
Genome-Wide Identification and Expression Analysis of SWEET Family Genes in Sweet Potato and Its Two Diploid Relatives
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of SWEETs in the Sweet Potato and Two Diploid Relatives
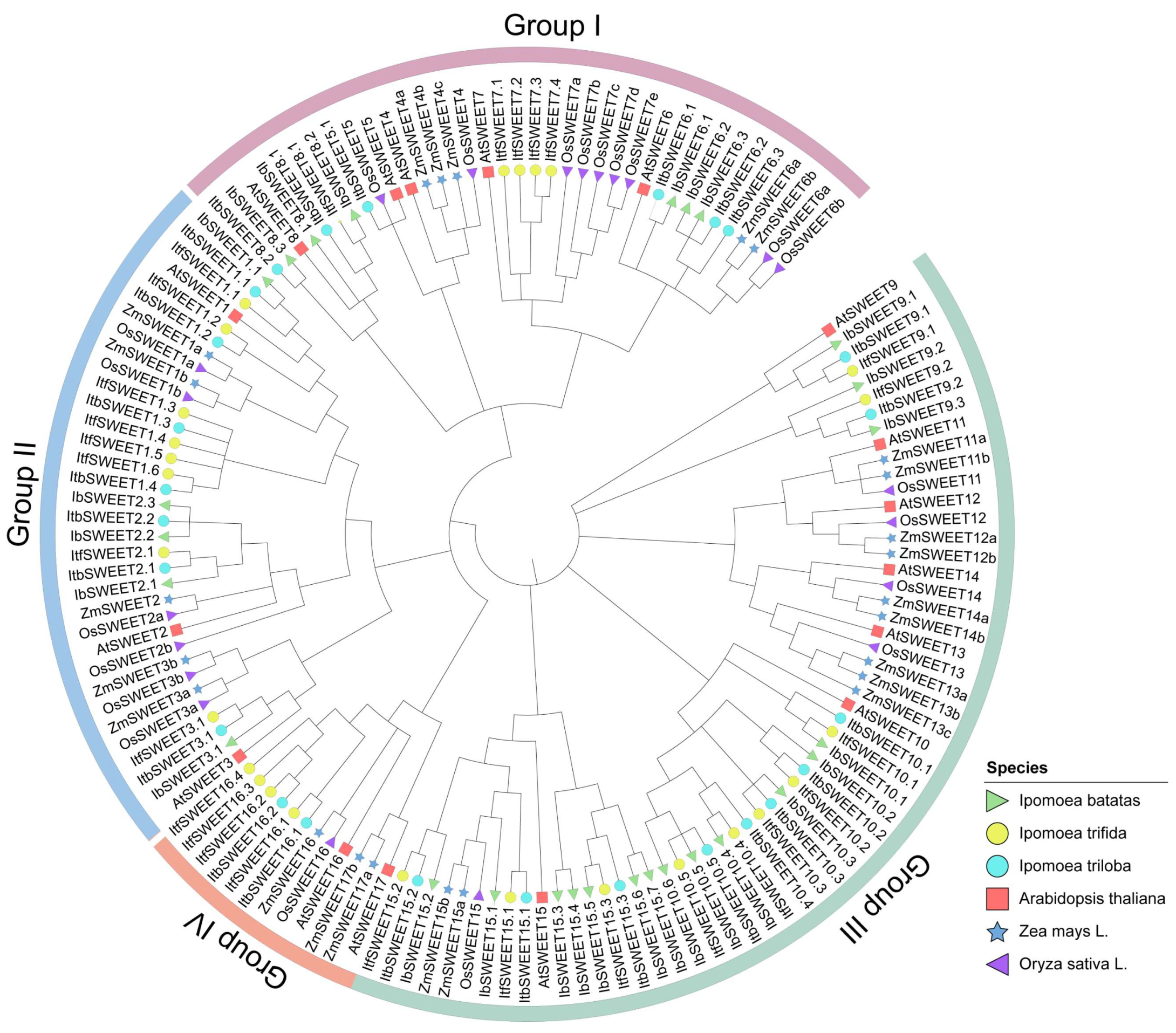
2.2. Phylogenetic Relationship of SWEETs in the Sweet Potato and Its Two Diploid Relatives
2.3. Conserved Motif and Exon–Intron Structure Analysis of SWEETs in the Sweet Potato and Two Diploid Relatives
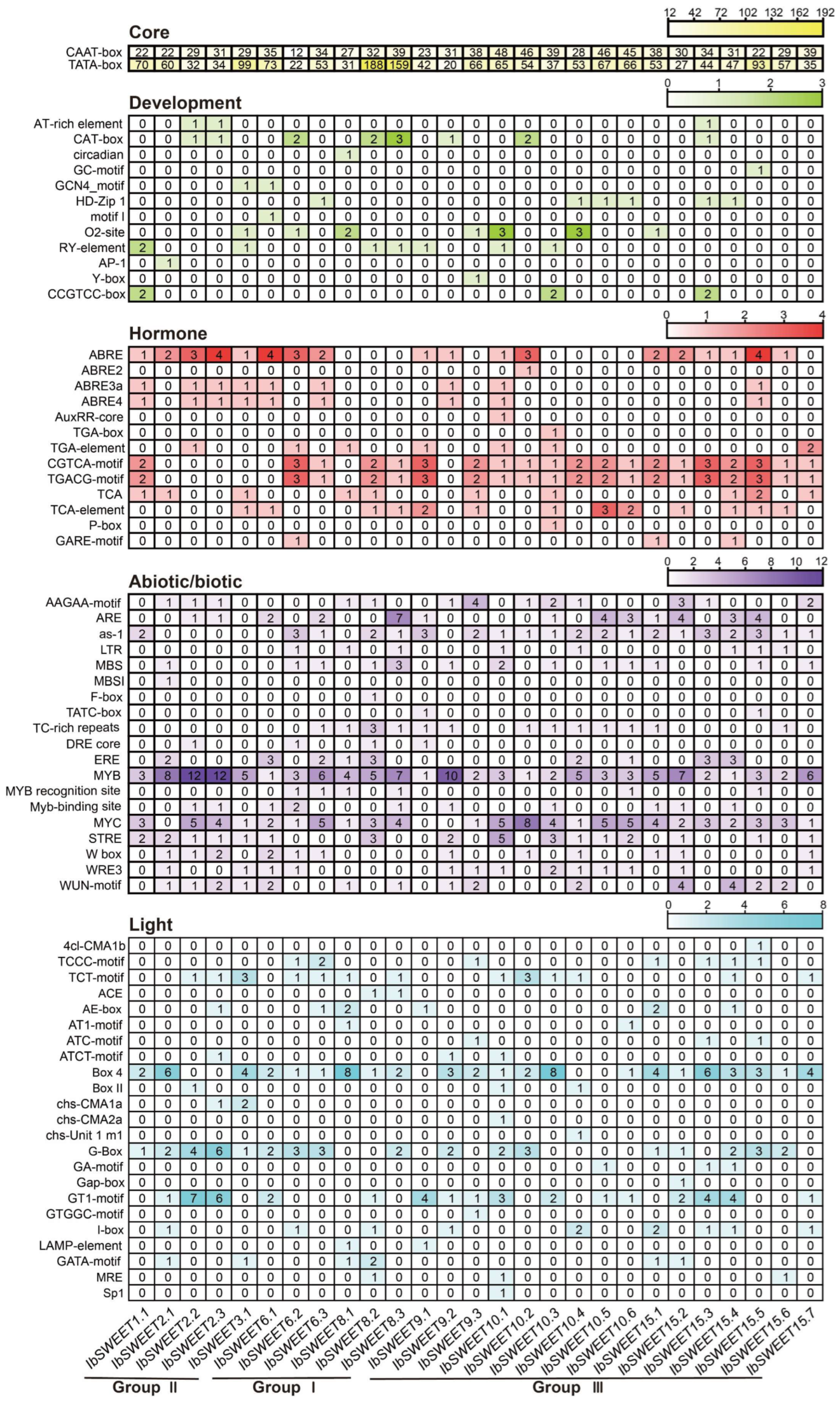
2.4. Cis-Element Analysis in the Promoter of IbSWEETs in Sweet Potato
2.5. Protein Interaction Network of IbSWEETs in the Sweet Potato
2.6. Expression Analysis of SWEETs in the Sweet Potato and Two Diploid Relatives
2.6.1. Expression Analysis in Various Tissues
2.6.2. Expression Analysis in Different Developmental Stages
2.6.3. Expression Analysis in Different Varieties
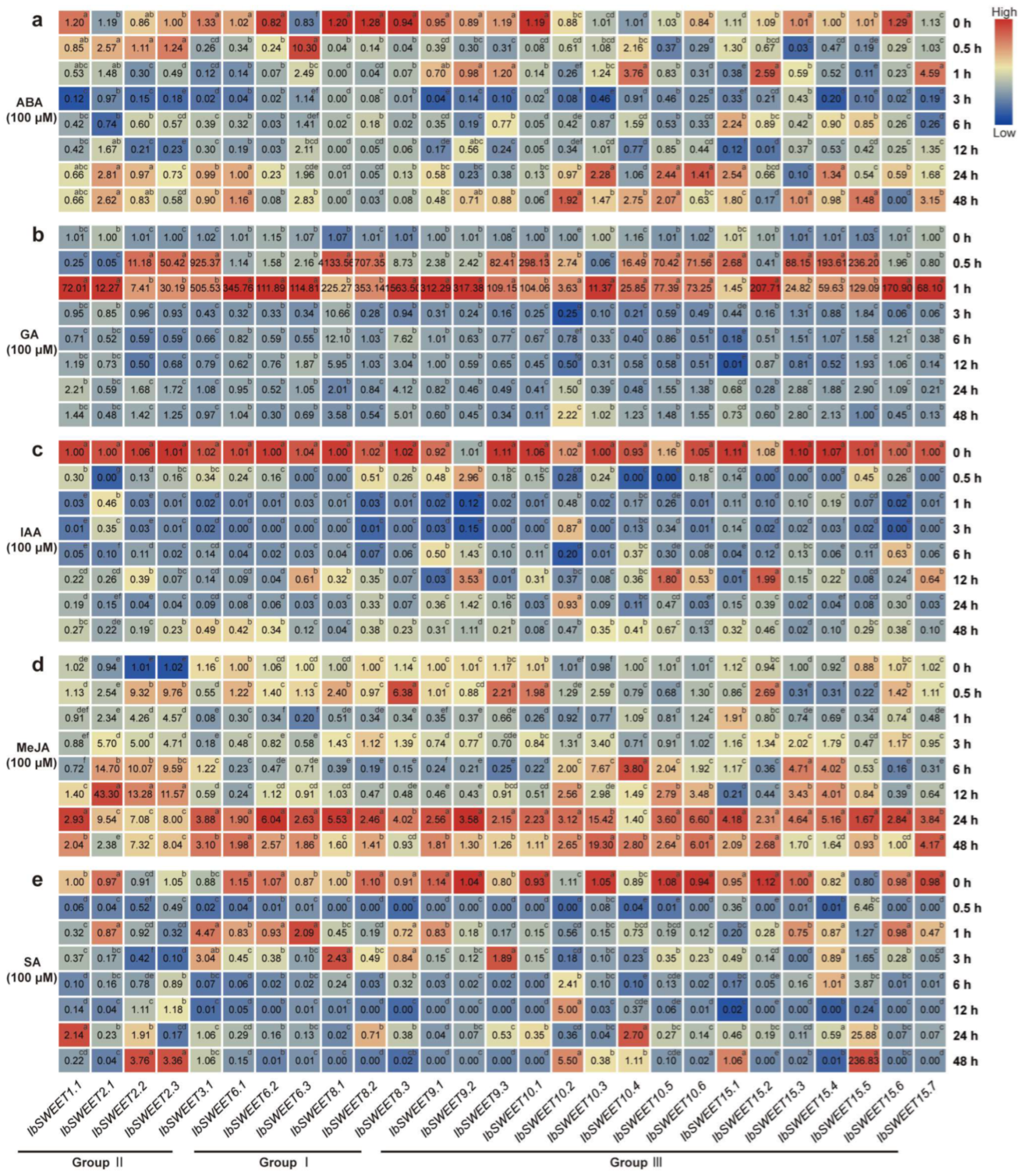
2.6.4. Expression Analysis of Hormone Response
2.6.5. Expression Analysis under Abiotic Stresses
3. Discussion
3.1. Evolution of the SWEET Gene Family in the Sweet Potato and Its Two Diploid Relatives
3.2. Different Functions of SWEETs in Tuberous Root Development in Sweet Potato
3.3. Different Functions of SWEETs in Hormone Crosstalk in the Sweet Potato and Its Two Diploid Relatives
3.4. Different Functions of SWEETs in Abiotic Stress Response in the Sweet Potato and Its Two Diploid Relatives
4. Materials and Methods
4.1. Identification of SWEETs
4.2. Chromosomal Distribution of SWEETs
4.3. Protein Properties Prediction of SWEETs
4.4. Phylogenetic Analysis of SWEETs
4.5. Domain Identification and Conserved Motif Analysis of SWEETs
4.6. Exon–Intron Structures and Promoter Analysis of SWEETs
4.7. Protein Interaction Network of SWEETs
4.8. qRT-PCR Analysis of SWEETs
4.9. Transcriptome Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Yan, C.; Li, Y.; Hirata, K.; Yamamoto, M.; Yan, N.; Hu, Q. Crystal structure of a bacterial homologue of SWEET transporters. Cell Res. 2014, 24, 1486–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.; Frommer, W.B. Structure and function of SemiSWEET and SWEET sugar transporters. Trends Biochem. Sci. 2015, 40, 480–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, Y.H.; Hu, Y.B.; Chen, L.Q.; Sosso, D.; Ducat, D.C.; Hou, B.H.; Frommer, W.B. Functional role of oligomerization for bacterial and plant SWEET sugar transporter family. Proc. Natl. Acad. Sci. USA 2013, 110, E3685–E3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjali, A.; Fatima, U.; Manu, M.S.; Ramasamy, S.; Senthil-Kumar, M. Structure and regulation of SWEET transporters in plants: An update. Plant Physiol. Biochem. 2020, 156, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gamas, P.; de Carvalho-Niebel, F.; Lescure, N.; Cullimore, J.V. Use of a subtractive hybridization approach to identify new Medicago truncatula genes induced during root nodule development. Mol. Plant Microbe Interact. 1996, 9, 233–242. [Google Scholar] [CrossRef]
