibited Neutrophil Activation, Aggregation and Infiltration in Cerebral I/R Rats
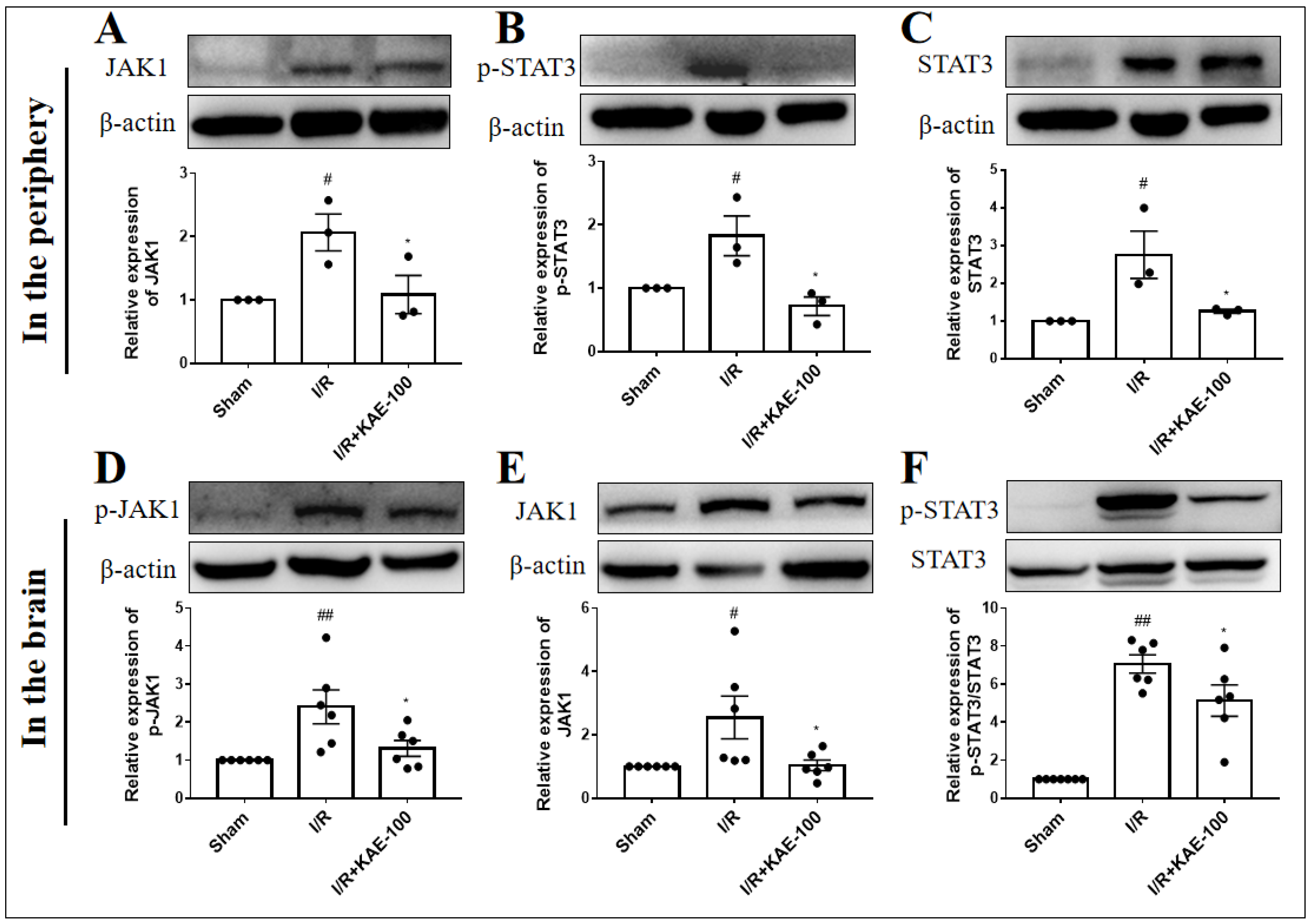
2.7. Kaempferol Down-Regulated the Expression of JAK1/STAT3 Signaling Pathway in Peripheral Blood Neutrophils and Ischemic Cortex
3. Discussion
4. Materials and Methods
4.1. Network Pharmacology Analysis
4.1.1. Prediction of Drug-Related Targets
4.1.2. Collection of Targets of Ischemic Stroke
4.1.3. PPI Network Construction
4.1.4. GO, KEGG Pathway and TRRUST Enrichment Analysis
4.2. Reagents and Antibody
4.3. Animals and Experiment Design
4.4. The mNSS Test
4.5. Infarct Volume Assessments
4.6. Brain Edema Detection
4.7. TUNEL Staining
4.8. Immunofluorescence Detection of NeuN, Iba-1 and GFAP
4.9. Evans Blue (EB) Leakage
4.10. Hematoxylin-Eosin (HE) Staining
4.11. Determination of Myeloperoxidase (MPO) Activity
4.12. Peripheral Blood Count
4.13. Blood Neutrophil Isolation
4.14. Western Blotting Analysis
4.15. Real-Time PCR
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khamchai, S.; Chumboatong, W.; Hata, J.; Tocharus, C.; Suksamrarn, A.; Tocharus, J. Morin protects the blood-brain barrier integrity against cerebral ischemia reperfusion through anti-inflammatory actions in rats. Sci. Rep. 2020, 10, 13379. [Google Scholar] [CrossRef]
- Chen, G.; Ye, X.; Zhang, J.; Tang, T.; Li, L.; Lu, P.; Wu, Q.; Yu, B.; Kou, J. Limb Remote Ischemic Postconditioning Reduces Ischemia-Reperfusion Injury by Inhibiting NADPH Oxidase Activation and MyD88-TRAF6-P38MAP-Kinase Pathway of Neutrophils. Int. J. Mol. Sci. 2016, 17, 1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, D.L.; Gajghate, S.; Reichenbach, N.L.; Winfield, M.; Persidsky, Y.; Heldt, N.A.; Rom, S. let-7g counteracts endothelial dysfunction and ameliorating neurological functions in mouse ischemia/reperfusion stroke model. Brain Behav. Immun. 2020, 87, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Tang, Y.; An, R.; Lin, M.; Chen, L.; Du, J. RTN1-C mediates cerebral ischemia/reperfusion injury via ER stress and mitochondria-associated apoptosis pathways. Cell Death Dis. 2017, 8, e3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, H.; Li, S.; Zhang, M. Berberine Ameliorates MCAO Induced Cerebral Ischemia/Reperfusion Injury via Activation of the BDNF-TrkB-PI3K/Akt Signaling Pathway. Neurochem. Res. 2018, 43, 702–710. [Google Scholar] [CrossRef]
- Wang, S.; Xu, H.; Xin, Y.; Li, M.; Fu, W.; Wang, Y.; Lu, Z.; Yu, X.; Sui, D. Neuroprotective effects of Kaempferide-7-O-(4″-O-acetylrhamnosyl)-3-O-rutinoside on cerebral ischemia-reperfusion injury in rats. Eur. J. Pharmacol. 2016, 788, 335–342. [Google Scholar] [CrossRef]
- Uzdensky, A.B. Apoptosis regulation in the penumbra after ischemic stroke: Expression of pro- and antiapoptotic proteins. Apoptosis 2019, 24, 687–702. [Google Scholar] [CrossRef]
- Chang, C.Y.; Kao, T.K.; Chen, W.Y.; Ou, Y.C.; Li, J.R.; Liao, S.L.; Raung, S.L.; Chen, C.J. Tetramethylpyrazine inhibits neutrophil activation following permanent cerebral ischemia in rats. Biochem. Biophys. Res. Commun. 2015, 463, 421–427. [Google Scholar] [CrossRef]
