1. Introduction
There are six enzymatic steps involved in the de novo pyrimidine biosynthesis pathway, which is nearly universal to all organisms [
1,
2,
3,
4]. The end product of the pathway, uridine monophosphate, is the starting point for the delivery of deoxynucleoside triphosphate (dNTP) precursors for the synthesis of DNA, nucleoside triphosphate (NTP) precursors for RNA, as well as glycoconjugates, and many more metabolically important molecules. The fourth enzymatic reaction in this pathway, the oxidation of dihydroorotate to orotate, is catalyzed by the flavoenzyme dihydroorotate dehydrogenase (DHODH) [
5,
6,
7].
DHODHs can be divided into two major families or classes, I and II, based on the sequence similarities, rather than convergent evolution of different ancestral proteins, with a further subdivision of Class I into sub-classes IA and IB [
8,
9]. This division correlates with the quaternary structure, subcellular location of the enzymes, as well as their preferences for electron acceptors. Members of Class I are cytosolic, with Class IA enzymes being homodimers and Class IB enzymes being heterotetrameric proteins composed of two different proteins. DHODH from
Escherichia coli is regarded as the prototype of Class II DHODHs [
8,
10]. In contrast to Class I enzymes, Class II DHODHs are monomeric membrane-bound proteins that use ubiquinones as electron receptors.
Human DHODH (
HsDHODH) is located on the outside of the inner mitochondrial membrane (IMM) and uses ubiquinone Q
10 as an electron acceptor, which functionally connects its activity to the respiratory chain [
11,
12,
13]. Literature suggests that
HsDHODH is a stand-alone enzyme not associated with any respiratory supercomplexes [
14].
HsDHODH is probably the most studied member of the Class II enzymes because it is a target for anti-inflammatory drugs, such as leflunomide (ARAVA
®), approved for rheumatoid arthritis and its active metabolite, and teriflunomide (AUBAGIO
®), approved for multiple sclerosis, both proposed to interact with the same region of the enzyme as ubiquinone [
15]. It was also recently validated as a target for the treatment of acute myeloid leukemia (AML) [
16], as its inhibition overcomes the myeloid cell differentiation blockade. Therefore, novel and old
HsDHODH inhibitors are of interest for the treatment of hematological malignancies and innovative treatment options [
17,
18], and four compounds have entered clinical trials [
19,
20]. Furthermore, mutations in
HsDHODH have been identified as the cause of Miller syndrome [
21,
22], a rare autosomal recessive disorder (OMIM %263750) resulting in numerous abnormalities of the head, face, and limbs.
HsDHODH was also recently identified as the mitochondrial gatekeeper of cell death by ferroptosis because its deletion promotes ferroptosis [
23].
HsDHODH inhibitors also exhibit antiviral activity against a range of different viruses, with their effect being attributed to the depletion of the nucleosides that are necessary for the replication of the viral genome.
HsDHODH inhibition has therefore been proposed as a potential alternative intervention strategy in severe viral infections, e.g., caused by the Ebola virus [
24,
25] or respiratory RNA viruses, including the coronaviruses [
26,
27]. During the ongoing COVID-19 pandemic, several
HsDHODH inhibitors have been shown to inhibit the replication of SARS-CoV-2 (and other RNA viruses) in cell cultures [
28,
29,
30]. It is therefore no wonder that approved anti-inflammatory
HsDHODH inhibitors, as well as compounds in clinical trials for AML, are now also tested for their effects in COVID-19 patients [
28,
31,
32]. The bacterial
E. coli DHODH (
EcDHODH) is peripherally associated with the cytosolic membrane [
33] and uses ubiquinones as electron acceptors during aerobic growth, but must also be able to use alternative electron receptors (menaquinone, demethylmenaquinone) during anaerobic growth [