- Tao, Y.Y.; Cheung, L.S.; Li, S.; Eom, J.S.; Chen, L.Q.; Xu, Y.; Perry, K.; Frommer, W.B.; Feng, L. Structure of a eukaryotic SWEET transporter in a homotrimeric complex. Nature 2015, 527, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Nishizawa, T.; Yamashita, K.; Ishitani, R.; Nureki, O. Structural basis for the facilitative diffusion mechanism by SemiSWEET transporter. Nat. Commun. 2015, 6, 6112. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Tao, Y.; Cheung, L.S.; Fan, C.; Chen, L.-Q.; Xu, S.; Perry, K.; Frommer, W.B.; Feng, L. Structures of bacterial homologues of SWEET transporters in two distinct conformations. Nature 2014, 515, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Q.; Hou, B.H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.Q.; Guo, W.J.; Kim, J.G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhang, C.; Han, X.; Wang, Z.Y.; Ma, L.; Yuan, D.P.; Wu, J.N.; Zhu, X.F.; Liu, J.M.; Li, D.P.; et al. Inhibition of OsSWEET11 function in mesophyll cells improves resistance of rice to sheath blight disease. Mol. Plant Pathol. 2018, 19, 2149–2161. [Google Scholar] [CrossRef]
- Chen, L.Q.; Qu, X.Q.; Hou, B.H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Reinders, A.; Schulze, W.; Kuhn, C.; Barker, L.; Schulz, A.; Ward, J.M.; Frommer, W.B. Protein-protein interactions between sucrose transporters of different affinities colocalized in the same enucleate sieve element. Plant Cell 2002, 14, 1567–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Wang, S. Rice MtN3/Saliva/SWEET family genes and their homologs in cellular organisms. Mol. Plant 2013, 6, 665–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manck-Goetzenberger, J.; Requena, N. Arbuscular mycorrhiza symbiosis induces a major transcriptional reprogramming of the potato SWEET sugar transporter family. Front. Plant Sci. 2016, 7, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, G.; Valliyodan, B.; Deshmukh, R.; Prince, S.; Nicander, B.; Zhao, M.Z.; Sonah, H.; Song, L.; Lin, L.; Chaudhary, J.; et al. Soybean (Glycine max) SWEET gene family: Insights through comparative genomics, transcriptome profiling and whole genome re-sequence analysis. BMC Genom. 2015, 16, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.-J.; Nagy, R.; Chen, H.-Y.; Pfrunder, S.; Yu, Y.-C.; Santelia, D.; Frommer, W.B.; Martinoia, E. SWEET17, a facilitative transporter, mediates fructose transport across the tonoplast of Arabidopsis roots and leaves. Plant Physiol. 2014, 164, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.L.; Holmes-Davis, R.; McCormick, S. Green sperm. Identification of male gamete promoters in arabidopsis. Plant Physiol. 2005, 138, 2124–2133. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Huh, J.H.; Yu, Y.C.; Ho, L.H.; Chen, L.Q.; Tholl, D.; Frommer, W.B.; Guo, W.J. The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection. Plant J. 2015, 83, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, R.; Spinner, L.; Klemens, P.A.W.; Chakraborti, D.; de Marco, F.; Vilaine, F.; Wolff, N.; Lemoine, R.; Porcheron, B.; Gery, C.; et al. Disruption of the sugar transporters AtSWEET11 and AtSWEET12 affects vascular development and freezing tolerance in Arabidopsis. Mol. Plant. 2015, 8, 1687–1690. [Google Scholar] [CrossRef] [Green Version]
- Abelenda, J.A.; Bergonzi, S.; Oortwijn, M.; Sonnewald, S.; Du, M.; Visser, R.G.F.; Sonnewald, U.; Bachem, C.W.B. Source-Sink regulation is mediated by interaction of an ft homolog with a SWEET protein in potato. Curr. Biol. 2019, 29, 1178–1186. [Google Scholar] [CrossRef]
- Ni, J.; Li, J.; Zhu, R.; Zhang, M.; Qi, K.; Zhang, S.; Wu, J. Overexpression of sugar transporter gene PbSWEET4 of pear causes sugar reduce and early senescence in leaves. Gene 2020, 743, 144582. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.F.; Huang, X.Y.; Zhu, J.; Gao, J.F.; Zhang, H.X.; Yang, Z.N. RUPTURED POLLEN GRAIN1, a member of the MtN3/saliva gene family, is crucial for exine pattern formation and cell integrity of microspores in arabidopsis. Plant Physiol. 2008, 147, 852–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Liu, S.; Wang, J.; Yokosho, K.; Zhou, B.; Yu, Y.-C.; Liu, Z.; Frommer, W.B.; Ma, J.F.; Chen, L.-Q.; et al. Simultaneous changes in seed size, oil content and protein content driven by selection of SWEET homologues during soybean domestication. Nat. Sci. Rev. 2020, 7, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Oikawa, T.; Chiba, Y.; Ishimaru, Y.; Shimizu, T.; Sano, N.; Koshiba, T.; Kamiya, Y.; Ueda, M.; Seo, M. AtSWEET13 and AtSWEET14 regulate gibberellin-mediated physiological processes. Nat. Commun. 2016, 7, 13245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morii, M.; Sugihara, A.; Takehara, S.; Kanno, Y.; Kawai, K.; Hobo, T.; Hattori, M.; Yoshimura, H.; Seo, M.; Ueguchi-Tanaka, M. The dual function of OsSWEET3a as a gibberellin and glucose transporter is important for young shoot development in rice. Plant Cell Physiol. 2020, 61, 1935–1945. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, L.; Huang, W.; Yuan, M.; Zhou, F.; Li, X.; Lin, Y. Overexpression of OsSWEET5 in rice causes growth retardation and precocious senescence. PLoS ONE 2014, 9, e94210. [Google Scholar] [CrossRef] [Green Version]