- Kim, S.W.; Lee, H.; Lee, H.K.; Kim, I.D.; Lee, J.K. Neutrophil extracellular trap induced by HMGB1 exacerbates damages in the ischemic brain. Acta Neuropathol. Commun. 2019, 7, 131. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Zhao, H.; Li, G.; Zhang, S.; Wang, R.; Tao, Z.; Zheng, Y.; Han, Z.; Liu, P.; Ma, Q.; et al. Intravenous antagomiR-494 lessens brain-infiltrating neutrophils by increasing HDAC2-mediated repression of multiple MMPs in experimental stroke. FASEB J. 2020, 34, 6934–6949. [Google Scholar] [CrossRef]
- Guo, A.J.; Choi, R.C.; Zheng, K.Y.; Chen, V.P.; Dong, T.T.; Wang, Z.T.; Vollmer, G.; Lau, D.T.; Tsim, K.W. Kaempferol as a flavonoid induces osteoblastic differentiation via estrogen receptor signaling. Chin. Med. 2012, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva Dos Santos, J.; Gonçalves Cirino, J.P.; de Oliveira Carvalho, P.; Ortega, M.M. The Pharmacological Action of Kaempferol in Central Nervous System Diseases: A Review. Front. Pharmacol. 2020, 11, 565700. [Google Scholar] [CrossRef] [PubMed]
- López-Sánchez, C.; Martín-Romero, F.J.; Sun, F.; Luis, L.; Samhan-Arias, A.K.; García-Martínez, V.; Gutiérrez-Merino, C. Blood micromolar concentrations of kaempferol afford protection against ischemia/reperfusion-induced damage in rat brain. Brain Res. 2007, 1182, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Cheng, X.; Yang, Y.L.; Liu, M.; Zhang, S.S.; Wang, Y.H.; Du, G.H. Kaempferol attenuates neuroinflammation and blood brain barrier dysfunction to improve neurological deficits in cerebral ischemia/reperfusion rats. Brain Res. 2019, 1722, 146361. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Luo, H.; Zhou, X.; Cheng, C.Y.; Lin, L.; Liu, B.L.; Liu, K.; Li, P.; Yang, H. Succinate-induced neuronal mitochondrial fission and hexokinase II malfunction in ischemic stroke: Therapeutical effects of kaempferol. Biochim. Biophys Acta Mol. Basis Dis. 2017, 1863, 2307–2318. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wei, X.; Hou, Y.; Liu, X.; Li, S.; Sun, B.; Liu, X.; Liu, H. Tetramethylpyrazine analogue CXC195 protects against cerebral ischemia/reperfusion-induced apoptosis through PI3K/Akt/GSK3β pathway in rats. Neurochem. Int. 2014, 66, 27–32. [Google Scholar] [CrossRef]
- Seol, S.I.; Kim, H.J.; Choi, E.B.; Kang, I.S.; Lee, H.K.; Lee, J.K.; Kim, C. Taurine Protects against Postischemic Brain Injury via the Antioxidant Activity of Taurine Chloramine. Antioxidants. 2021, 10, 372. [Google Scholar] [CrossRef]
- Cai, W.; Wang, J.; Hu, M.; Chen, X.; Lu, Z.; Bellanti, J.A.; Zheng, S.G. All trans-retinoic acid protects against acute ischemic stroke by modulating neutrophil functions through STAT1 signaling. J. Neuroinflammation 2019, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Mao, J.; Wang, R.; Li, S.; Wu, B.; Yuan, Y. Kaempferol Protects Against Cerebral Ischemia Reperfusion Injury Through Intervening Oxidative and Inflammatory Stress Induced Apoptosis. Front. Pharmacol. 2020, 11, 424. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Tu, M.; Gao, W.; Cai, X.; Song, F.; Chen, Z.; Zhang, Q.; Wang, J.; Jin, C.; Shi, J.; et al. Hollow Prussian Blue Nanozymes Drive Neuroprotection against Ischemic Stroke via Attenuating Oxidative Stress. Count. Inflamm. Suppressing Cell Apoptosis 2019, 19, 2812–2823. [Google Scholar]