34]. The structure and proposed orientations of
HsDHODH in the IMM and
EcDHODH at the cytosolic bacterial membrane are illustrated in
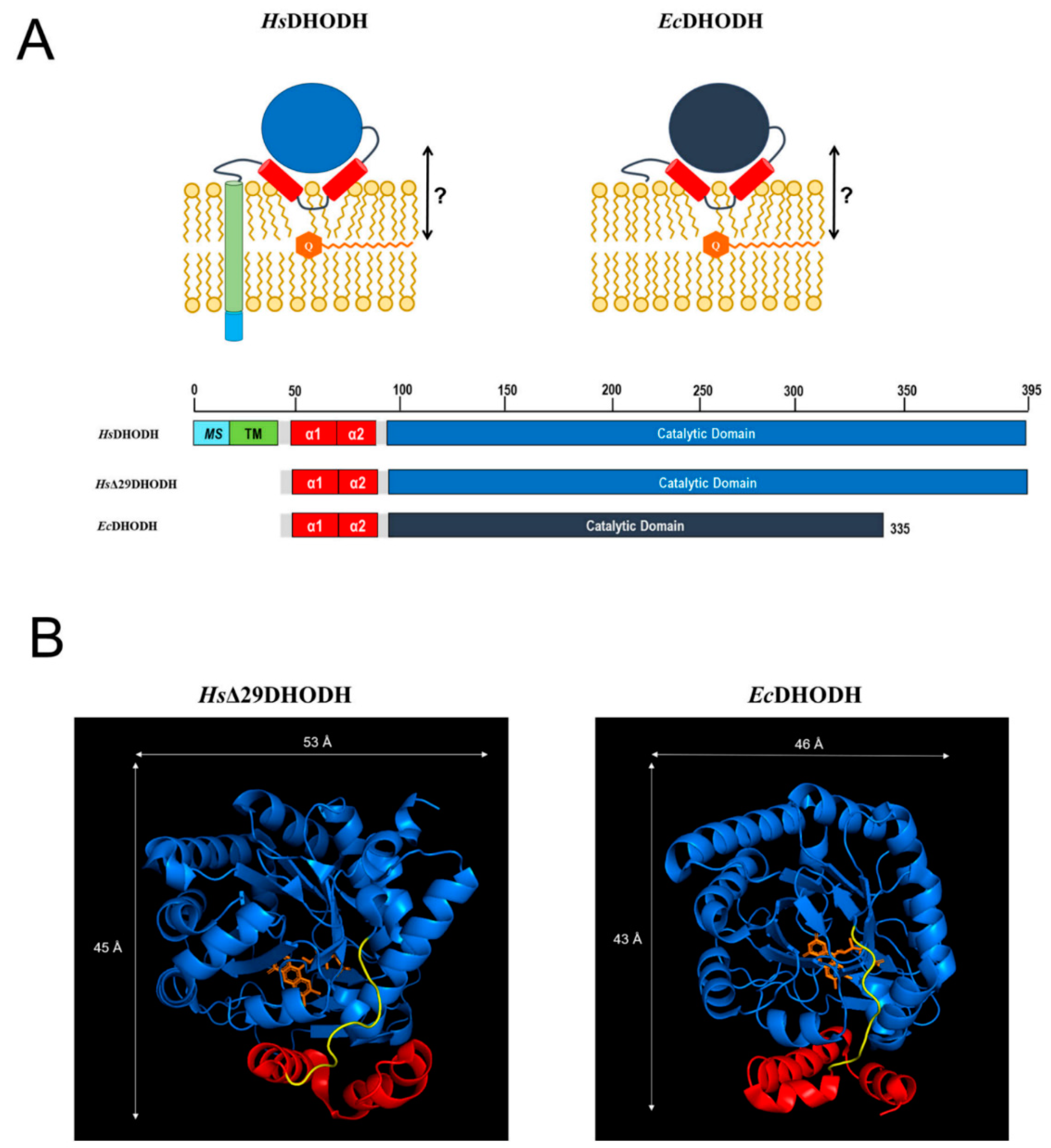
Figure 1A, which also displays the proposed location of ubiquinone within the membrane. The N-terminus of
HsDHODH contains a bipartite signal consisting of a mitochondrial signal (MS) and transmembrane domain (TM) that determines its import and correct insertion into the IMM [
13]. The N-terminal segments of Class II DHODHs are connected to the catalytic domain via a microdomain containing two alpha-helices (α1, α2). In the
E. coli enzyme, this microdomain is an N-terminal patch of two alpha-helices followed by a short 3
10-helix [
9]. This microdomain is proposed to determine the interaction of DHODH with the membrane, and the binding of the electron acceptor ubiquinone. We will henceforth refer to this protein segment simply as the α1-α2 microdomain. In
HsDHODH and in the DHODH from
Rattus norvegicus, this part of the enzyme is an important target for the binding of clinically used DHODH inhibitors, such as the active metabolite of ARAVA
® or atovaquone, and the former drug candidate brequinar [
7,
35,
36]. In
EcDHODH, the structural elements corresponding to the MS and TM are missing, as illustrated in
Figure 1A.
Many crystal structures of the soluble catalytic domain of
HsDHODH in complex with inhibitors are currently available [
7], but there are hardly any experimental structural data regarding the common feature of Class II DHODHs, the α1-α2 microdomain, or data concerning the enzyme in a membrane-bound state. To date, spectroscopic measurements indicate that the α1-α2 microdomain assumes a different conformation in detergent micelles and phospholipid vesicles [
37], indicating that the nature of the membrane environment may play a role in the conformation adopted. Another interesting question is how the enzyme interacts with ubiquinone, which has been shown to be located in the hydrophobic center of lipid membranes [
38], but it also needs to access the DHODH membrane-targeting α1-α2 microdomain as many DHODH inhibitors do [
15]. Recently, it also has been proposed that inhibitor binding to DHODH is influenced by interactions with lipids in the IMM [
39]. This is supported by previous findings showing differences in the inhibition of DHODH by brequinar, depending on whether the TM is present or not [
35].
We therefore designed this study to understand the interaction of Class II DHODHs with membrane lipids and with the electron acceptor, ubiquinone. For our study, we chose the following two prominent members of Class II DHODHs:
EcDHODH and
HsDHODH. We performed a series of neutron reflectometry (NR) experiments in order to investigate the binding of both enzymes to a range of supported lipid bilayers with and without the co-substrate ubiquinone in order to determine what features of the membrane are important for the interaction with the enzymes and what role ubiquinone plays. These bilayers ranged from simple model bilayers that were prepared from synthetic lipids to complex lipid mixtures what were extracted from cell cultures. In order to focus on the proposed common feature of Class II DHODHs interaction with the membrane in this study, the α1-α2 microdomain, we compared the
EcDHODH with an N-terminal truncated version of
HsDHODH (
HsΔ29DHODH), which lacks the MS and TM domains (
Figure 1A).
HsΔ29DHODH is comparable in enzymatic activity to the full-length
HsDHODH that is found naturally in mitochondria and the is most used variant of
HsDHODH studied throughout literature, also in interaction with lipids. Several reports of this, as well as comparisons of Class II DHODH with and without their N-terminal TM in vivo and
in vitro studies have been published [
13,
35,
40,
41,
42,
43,
44].