- Bezrutczyk, M.; Hartwig, T.; Horschman, M.; Char, S.N.; Yang, J.; Yang, B.; Frommer, W.B.; Sosso, D. Impaired phloem loading in zmsweet13a,b,c sucrose transporter triple knock-out mutants in Zea mays. New Phytol. 2018, 218, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, Y.; Zhang, H.; Zhang, Q.; Zhai, H.; Liu, Q.; He, S. The Plasma membrane-localized sucrose transporter IbSWEET10 Contributes to the resistance of sweet potato to Fusarium oxysporum. Front. Plant Sci. 2017, 8, 197. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Wu, H.; Huang, W.; Song, J.; Zhou, Y.; Lin, Y. SWEET gene family in Medicago truncatula: Genome-wide identification, expression and substrate specificity analysis. Plants 2019, 8, 338. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Zhang, F.; Song, S.; Yu, X.; Ren, Y.; Zhao, X.; Liu, H.; Liu, G.; Wang, Y.; He, H. CsSWEET2, a hexose transporter from cucumber (Cucumis sativus L.), affects sugar metabolism and improves cold tolerance in Arabidopsis. Int. J. Mol. Sci. 2022, 31, 23. [Google Scholar] [CrossRef]
- Liu, Q.C. Improvement for agronomically important traits by gene engineering in sweetpotato. Breed. Sci. 2017, 67, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Nhanala, S.E.C.; Yencho, G.C. Assessment of the potential of wild Ipomoea spp. for the improvement of drought tolerance in cultivated sweetpotato Ipomoea batatas (L.) Lam. Crop. Sci. 2021, 61, 234–249. [Google Scholar] [CrossRef]
- Nakatani, M.; Komeichi, M. Changes in the endogenous level of zeatin riboside, abscisic-acid and indole acetic-acid during formation and thickening of tuberous roots in sweet-potato. Jpn. J. Crop. Sci. 1991, 60, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Komaki, K.; Katayama, K. Root thickness of diploid Ipomoea trifida (H. B. K.) G. Don and performance of progeny derived from the cross with sweetpotato. Breed. Sci. 1999, 49, 123–129. [Google Scholar] [CrossRef]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.L.; Zheng, J.L.; Sun, Z.; Fan, W.J.; et al. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef]
- Wu, S.; Lau, K.H.; Cao, Q.H.; Hamilton, J.P.; Sun, H.H.; Zhou, C.X.; Eserman, L.; Gemenet, D.C.; Olukolu, B.A.; Wang, H.Y.; et al. Genome sequences of two diploid wild relatives of cultivated sweetpotato reveal targets for genetic improvement. Nat. Commun. 2018, 9, 4580. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; He, J.; Velanis, C.N.; Zhu, Y.; He, Y.; Tang, K.; Zhu, M.; Graser, L.; de Leau, E.; Wang, X.; et al. A domesticated Harbinger transposase forms a complex with HDA6 and promotes histone H3 deacetylation at genes but not TEs in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 1462–1474. [Google Scholar] [CrossRef]
- Paxson-Sowders, D.M.; Dodrill, C.H.; Owen, H.A.; Makaroff, C.A. DEX1, a novel plant protein, is required for exine pattern formation during pollen development in Arabidopsis. Plant Physiol. 2001, 127, 1739–1749. [Google Scholar] [CrossRef]
- Toda, Y.; Kudo, T.; Kinoshita, T.; Nakamichi, N. Evolutionary insight into the clock-associated PRR5 transcriptional network of flowering plants. Sci. Rep. 2019, 9, 2983. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Li, X.; Zeng, B.; Zhong, M.; Yang, J.; Yang, P.; Li, X.; He, C.; Lin, J.; Liu, X.; et al. FKF1 F-box protein promotes flowering in part by negatively regulating DELLA protein stability under long-day photoperiod inArabidopsis. J. Integr. Plant Biol. 2020, 62, 1717–1740. [Google Scholar] [CrossRef]