- Yang, J.L.; Mukda, S.; Chen, S.D. Diverse roles of mitochondria in ischemic stroke. Redox Biol. 2018, 16, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Khoshnam, S.E.; Winlow, W.; Farzaneh, M.; Farbood, Y.; Moghaddam, H.F. Pathogenic mechanisms following ischemic stroke. Neurol. Sci. 2017, 38, 1167–1186. [Google Scholar] [CrossRef] [PubMed]
- Djordje Radaka, N.K.; Ivana Resanovicc, A.J.; Emina Sudar-Milovanovicc, S.Z.; Isenovicc, S.A.M.a.E.R. Apoptosis and Acute Brain Ischemia in Ischemic Stroke. Curr. Vasc. Pharmacol. 2017, 15, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Yue, J.K.; Zusman, B.E.; Nwachuku, E.L.; Puccio, A.M. B-Cell Lymphoma 2 (Bcl-2) and Regulation of Apoptosis after Traumatic Brain Injury: A Clinical Perspective. Medicina 2020, 56, 300. [Google Scholar] [CrossRef]
- Xie, C.J.; Gu, A.P.; Cai, J.; Wu, Y.; Chen, R.C. Curcumin protects neural cells against ischemic injury in N2a cells and mouse brain with ischemic stroke. Brain Behav. 2018, 8, e00921. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Kim, C.J.; Shin, M.S.; Lim, B.V. Treadmill exercise ameliorates memory impairment through ERK-Akt-CREB-BDNF signaling pathway in cerebral ischemia gerbils. J. Exerc. Rehabil. 2020, 16, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xiang, L.; Wang, C.; Song, Y.; Miao, J.; Miao, M. Protection against acute cerebral ischemia/reperfusion injury by Leonuri Herba Total Alkali via modulation of BDNF-TrKB-PI3K/Akt signaling pathway in rats. Biomed. Pharm. 2021, 133, 111021. [Google Scholar] [CrossRef]
- Zhong, J.B.; Li, X.; Zhong, S.M.; Liu, J.D.; Chen, C.B.; Wu, X.Y. Knockdown of long noncoding antisense RNA brain-derived neurotrophic factor attenuates hypoxia/reoxygenation-induced 5nerve cell apoptosis through the BDNF-TrkB-PI3K/Akt signaling pathway. Neuroreport 2017, 28, 910–916. [Google Scholar] [CrossRef]
- Yun, Y.C.; Jang, D.; Yoon, S.B.; Kim, D.; Choi, D.H.; Kwon, O.S.; Lee, Y.M.; Youn, D. Laser Acupuncture Exerts Neuroprotective Effects via Regulation of Creb, Bdnf, Bcl-2, and Bax Gene Expressions in the Hippocampus. Evid. Based Complement. Alternat. Med. 2017, 2017, 7181637. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Yang, Y.L.; Yang, H.; Wang, Y.H.; Du, G.H. Kaempferol alleviates LPS-induced neuroinflammation and BBB dysfunction in mice via inhibiting HMGB1 release and down-regulating TLR4/MyD88 pathway. Int. Immunopharmacol. 2018, 56, 29–35. [Google Scholar] [CrossRef]
- Yang, Y.L.; Cheng, X.; Li, W.H.; Liu, M.; Wang, Y.H.; Du, G.H. Kaempferol Attenuates LPS-Induced Striatum Injury in Mice Involving Anti-Neuroinflammation, Maintaining BBB Integrity, and Down-Regulating the HMGB1/TLR4 Pathway. Int. J. Mol. Sci. 2019, 20, 491. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.S.; Wang, D.; Jiang, Y.H.; Ma, H.; Li, X.Q.; Wang, H. Dexmedetomidine postconditioning alleviates spinal cord ischemia-reperfusion injury in rats via inhibiting neutrophil Infiltration, microglia activation, reactive gliosis and CXCL13/CXCR5 axis activation. Int. J. Neurosci. 2021, 11, 1–13. [Google Scholar]