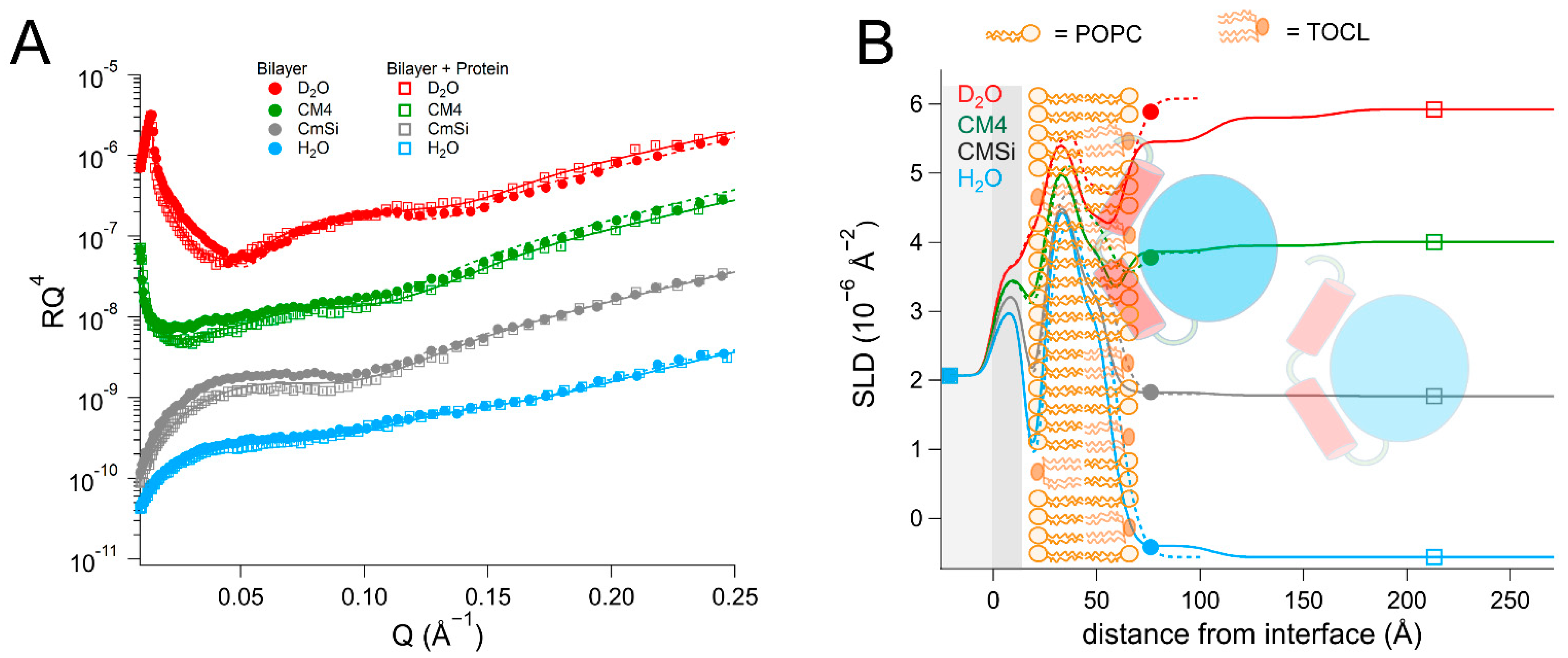
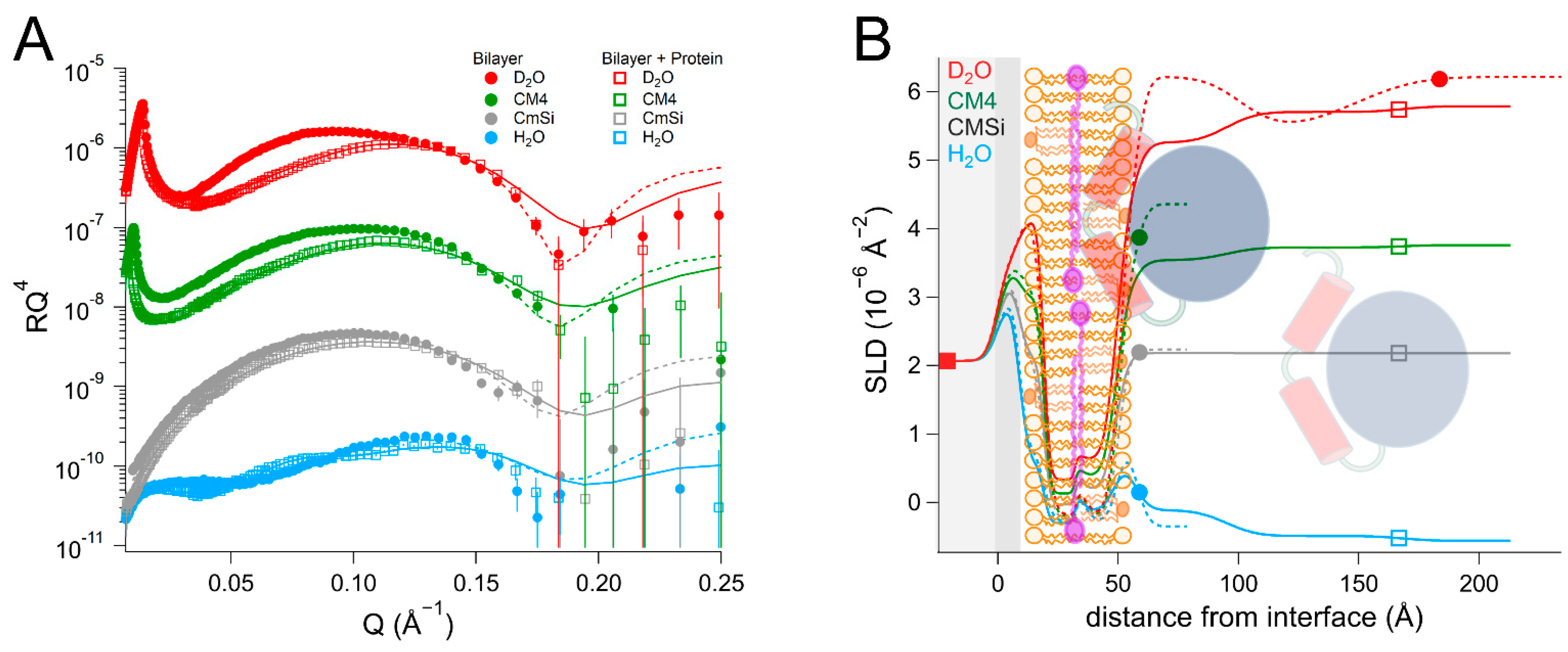
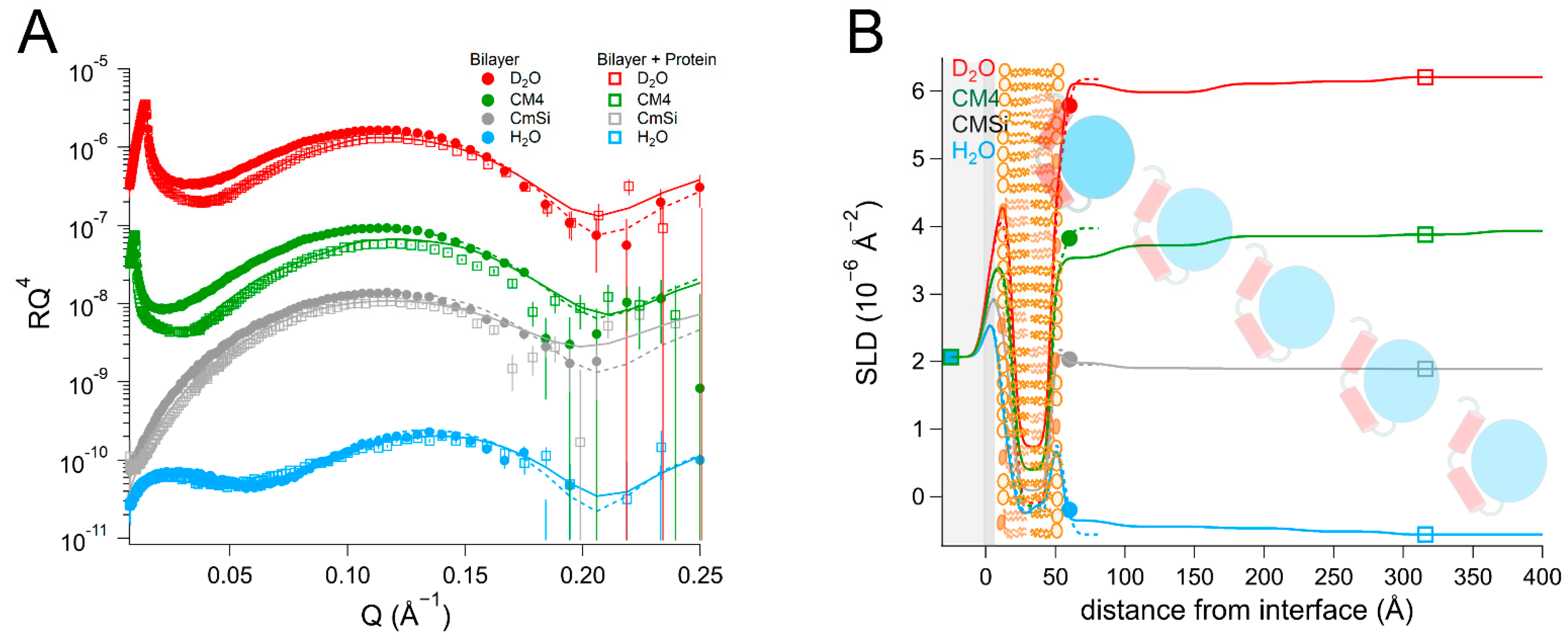
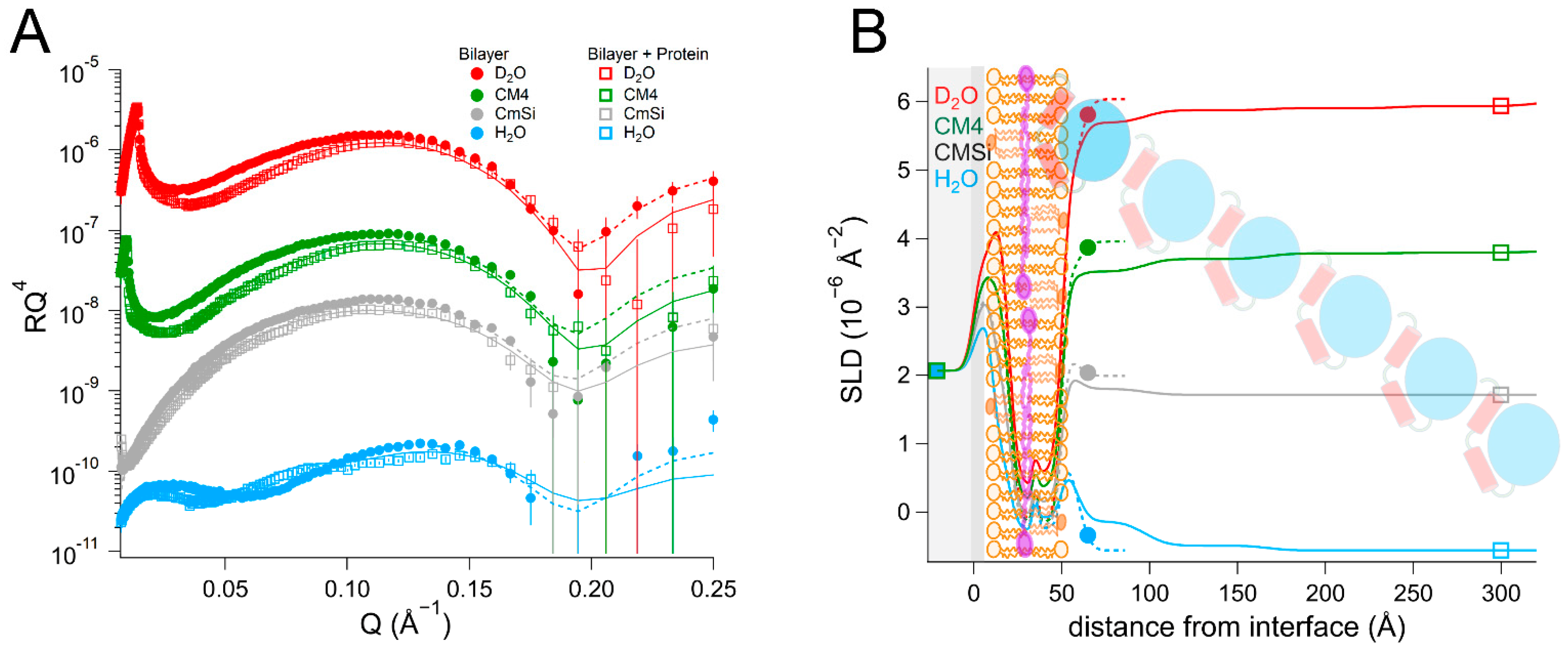
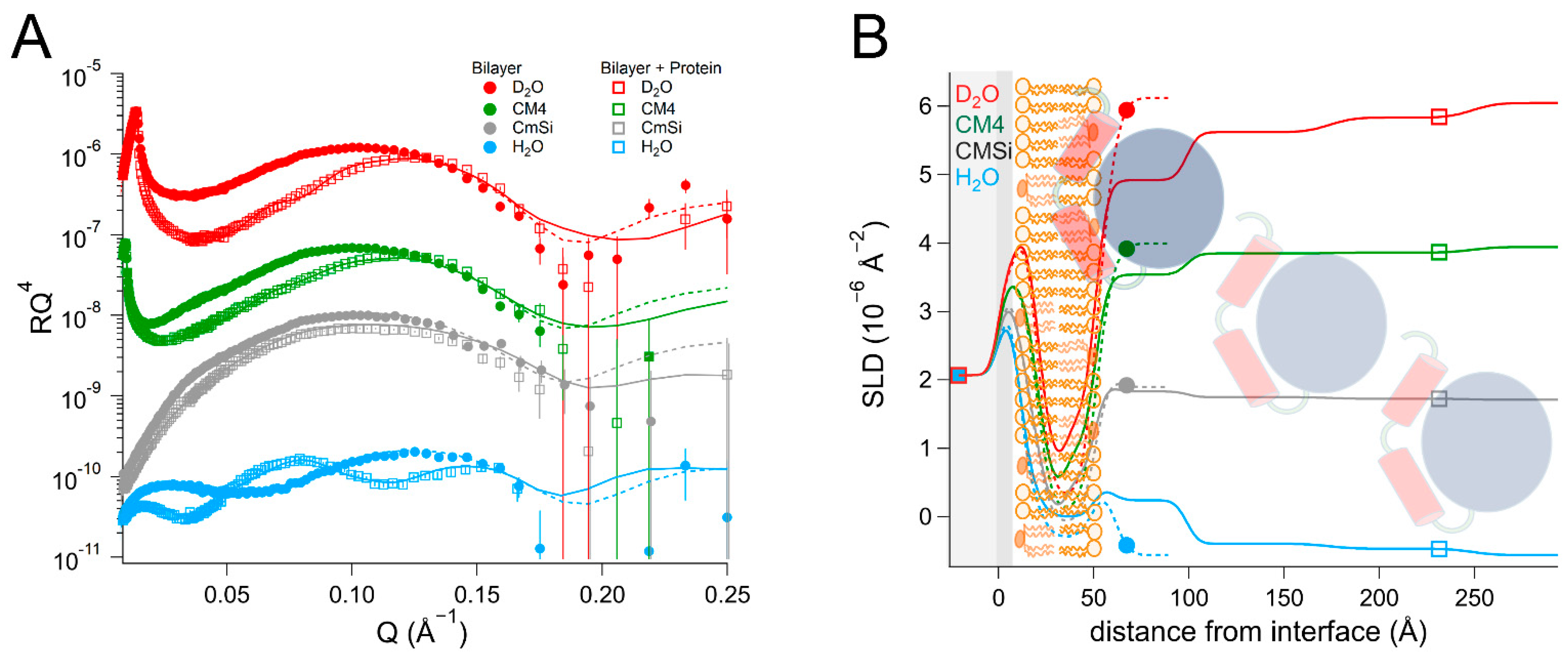
NR is a particularly well-suited technique for addressing these questions as it can determine the one-dimensional depth profile of single supported lipid membranes to 2–3 Å resolution, under solution conditions [
45,
46,
47,
48,
49], and provide a low-resolution distribution profile of the embedded proteins using selective deuterium labeling of either the protein or the lipids. Recently developed methods allow the selective reconstitution of physiologically relevant membranes with a well-defined lipid composition [
48]. NR is also well-suited for studying mechanisms and protein activity in situ [
50,
51,