- Choi, S.; Prokchorchik, M.; Lee, H.; Gupta, R.; Lee, Y.; Chung, E.-H.; Cho, B.; Kim, M.-S.; Kim, S.T.; Sohn, K.H. Direct acetylation of a conserved threonine of RIN4 by the bacterial effector HopZ5 or AvrBsT activates RPM1-dependent immunity in Arabidopsis. Mol. Plant 2021, 14, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Redditt, T.J.; Chung, E.-H.; Zand Karimi, H.; Rodibaugh, N.; Zhang, Y.; Trinidad, J.C.; Kim, J.H.; Zhou, Q.; Shen, M.; Dangl, J.L.; et al. AvrRpm1 Functions as an ADP-Ribosyl Transferase to Modify NOI-domain Containing Proteins, Including Arabidopsis and Soybean RPM1-interacting Protein 4. Plant Cell 2019, 2664–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.-C.; Lin, W.-C.; Cheng, W.-H. Salt hypersensitive mutant 9, a nucleolar APUM23 protein, is essential for salt sensitivity in association with the ABA signaling pathway in Arabidopsis. BMC Plant Biol. 2018, 18, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatov, E.; Ludewig, U.; Zentgraf, U. Disparate dynamics of gene body and cis-regulatory element evolution illustrated for the senescence-associated cysteine protease gene SAG12 of plants. Plants 2021, 10, 1380. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhou, Y.; Zhai, H.; He, S.; Zhao, N.; Liu, Q. Transcriptome profiling reveals insights into the molecular mechanism of drought tolerance in sweetpotato. J. Integr. Agri. 2019, 18, 9–23. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Q.; Zhai, H.; Li, Y.; Wang, X.; Liu, Q.; He, S. Transcript profile analysis reveals important roles of jasmonic acid signalling pathway in the response of sweet potato to salt stress. Sci. Rep. 2017, 7, 40819. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, L.; Zhu, P.; Cao, Q.; Sun, J.; Li, Z.; Xu, T. Genome-wide identification, characterisation and functional evaluation of WRKY genes in the sweet potato wild ancestor Ipomoea trifida (HBK) G. Don. under abiotic stresses. BMC Genet. 2019, 20, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Sun, J.; Yang, Z.; Zhao, C.; Zhu, M.; Ma, D.; Dong, T.; Zhou, Z.; Liu, M.; Yang, D.; et al. Genome-wide identification and expression analysis of glycine-rich RNA-binding protein family in sweet potato wild relative Ipomoea trifida. Gene 2019, 686, 177–186. [Google Scholar] [CrossRef]
- Wan, R.; Liu, J.; Yang, Z.; Zhu, P.; Cao, Q.; Xu, T. Genome-wide identification, characterisation and expression profile analysis of DEAD-box family genes in sweet potato wild ancestor Ipomoea trifida under abiotic stresses. Genes Genom. 2020, 42, 325–335. [Google Scholar] [CrossRef]