- Chen, S.; Dong, Z.; Cheng, M.; Zhao, Y.; Wang, M.; Sai, N.; Wang, X.; Liu, H.; Huang, G.; Zhang, X. Homocysteine exaggerates microglia activation and neuroinflammation through microglia localized STAT3 overactivation following ischemic stroke. J. Neuroinflammation 2017, 14, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S. Dynamic reactive astrocytes after focal ischemia. Neural Regen Res. 2014, 9, 2048–2052. [Google Scholar] [CrossRef]
- Zhang, S.S.; Liu, M.; Liu, D.N.; Yang, Y.L.; Du, G.H.; Wang, Y.H. TLR4-IN-C34 Inhibits Lipopolysaccharide-Stimulated Inflammatory Responses via Downregulating TLR4/MyD88/NF-κB/NLRP3 Signaling Pathway and Reducing ROS Generation in BV2 Cells. Inflammation 2021, 45, 838–850. [Google Scholar] [CrossRef]
- Mo, Y.; Sun, Y.Y.; Liu, K.Y. Autophagy and inflammation in ischemic stroke. Neural. Regen Res. 2020, 15, 1388–1396. [Google Scholar]
- Jickling, G.C.; Liu, D.; Ander, B.P.; Stamova, B.; Zhan, X.; Sharp, F.R. Targeting neutrophils in ischemic stroke: Translational insights from experimental studies. J. Cereb. Blood Flow Metab. 2015, 35, 888–901. [Google Scholar] [CrossRef] [Green Version]
- Loria, V.; Dato, I.; Graziani, F.; Biasucci, L.M. Myeloperoxidase: A New Biomarker of Inflammation in Ischémie Heart Disease and Acute Coronary Syndromes. Mediat. Inflamm. 2008, 2008, 135625. [Google Scholar]
- Bian, Y.; Lei, J.; Zhong, J.; Wang, B.; Wan, Y.; Li, J.; Liao, C.; He, Y.; Liu, Z.; Ito, K.; et al. Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J. Nutr. Biochem. 2022, 99, 108840. [Google Scholar] [CrossRef]
- Yang, C.; Yang, W.; He, Z.; He, H.; Yang, X.; Lu, Y.; Li, H. Kaempferol Improves Lung Ischemia-Reperfusion Injury via Antiinflammation and Antioxidative Stress Regulated by SIRT1/HMGB1/NF-κB Axis. Front. Pharmacol. 2019, 10, 1635. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Li, C.; Yin, M.; Lin, J.; Zhang, L.; Li, N.; Jiang, N.; Xu, Q.; Wang, Q.; Gu, L.; et al. Kaempferol ameliorate the prognosis of Aspergillus fumigatus keratitis by reducing fungal load and inhibiting the Dectin-1 and p38 MAPK pathway. Exp. Eye Res. 2022, 216, 108960. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, X.; Xuan, Y.; Ying, J.; Fei, Y.; Rong, J.; Zhang, Y.; Zhang, J.; Liu, C.; Liu, Z. Kaempferol protects ethanol-induced gastric ulcers in mice via pro-inflammatory cytokines and NO. Acta Biochim. Biophys. Sin. 2018, 50, 246–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagami, S.; Tamura, M.; Hayashi, M.; Endo, N.; Tanabe, H.; Katsuura, Y.; Komoriya, K. Differential production of MCP-1 and cytokine-induced neutrophil chemoattractant in the ischemic brain after transient focal ischemia in rats. J. Leukoc. Biol. 1999, 65, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Dumbuya, J.S.; Chen, L.; Shu, S.Y.; Ma, L.; Luo, W.; Li, F.; Wu, J.Y.; Wang, B. G-CSF attenuates neuroinflammation and neuronal apoptosis via the mTOR/p70SK6 signaling pathway in neonatal Hypoxia-Ischemia rat model. Brain Res. 2020, 1739, 146817. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, A.P.; Diez, D.; Miranda-Saavedra, D. The IL-10/STAT3-mediated anti-inflammatory response: Recent developments and future challenges. Brief. Funct. Genomics 2013, 12, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Zhu, Y.; Hou, X.; Chen, W.; Qu, X.; Zhang, Y.; Li, Z.; Wang, C.; Chen, J.; Lv, L.; et al. NETs Lead to Sympathetic Hyperactivity After Traumatic Brain Injury Through the LL37-Hippo/MST1 Pathway. Front. Neurosci. 2021, 15, 621477. [Google Scholar] [CrossRef]