52], due to the flow-cell geometry of the samples. Additionally, the development of the synthesis of unsaturated deuterated lipids [
53], and the purification of deuterated lipids from cell cultures [
54], means that protein deuteration is not always required in order to achieve the required neutron contrast.
In mammalian cells, the inner mitochondrial membrane hosting
HsDHODH consists mainly of phosphatidylcholine (38–45%), phosphatidylethanolamine (32–39%), cardiolipin (14–23%), and phosphatidylinositol (2–7%) lipids. The outer mitochondrial membrane also consists mostly of phosphatidylcholine (44–59%) and phosphatidylethanolamine (20–35%) but contains a significantly lower amount of cardiolipin (1–10%) and a higher proportion of phosphatidylinositol (~13%) [
55,
56]. There are also variations in lipid composition depending on the tissue and species. In rats, for example, the cardiolipin content is significantly higher in the liver and kidneys (9–20%) than in the brain, lungs, or heart, where it ranges from 2 to 8% [
55]. The
E. coli cytosolic membrane that
EcDHODH [
57] is associated with is composed of mainly phosphatidylethanolamine (76–77%), with phosphatidylglycerol and cardiolipin present at 11–12% each. Both the IMM and
E. coli membranes are dominated by C16 and C18 acyl chain lengths, with the main differences being that
E. coli membranes are more saturated and contain C17 cyclic fatty acids [
58]. Cardiolipins, in particular, are mostly (~80%) composed of linoleic acyl chains in mammalian cells [
59], whereas the acyl chains are primarily derived from palmitic acid in bacteria, such as
E. coli [
60].