- Mukherjee, D.; Saha, D.; Acharya, D.; Mukherjee, A.; Chakraborty, S.; Ghosh, T.C. The role of introns in the conservation of the metabolic genes of Arabidopsis thaliana. Genomics 2018, 110, 310–317. [Google Scholar] [CrossRef]
- Morello, L.; Giani, S.; Troina, F.; Breviario, D. Testing the IMEter on rice introns and other aspects of intron-mediated enhancement of gene expression. J. Exp. Bot. 2011, 62, 533–544. [Google Scholar] [CrossRef]
- Ma, J.; Deng, S.; Jia, Z.; Sang, Z.; Zhu, Z.; Zhou, C.; Ma, L.; Chen, F. Conservation and divergence of ancestral AGAMOUS/SEEDSTICK subfamily genes from the basal angiosperm Magnolia wufengensis. Tree Physiol. 2020, 40, 90–107. [Google Scholar] [CrossRef]
- Ma, R.; Song, W.; Wang, F.; Cao, A.; Xie, S.; Chen, X.; Jin, X.; Li, H. A Cotton (Gossypium hirsutum) Myo-Inositol-1-Phosphate Synthase (GhMIPS1D) gene promotes root cell elongation in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 1224. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.; Wei, Y.; Cheng, Y.; Pan, L.; Ye, Q.; Wang, R.; Ruan, M.; Zhou, G.; Yao, Z.; Li, Z.; et al. The Tryptophan decarboxylase in Solanum lycopersicum. Molecules 2018, 23, 5. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zhang, D.; Miao, Q.; Yang, J.; Xuan, Y.; Hu, Y. Essential role of sugar transporter OsSWEET11 during the early stage of rice grain filling. Plant Cell Physiol. 2017, 58, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Rukundo, P.; Shimelis, H.; Laing, M.; Gahakwa, D. Storage root formation, dry matter synthesis, accumulation and genetics in sweet potato. Aust. J. Crop. Sci. 2013, 7, 2054–2061. [Google Scholar]
- Bihmidine, S.; Julius, B.T.; Dweikat, I.; Braun, D.M. Tonoplast Sugar Transporters (SbTSTs) putatively control sucrose accumulation in sweet sorghum stems. Plant Signal. Behav. 2016, 11, e1117721. [Google Scholar] [CrossRef] [Green Version]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Perez-Perez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin-Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [Green Version]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Tang, Z.; Zhang, Y.; Niu, L.; Yang, F.; Zhang, D.; Hu, Y. Rice SUT and SWEET transporters. Int. J. Mol. Sci. 2021, 22, 20. [Google Scholar] [CrossRef]
- Mizuno, H.; Kasuga, S.; Kawahigashi, H. The sorghum SWEET gene family: Stem sucrose accumulation as revealed through transcriptome profiling. Biotechnol. Biofuels 2016, 9, 127. [Google Scholar] [CrossRef]
- Jameson, P.E.; Dhandapani, P.; Novak, O.; Song, J. Cytokinins and expression of SWEET, SUT, CWINV and AAP genes increase as pea seeds germinate. Int. J. Mol. Sci. 2016, 17, 12. [Google Scholar] [CrossRef] [Green Version]
- Conde, A.; Soares, F.; Breia, R.; Geros, H. Postharvest dehydration induces variable changes in the primary metabolism of grape berries. Food Res. Int. 2018, 105, 261–270. [Google Scholar] [CrossRef]