- Díaz-Alvarez, L.; Ortega, E. The Many Roles of Galectin-3, a Multifaceted Molecule, in Innate Immune Responses against Pathogens. Mediators Inflamm. 2017, 2017, 9247574. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Qu, Z.; Wang, H.; Su, Y.; Wang, Y.; Zhang, W.; Wang, Z.; Xu, Q. The effect of hydroxy safflower yellow A on coronary heart disease through Bcl-2/Bax and PPAR-γ. Exp. Ther. Med. 2018, 15, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Cai, F.; Li, C.R.; Wu, J.L.; Chen, J.G.; Liu, C.; Min, Q.; Yu, W.; Ouyang, C.-H.; Chen, J.H. Theaflavin ameliorates cerebral ischemia-reperfusion injury in rats through its anti-inflammatory effect and modulation of STAT-1. Mediat. Inflamm. 2006, 2006, 30490. [Google Scholar] [CrossRef] [Green Version]
- Panopoulos, A.D.; Zhang, L.; Snow, J.W.; Jones, D.M.; Smith, A.M.; El Kasmi, K.C.; Liu, F.; Goldsmith, M.A.; Link, D.C.; Murray, P.J.; et al. STAT3 governs distinct pathways in emergency granulopoiesis and mature neutrophils. Blood 2006, 108, 3682–3690. [Google Scholar] [CrossRef] [Green Version]
- de Groot, R.P.; Coffer, P.J.; Koenderman, L. Regulation of proliferation, differentiation and survival by the IL-3/IL-5/GM-CSF receptor family. Cell. Signal. 1998, 10, 619–628. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, J.J.; Murray, P.J. Cytokine signaling modules in inflammatory responses. Immunity 2008, 28, 477–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, A.; Das, A.S.; Sharma, M.; Pathak, M.P.; Chattopadhyay, P.; Biswas, K.; Mukhopadhyay, R. STAT3 and NF-κB are common targets for kaempferol-mediated attenuation of COX-2 expression in IL-6-induced macrophages and carrageenan-induced mouse paw edema. Biochem. Biophys. Rep. 2017, 12, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Cortes, J.R.; Perez-G, M.; Rivas, M.D.; Zamorano, J. Kaempferol inhibits IL-4-induced STAT6 activation by specifically targeting JAK3. J. Immunol. 2007, 179, 3881–3887. [Google Scholar] [CrossRef] [PubMed]
- Che, N.; Ma, Y.; Xin, Y. Protective Role of Fucoidan in Cerebral Ischemia-Reperfusion Injury through Inhibition of MAPK Signaling Pathway. Biomol. Ther. 2017, 25, 272–278. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Yang, Y.L.; Li, W.H.; Liu, M.; Zhang, S.S.; Wang, Y.H.; Du, G.H. Dynamic Alterations of Brain Injury, Functional Recovery, and Metabolites Profile after Cerebral Ischemia/Reperfusion in Rats Contributes to Potential Biomarkers. J. Mol. Neurosci. 2020, 70, 667–676. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, C.; Ji, Y.; Teng, L.; Guo, Y. Neuregulin-1β Plays a Neuroprotective Role by Inhibiting the Cdk5 Signaling Pathway after Cerebral Ischemia-Reperfusion Injury in Rats. J. Mol. Neurosci. 2018, 66, 261–272. [Google Scholar] [CrossRef]