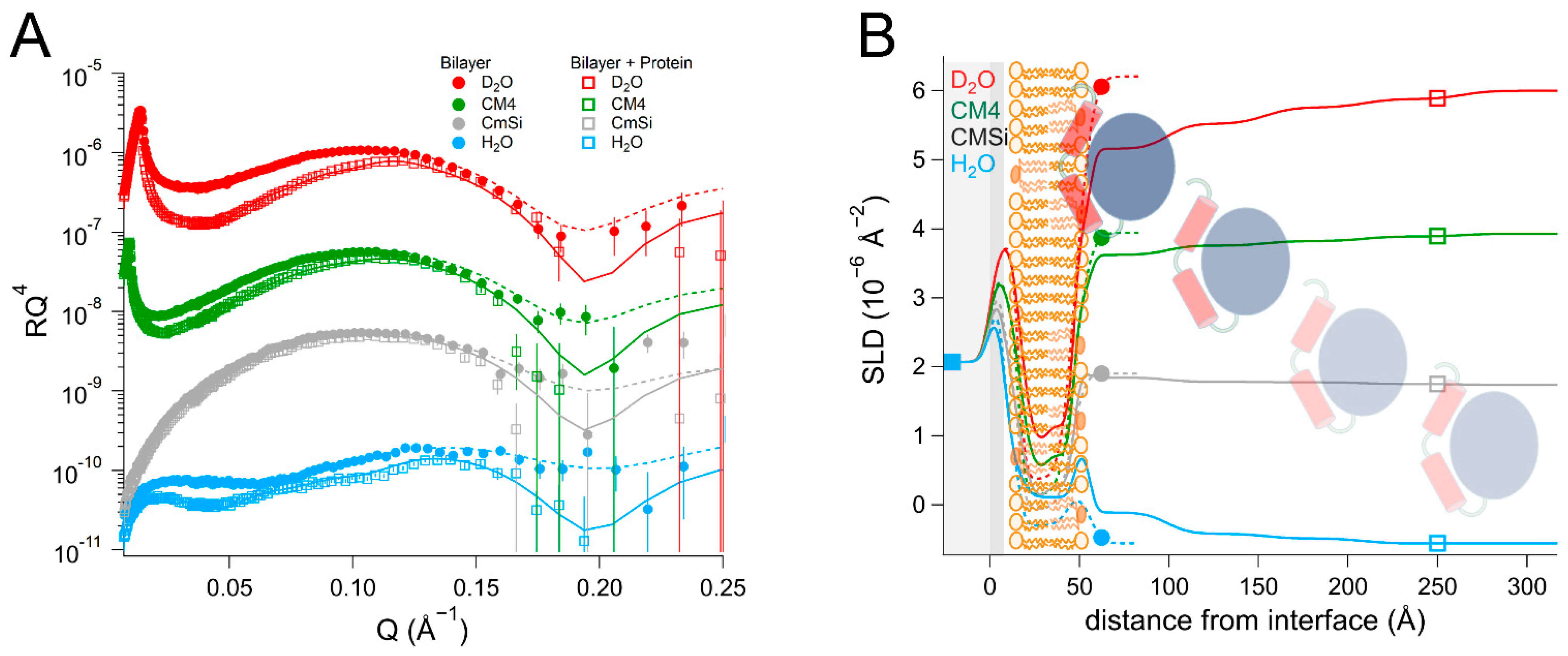
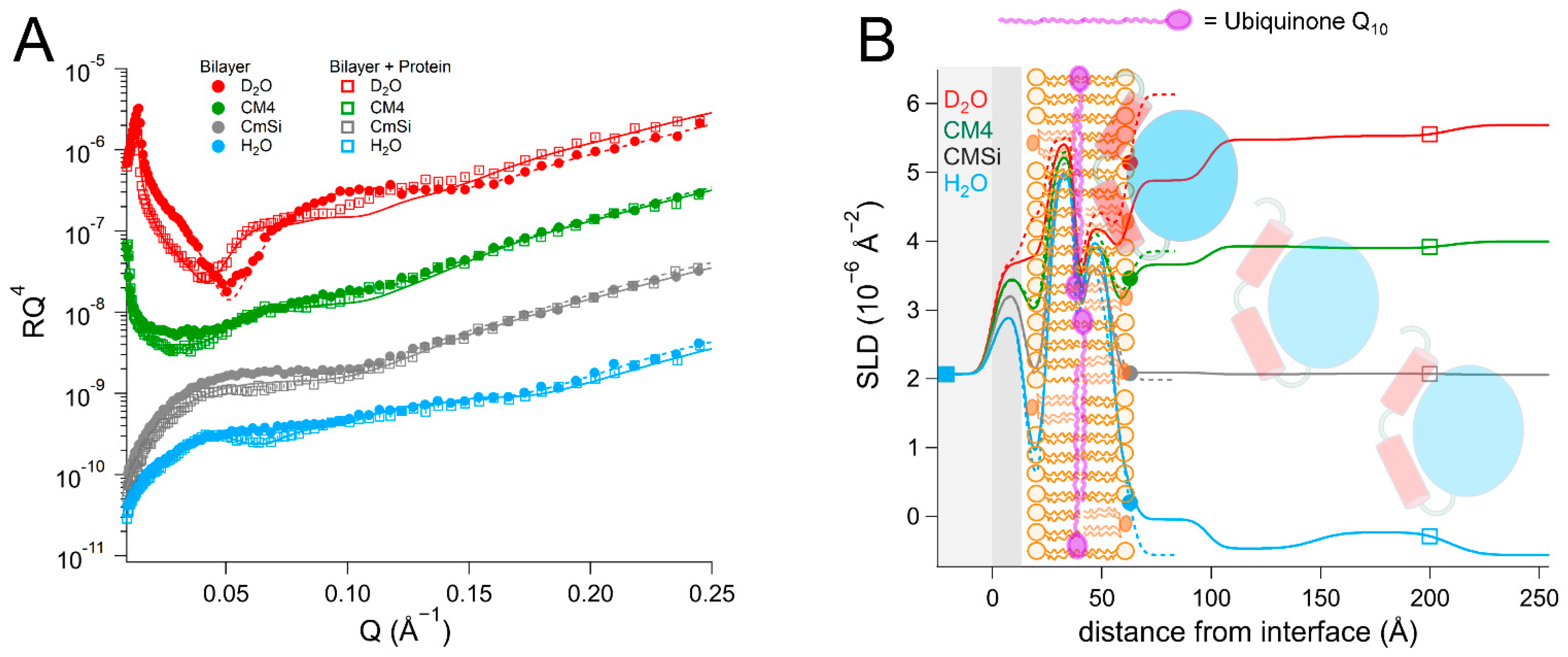
As there are no published data on the interaction of DHODH with lipid membranes, we have focused on a selection of simple model membranes in order to begin to elucidate the relative membrane-binding strength of HsΔ29DHODH and EcDHODH and the dependence on the presence of ubiquinone and some of the lipids found in human mitochondria and the E. coli cytosolic membrane. Our results show that both of the enzymes bind to the membranes containing cardiolipin in the absence and presence of ubiquinone Q10, which is located at the center of the bilayers. EcDHODH binds more efficiently to the surface of simple bilayers consisting of 1-palmitoyl, 2-oleoyl phosphatidylcholine (POPC), and tetraoleoyl cardiolipin (TOCL) compared to HsΔ29DHODH. Q10 is located at the center of all of the lipid bilayers studied, including those prepared from POPC and TOCL, as well as those prepared from more complex lipid mixtures. We also show that both human and bacterial DHODH penetrate into the hydrophobic chain region of the outer lipid leaflet to meet the Q10, which remains in the central layer. We therefore show that the interaction between the enzymes and the membrane-embedded ubiquinone is mediated by enzyme penetration, and not by ubiquinone reorientation. The degree of enzyme penetration depends on the enzyme but also on the lipid composition of the bilayer. We hereby also highlight the importance of the lipid bilayer composition for the enzyme interaction.