- Durand, M.; Porcheron, B.; Hennion, N.; Maurousset, L.; Lemoine, R.; Pourtau, N. Water deficit enhances C export to the roots in Arabidopsis thaliana plants with contribution of sucrose transporters in both shoot and roots. Plant Physiol. 2016, 170, 1460–1479. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, C.; Xu, H.; Lin, S.; Deng, W.; Zhou, J.; Zhang, Y.; Shi, Y.; Peng, D.; Xue, Y. GPS 5.0: An update on the prediction of kinase-specific phosphorylation sites in proteins. Genom. Proteom. Bioinform. 2020, 18, 72–80. [Google Scholar] [CrossRef]
- Kopp, J.; Schwede, T. The SWISS-MODEL Repository of annotated three-dimensional protein structure homology models. Nucleic Acids Res. 2004, 32, D230–D234. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. Methods Mol. Biol. 2011, 696, 291–303. [Google Scholar] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | PI | MW/kDa | Genomic Length/bp | CDS Length/bp | Phosphorylation Site | Protein Size/aa | Aliphatic Index | GRAVY | TMHs | Subcellular Locations | Arabidopsis Homologous | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ser | Thr | Tyr | ||||||||||||
| g42355 | IbSWEET1.1 | 9.55 | 27.63 | 1949 | 1158 | 17 | 12 | 6 | 254 | 120.47 | 0.819 | 7 | Cell membrane Golgi apparatus | SWEET1 |
| g45970 | IbSWEET2.1 | 9.18 | 30.50 | 2865 | 1303 | 30 | 11 | 12 | 273 | 114.58 | 0.788 | 7 | Cell membrane | SWEET2 |
| g37512 | IbSWEET2.2 | 8.97 | 26.17 | 2620 | 1086 | 23 | 12 | 14 | 235 | 125.19 | 1.003 | 7 | Cell membrane | SWEET2 |
| g37574 | IbSWEET2.3 | 9.44 | 19.99 | 4204 | 1557 | 17 | 11 | 9 | 179 | 125.70 | 1.070 | 5 | Cell membrane | SWEET2 |
| g20639 | IbSWEET3.1 | 8.83 | 24.44 | 1825 | 829 | 20 | 12 | 10 | 217 | 98.25 | 0.281 | 5 | Cell membrane | SWEET3 |
| g39263 | IbSWEET6.1 | 8.46 | 30.93 | 2934 | 1046 | 19 | 17 | 12 | 278 | 126.19 | 0.871 | 7 | Cell membrane | SWEET6 |
| g39260 | IbSWEET6.2 | 9.15 | 25.53 | 2101 | 868 | 19 | 16 | 11 | 233 | 105.41 | 0.481 | 5 | Cell membrane | SWEET6 |
| g39262 | IbSWEET6.3 | 9.30 | 25.79 | 2900 | 983 | 22 | 15 | 11 | 237 | 112.32 | 0.523 | 6 | Cell membrane | SWEET6 |
| g5800 | IbSWEET8.1 | 9.83 | 22.47 | 1052 | 966 | 15 | 10 | 7 | 206 | 117.86 | 0.639 | 6 | Cell membrane | SWEET8 |
| g346 | IbSWEET8.2 | 9.47 | 25.72 | 1977 | 1065 | 17 | 11 | 10 | 235 | 120.68 | 0.681 | 7 | Cell membrane | SWEET8 |
| g51687 | IbSWEET8.3 | 9.98 | 26.48 | 2536 | 1055 | 16 | 14 | 7 | 239 | 108.20 | 0.592 | 6 | Cell membrane | SWEET8 |
| g41769 | IbSWEET9.1 | 9.16 | 27.26 | 1912 | 823 | 12 | 7 | 14 | 241 | 119.71 | 0.747 | 7 | Cell membrane | SWEET9 |
| g49942 | IbSWEET9.2 | 9.48 | 30.39 | 5035 | 1049 | 15 | 14 | 17 | 267 | 114.68 | 0.696 | 6 | Cell membrane | SWEET9 |
| g33162 | IbSWEET9.3 | 8.72 | 30.49 | 2028 | 1395 | 16 | 22 | 13 | 275 | 122.15 | 0.691 | 6 | Cell membrane | SWEET9 |