- Lee, H.K.; Park, J.Y.; Lee, H.; Kim, I.D.; Kim, S.W.; Yoon, S.H.; Lee, J.K. Anti-Inflammatory and Neuroprotective Effects of DIPOPA (N,N-Diisopropyl-2-Oxopropanamide), an Ethyl Pyruvate Bioisoster, in the Postischemic Brain. Neurotherapeutics 2019, 16, 523–537. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.T.; Zhang, P.; Gao, Y.; Li, C.L.; Wang, H.J.; Chen, L.C.; Feng, Y.; Li, R.Y.; Li, Y.L.; Jiang, C.L. Early VEGF inhibition attenuates blood-brain barrier disruption in ischemic rat brains by regulating the expression of MMPs. Mol. Med. Rep. 2017, 15, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Zuo, X.; Lu, J.; Manaenko, A.; Qi, X.; Tang, J.; Mei, Q.; Xia, Y.; Hu, Q. MicroRNA-132 attenuates cerebral injury by protecting blood-brain-barrier in MCAO mice. Exp. Neurol. 2019, 316, 12–19. [Google Scholar] [CrossRef]
- Yang, Y.L.; Liu, M.; Cheng, X.; Li, W.H.; Zhang, S.S.; Wang, Y.H.; Du, G.H. Myricitrin blocks activation of NF-κB and MAPK signaling pathways to protect nigrostriatum neuron in LPS-stimulated mice. J. Neuroimmunol. 2019, 337, 577049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Song, J.; Li, W.; Kong, D.; Liang, Y.; Zhao, X.; Du, G. Salvianolic Acid D Alleviates Cerebral Ischemia-Reperfusion Injury by Suppressing the Cytoplasmic Translocation and Release of HMGB1-Triggered NF-κB Activation to Inhibit Inflammatory Response. Mediators Inflamm. 2020, 2020, 9049614. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Yin, J.; Ge, M.; Wang, S.; Xie, L.; Li, Y.; Si, J.Q.; Ma, K. Isoflurane Post-conditioning Ameliorates Cerebral Ischemia/Reperfusion Injury by Enhancing Angiogenesis Through Activating the Shh/Gli Signaling Pathway in Rats. Front. Neurosci. 2019, 13, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.H.; Yang, Y.L.; Cheng, X.; Liu, M.; Zhang, S.S.; Wang, Y.H.; Du, G.H. Baicalein attenuates caspase-independent cells death via inhibiting PARP-1 activation and AIF nuclear translocation in cerebral ischemia/reperfusion rats. Apoptosis 2020, 25, 354–369. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, J.; Cai, J.; Qiu, Y.; Zheng, H.; Lai, X.; Sui, X.; Wang, Y.; Lu, Q.; Zhang, Y.; et al. Targeted homing of CCR2-overexpressing mesenchymal stromal cells to ischemic brain enhances post-stroke recovery partially through PRDX4-mediated blood-brain barrier preservation. Theranostics 2018, 8, 5929–5944. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Chen, Q. Resveratrol attenuates brain damage in permanent focal cerebral ischemia via activation of PI3K/Akt signaling pathway in rats. Neurol. Res. 2018, 40, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Ren, Y.S.; Zhang, C.X.; Yue, W.B.; Lei, F.L. Effects of organic selenium (Se-enriched yeast) supplementation in gestation diet on antioxidant status, hormone profile and haemato-biochemical parameters in Taihang Black Goats. Anim. Feed Sci. Technol. 2018, 238, 57–65. [Google Scholar]