3. Discussion
To the best of our knowledge, our study is the first in situ structural investigation of the interaction between lipid bilayers and Class II DHODHs, in which both the lipid and the protein structures are resolved in one dimension. This provides the benefit of observing how the lipid bilayer structures differ based on their composition and how this influences the interaction with the protein.
To date, the interaction of
E. coli DHODH with mixed DOPC/Triton X-100 vesicles has been investigated by electronic spin resonance [
71] and the ability of N-terminally Class II truncated
Plasmodium falciparum DHODH to bind to PC and PE liposomes was shown by size-exclusion chromatography [
72]. Furthermore, the α1-α2 microdomain in
HsDHODH, as an isolated synthetic peptide, has also been studied. Using this peptide, spectroscopic measurements indicate that the α1-α2 microdomain assumes a different conformation in detergent micelles and phospholipid vesicles [
37].
These studies indicate that DHODHs lacking transmembrane domains, such as the one from E. coli, can interact with lipid bilayers and that the α1-α2 microdomain undergoes conformational changes depending on the interaction partner. Therefore, it is likely that the lipid composition plays a major role on the protein–membrane interactions. In this study we set out to investigate this phenomenon. A second question that we addressed is how DHODHs belonging to Class II might interact with the co-substrate, ubiquinone, and how this affects protein–membrane interactions.
By using multidimensional neutron contrast variation of the lipids and the aqueous solvent, we have directly observed the location of ubiquinone in the bilayers, consistent with previous experiments with neutron diffraction [
38], showing that ubiquinone tends to localize at the center of the lipid bilayer in multilamellar lipid stacks and fills the interstitial space in the inverse hexagonal H
II phase of POPE [
73]. Our results clearly indicate a perpendicular orientation of Q
10 relative to the phospholipids. We could show this location in a variety of bilayers, including those that were prepared from complex lipid mixtures derived directly from a eukaryotic organism (
Candida glabrata), translating previous findings with two synthetic lipids alone [
38] to a more complex and physiologically relevant setting. Our most significant finding is that the binding of both
HsΔ29DHODH and
EcDHODH to the surface of the lipid bilayers does not result in the migration or reorientation of ubiquinone from the middle layer towards the membrane–water interface where the protein is located. Instead, our data show that, upon binding to the lipid bilayer, both of the enzymes penetrate into the outer lipid chain region, both in the absence and presence of Q
10. A direct interaction between Q
10 and the enzyme is suggested by the increased retention upon rinsing that
HsΔ29DHODH displays on POPC/TOCL bilayers containing Q
10 in comparison to those without. This effect was both observed in our study here and in our previous study [
43] using QCM-D. The location of ubiquinone at the center of the lipid bilayers and the lack of observable reorientation in the bilayer to bind DHODH are consistent with its large molecular size and branched chain structure, which make it poorly soluble in the phospholipids.
In most cases, the inner protein layer on the surface of the lipid bilayer is somewhat thinner (35–46 Å) than the protein dimensions in the crystal structure and suggest the formation of a monolayer of the protein. However, according to the data analysis, the proteins also penetrate into the outer lipid leaflet by up to 23–24 Å, which would make the total thickness of the first protein layer clearly greater than the crystal structure suggests. Two possible scenarios could explain this. The crystallographic data indicates the α1-α2 microdomains of both of the enzymes (
Figure 1B) are connected to their respective core catalytic domains by potentially flexible loops. Movements in this loop have been shown upon inhibitor binding to N-terminal truncated
HsDHODH and reveal the possibility of conformational flexibility in this region of DHODHs [
15]. Although there are no published reports of this to date, it is likely that the amphipathic α1-α2 microdomain penetrates into the outer lipid chain region, whereas the rest of the catalytic domain remains on the surface of the bilayer, such a conformational rearrangement is facilitated by the flexible loop, as mentioned previously. Alternatively, it is possible that two partially overlapping layers of protein are in contact with the lipid bilayer, one penetrating into the lipids and one on the surface. Our NR data is consistent with both of these scenarios, however it could also be explained by a combination of α1-α2 microdomain penetration and a second, partially overlapping protein layer on the membrane surface. The higher thicknesses of the outer protein layers are less defined, due to the very high solvent fraction (>90 vol% water) and do not necessarily portray individual protein layers.