| g6315 | IbSWEET10.1 | 8.83 | 31.13 | 2310 | 1122 | 16 | 16 | 14 | 278 | 117.73 | 0.700 | 7 | Cell membrane | SWEET10 |
| g33248 | IbSWEET10.2 | 9.34 | 34.07 | 3208 | 1235 | 17 | 18 | 11 | 305 | 114.72 | 0.549 | 7 | Cell membrane | SWEET10 |
| g55355 | IbSWEET10.3 | 9.20 | 34.65 | 1851 | 1231 | 18 | 11 | 13 | 314 | 122.26 | 0.689 | 7 | Cell membrane | SWEET10 |
| g38390 | IbSWEET10.4 | 9.19 | 34.25 | 2664 | 1264 | 21 | 17 | 11 | 304 | 117.57 | 0.607 | 7 | Cell membrane | SWEET10 |
| g14486 | IbSWEET10.5 | 9.48 | 30.78 | 3130 | 1123 | 18 | 15 | 9 | 272 | 106.76 | 0.521 | 5 | Cell membrane | SWEET10 |
| g14649 | IbSWEET10.6 | 9.39 | 32.65 | 3831 | 1188 | 17 | 17 | 11 | 288 | 116.39 | 0.678 | 7 | Cell membrane | SWEET10 |
| g4174 | IbSWEET15.1 | 7.61 | 35.41 | 2008 | 1238 | 19 | 19 | 11 | 321 | 114.70 | 0.568 | 7 | Cell membrane | SWEET15 |
| g39828 | IbSWEET15.2 | 8.19 | 33.64 | 2933 | 1057 | 19 | 16 | 12 | 302 | 115.79 | 0.541 | 6 | Cell membrane | SWEET15 |
| g13599 | IbSWEET15.3 | 9.46 | 24.64 | 1780 | 896 | 16 | 10 | 8 | 221 | 127.87 | 0.802 | 6 | Cell membrane | SWEET15 |
| g13600 | IbSWEET15.4 | 9.30 | 24.86 | 1917 | 920 | 19 | 12 | 8 | 222 | 124.19 | 0.821 | 6 | Cell membrane | SWEET15 |
| g13601 | IbSWEET15.5 | 7.74 | 32.80 | 1767 | 1103 | 24 | 9 | 11 | 292 | 120.17 | 0.664 | 7 | Cell membrane | SWEET15 |
| g61464 | IbSWEET15.6 | 5.81 | 31.87 | 1594 | 1026 | 29 | 13 | 9 | 278 | 119.10 | 0.729 | 4 | Cell membrane Chloroplast | SWEET15 |
| g61461 | IbSWEET15.7 | 9.47 | 17.64 | 5747 | 988 | 14 | 9 | 7 | 153 | 127.97 | 0.907 | 6 | Cell membrane Chloroplast | SWEET15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Z.; Yan, P.; He, S.; Jia, L.; Wang, Y.; Liu, Q.; Zhai, H.; Zhao, N.; Gao, S.; Zhang, H. Genome-Wide Identification and Expression Analysis of SWEET Family Genes in Sweet Potato and Its Two Diploid Relatives. Int. J. Mol. Sci. 2022, 23, 15848. https://doi.org/10.3390/ijms232415848
Dai Z, Yan P, He S, Jia L, Wang Y, Liu Q, Zhai H, Zhao N, Gao S, Zhang H. Genome-Wide Identification and Expression Analysis of SWEET Family Genes in Sweet Potato and Its Two Diploid Relatives. International Journal of Molecular Sciences. 2022; 23(24):15848. https://doi.org/10.3390/ijms232415848
Chicago/Turabian StyleDai, Zhuoru, Pengyu Yan, Shaozhen He, Licong Jia, Yannan Wang, Qingchang Liu, Hong Zhai, Ning Zhao, Shaopei Gao, and Huan Zhang. 2022. "Genome-Wide Identification and Expression Analysis of SWEET Family Genes in Sweet Potato and Its Two Diploid Relatives" International Journal of Molecular Sciences 23, no. 24: 15848. https://doi.org/10.3390/ijms232415848
APA StyleDai, Z., Yan, P., He, S., Jia, L., Wang, Y., Liu, Q., Zhai, H., Zhao, N., Gao, S., & Zhang, H. (2022). Genome-Wide Identification and Expression Analysis of SWEET Family Genes in Sweet Potato and Its Two Diploid Relatives. International Journal of Molecular Sciences, 23(24), 15848. https://doi.org/10.3390/ijms232415848