- Liu, M.; Zhang, S.S.; Liu, D.N.; Yang, Y.L.; Wang, Y.H.; Du, G.H. Chrysomycin A Attenuates Neuroinflammation by Down-Regulating NLRP3/Cleaved Caspase-1 Signaling Pathway in LPS-Stimulated Mice and BV2 Cells. Int. J. Mol. Sci. 2021, 22, 6799. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Lu, D.; Duan, S.F.; Gao, Y.R.; Liu, S.Y.; Hong, Y.; Dong, P.Z.; Chen, Y.G.; Li, T.; Wang, D.Y.; et al. Hydrogen Sulfide Alleviates Lipopolysaccharide-Induced Diaphragm Dysfunction in Rats by Reducing Apoptosis and Inflammation through ROS/MAPK and TLR4/NF-κB Signaling Pathways. Oxid. Med. Cell. Longev. 2018, 2018, 9647809. [Google Scholar] [CrossRef] [Green Version]
- Boshagh, M.A.; Foroutan, P.; Moloudi, M.R.; Fakhari, S.; Malakouti, P.; Nikkhoo, B.; Jalili, A. ELR positive CXCL chemokines are highly expressed in an animal model of ulcerative colitis. J. Inflamm. Res. 2019, 12, 167–174. [Google Scholar] [CrossRef] [Green Version]
- de Souza, M.C.; Vieira, A.J.; Beserra, F.P.; Pellizzon, C.H.; Nóbrega, R.H.; Rozza, A.L. Gastroprotective effect of limonene in rats: Influence on oxidative stress, inflammation and gene expression. Phytomedicine 2019, 53, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, B.; Fan, K.; Wu, J.; Wang, T. CypB-CD147 Signaling Is Involved in Crosstalk between Cartilage and FLS in Collagen-Induced Arthritis. Mediators Inflamm. 2020, 2020, 6473858. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Li, Y.; Xiao, L.; Dai, G.; Lu, P.; Wang, Y.; Rui, Y. AQP1 modulates tendon stem/progenitor cells senescence during tendon aging. Cell Death Dis. 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.; Zhao, L.; Xu, C.; Zhang, L.; Zhao, H. High Molecular Weight Hyaluronan Suppresses Macrophage M1 Polarization and Enhances IL-10 Production in PM2.5-Induced Lung Inflammation. Molecules 2019, 24, 1766. [Google Scholar] [CrossRef] [Green Version]
- Hamid, M.; Liu, D.; Abdulrahim, Y.; Liu, Y.; Qian, G.; Khan, A.; Gan, F.; Huang, K. Amelioration of CCl(4)-induced liver injury in rats by selenizing Astragalus polysaccharides: Role of proinflammatory cytokines, oxidative stress and hepatic stellate cells. Res. Vet. Sci. 2017, 114, 202–211. [Google Scholar] [CrossRef]
- Li, Y.; Xiong, Y.; Zhang, H.; Li, J.; Wang, D.; Chen, W.; Yuan, X.; Su, Q.; Li, W.; Huang, H.; et al. Ginkgo biloba extract EGb761 attenuates brain death-induced renal injury by inhibiting pro-inflammatory cytokines and the SAPK and JAK-STAT signalings. Sci. Rep. 2017, 7, 45192. [Google Scholar] [CrossRef]
- Yun, Y.H.; Kim, H.Y.; Do, B.S.; Kim, H.S. Angiotensin II inhibits chemokine CCL5 expression in vascular smooth muscle cells from spontaneously hypertensive rats. Hypertens. Res. 2011, 34, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Gu, Y.; Liu, Y.; Zhang, M.; Wang, Y.; Hu, L. Ticagrelor attenuates myocardial ischaemia-reperfusion injury possibly through downregulating galectin-3 expression in the infarct area of rats. Br. J. Clin. Pharmacol. 2018, 84, 1180–1186. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, Y.; Yan, S.; Du, T.; Fu, X.; Gong, X.; Zhou, X.; Zhang, T.; Wang, X. Disease Progression-Dependent Expression of CD200R1 and CX3CR1 in Mouse Models of Parkinson’s Disease. Aging Dis. 2020, 11, 254–268. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yang, W.; Zhang, X.; Yang, S.; Peng, G.; Wu, T.; Zhou, Y.; Huang, C.; Reinach, P.S.; Li, W.; et al. MK2 inhibitor reduces alkali burn-induced inflammation in rat cornea. Sci. Rep. 2016, 6, 28145. [Google Scholar] [CrossRef] [Green Version]
- Mohamadi, Y.; Noori Moghahi, S.; Mousavi, M.; Borhani-Haghighi, M.; Abolhassani, F.; Kashani, I.R.; Hassanzadeh, G. Intrathecal transplantation of Wharton’s jelly mesenchymal stem cells suppresses the NLRP1 inflammasome in the rat model of spinal cord injury. J. Chem. Neuroanat. 2019, 97, 1–8. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Reference |
|---|---|---|---|
| IL-10 | CCTCTGGATACAGCTGCGAC | ATGGCCTTGTAGACACCTTTGT | [69] |
| CXCL1 | AGACAGTGGCAGGGATTCAC | AAGCCTCGCGACCATTCTTG | [70] |
| MPO | ATGTCACAGGGGACATGCG | TCTGTGGCCAGACGGTTATG | [71] |
| NE | CTCCGTGGCCAACGATAAGA | AGGTCACTCGTCCTACAGGT | [72] |
| MMP9 | GCAAACCCTGCGTATTTCCATT | GCGATAACCATCCGAGCGAC | [73] |
| MMP3 | CATGAACTTGGCCACTCCCT | TGGGTACCACGAGGACATCA | [73] |
| Arg-1 | ACAAGACAGGGCTACTTTCAGG | ACAAGACAAGGTCAACGCCA | [74] |
| TNF-α | CTGTGCCTCAGCCTCTTCTC | ACTGATGAGAGGGAGCCCAT | [75] |
| CXCL9 | TGCCTAGACCCAGATTCAGC | AGATGCAGAGCGCTTGTTGG | [70] |
| CXCL10 | TTATTGAAAGCGGTGAGCCAAAG | GCTGTCCATCGGTCTCAGCA | [76] |
| CCL5 | CGTGAAGGAGTATTTTTACACCAGC | CTTGAACCCACTTCTTCTCTGGG | [77] |
| IL-1β | CCCTGAACTCAACTGTGAAATAGCA | CCCAAGTCAAGGGCTTGGAA | [76] |
| Lgals3 | CAACTGGCCCTAGTGCTTATC | CAGAGTGATACTGTTTGCGTTG | [78] |
| IL-6 | GCCCACCAGGAACGAAAGTC | GGCTGGAAGTCTCTTGCGGA | [79] |
| CCL3 | TGCCCTTGCTGTTCTTCTCT | AAAGGCTGCTGGTCTCAAAA | [80] |
| CCL2 | ATGCAGTTAATGCCCCACTC | TTCCTTATTGGGGTCAGCAC | [80] |
| GAPDH | AAGTTCAACGGCACAGTCAAG | ACATACTCAGCACCAGCATCA | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.-S.; Liu, M.; Liu, D.-N.; Shang, Y.-F.; Du, G.-H.; Wang, Y.-H. Network Pharmacology Analysis and Experimental Validation of Kaempferol in the Treatment of Ischemic Stroke by Inhibiting Apoptosis and Regulating Neuroinflammation Involving Neutrophils. Int. J. Mol. Sci. 2022, 23, 12694. https://doi.org/10.3390/ijms232012694
Zhang S-S, Liu M, Liu D-N, Shang Y-F, Du G-H, Wang Y-H. Network Pharmacology Analysis and Experimental Validation of Kaempferol in the Treatment of Ischemic Stroke by Inhibiting Apoptosis and Regulating Neuroinflammation Involving Neutrophils. International Journal of Molecular Sciences. 2022; 23(20):12694. https://doi.org/10.3390/ijms232012694
Chicago/Turabian StyleZhang, Shan-Shan, Man Liu, Dong-Ni Liu, Yu-Fu Shang, Guan-Hua Du, and Yue-Hua Wang. 2022. "Network Pharmacology Analysis and Experimental Validation of Kaempferol in the Treatment of Ischemic Stroke by Inhibiting Apoptosis and Regulating Neuroinflammation Involving Neutrophils" International Journal of Molecular Sciences 23, no. 20: 12694. https://doi.org/10.3390/ijms232012694
APA StyleZhang, S. -S., Liu, M., Liu, D. -N., Shang, Y. -F., Du, G. -H., & Wang, Y. -H. (2022). Network Pharmacology Analysis and Experimental Validation of Kaempferol in the Treatment of Ischemic Stroke by Inhibiting Apoptosis and Regulating Neuroinflammation Involving Neutrophils. International Journal of Molecular Sciences, 23(20), 12694. https://doi.org/10.3390/ijms232012694