The observation that both
HsΔ29DHODH and
EcDHODH clearly penetrate into the lipid bilayer to a significant extent is new, to our best knowledge, and indicates that the interaction of the α1-α2 microdomain with lipids has perhaps a larger role to play in the membrane–DHODH interaction than previously thought. This is particularly interesting in the context of the present comparison of the naturally soluble
EcDHODH for which this is the main interaction with membrane lipids, with the truncated
HsΔ29DHODH, which in vivo has a transmembrane domain anchoring it to the IMM. The differences that were observed in the membrane-binding strength and reversibility of the lipid interaction for the two enzymes support a stronger ability of
EcDHODH to bind to the bacterial plasma membrane using only the alpha-helical domain, whereas
HsΔ29DHODH also requires the TM to remain in the IMM. The model of enzyme penetration towards the ubiquinone located at the center of lipid bilayers (
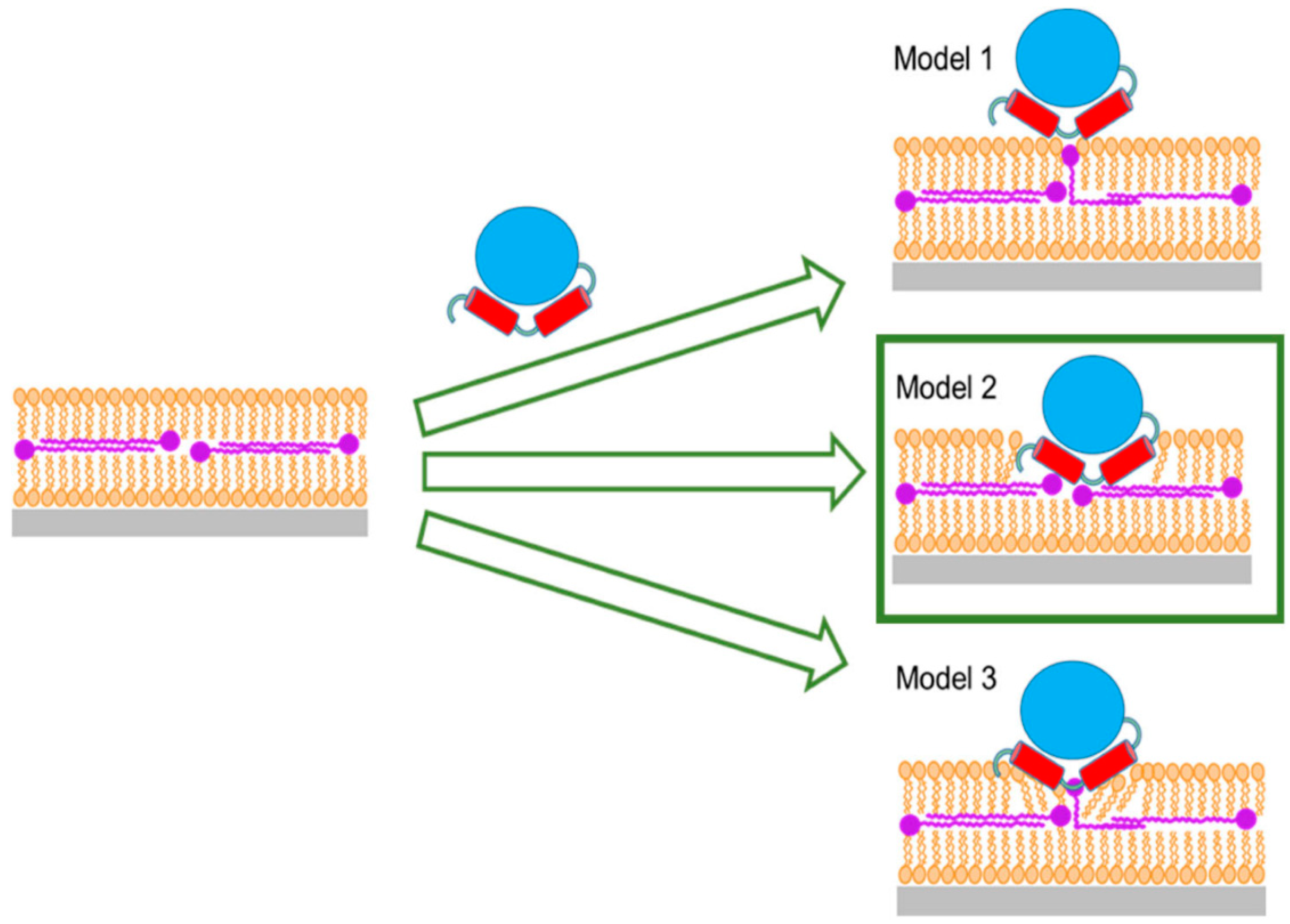
Figure 9) could be relevant for other enzymes that use ubiquinones as electron acceptors. Ubiquinones are found in all membranes, but Q
10 is the key node in the mitochondrial respiratory chain and a substrate for other enzymes comparable to DHODH, such as succinate dehydrogenase, glycerol-3-phosphate dehydrogenase, or electron transfer flavoprotein coenzyme Q, located at the IMM [
74,
75]. In summary, our results provide evidence suggesting that the protein–ubiquinone interaction is facilitated by penetration of the enzyme into the outer lipid chain region (Model 2 in
Figure 9), and, not to any degree observable in our experiments using NR, by migration of ubiquinone from the middle layer towards the outer lipid chain with enzyme partially, or not at all penetrating.
The presence of ubiquinone increases the binding of
HsΔ29DHODH to bilayers consisting of synthetic lipids (POPC, TOCL), as indicated by a more densely populated innermost protein layer (containing less solvent), which is also more stable, as indicated by a lower degree of protein removal by rinsing. Lipid composition also has an effect on the binding of
HsΔ29DHODH to the lipid bilayers. The bilayers consisting of a eukaryotic phospholipid mixture that were derived from yeast display a significantly higher protein binding ability compared to the bilayers that were prepared from synthetic lipids. However, incorporation of Q
10 into bilayers prepared with this complex mixture of lipids does not result in a significantly increased binding of the enzyme or a higher retention, as is the case with synthetic lipid bilayers. Our results are consistent with previous studies using non-denaturing electrospray ionization mass spectrometry (nESI-MS) that have inferred that an N-terminally truncated
HsDHODH, resembling our
HsΔ29DHODH, displays a higher relative binding to lipids, such as TOCL and POPE, compared to POPC [
40], as they are detected by MS in protein–lipid complexes.
We previously demonstrated, by using QCM-D experiments, that both
HsΔ29DHODH and
EcDHODH bind much more strongly to POPC bilayers that contain 10 mol% of TOCL, whereas in its absence the binding of both of the enzymes is reversible [
43]. As the lipid chains are similar in both POPC and TOCL, this suggests that the cardiolipin headgroup is predominantly responsible for this effect. We therefore focused, in this study, on the effect of the lipid headgroups and the electrostatic interaction with the enzyme, by matching the acyl chains of the CL, PE, and PG to the POPC-based samples. The composition of the yeast lipid bilayer that was used was 52 mol% phosphatidylcholine (PC), 27 mol% phosphatidylserine (PS), 14 mol% phosphatidylethanolamine (PE), 4 mol% phosphatidylinositol (PI), and 3 mol% cardiolipin (CL) [
69], and the fatty acid composition of the mixture was as follows (mol%): 40% oleic, 38% palmitoleic, 11% stearic, 6% palmitic, 4% linoleic, and 2% linolenic [
69]. The CL in
C. glabrata has the native acyl chain distribution similar to the overall mixture, apart from the increased linoleic acid (37.4% oleic, 33.7% palmitoleic, 11.3% linoleic, 9.1% palmitic, and 8.6% stearic). Compared to the 90 mol% POPC/10 mol% TOCL and the 80 mol% POPC/10 mol% TOCL/10 mol% Q
10 bilayers, the yeast lipid mixture contains fewer neutral lipids (POPC) and more negatively-charged lipids, such as PS and CL. In line with what has been suggested by Costeira-Paulo et al. [
40], we hypothesize that the electrostatic interactions between these negatively-charged lipid headgroups and the cationic residues present in the amphipathic α1-α2 microdomain of
HsΔ29DHODH are likely to be the main drivers for protein–membrane interaction. However, as there are also differences in the lipid chain composition of the cardiolipins found in human mitochondria and in
E. coli, with the latter being more saturated, an additional role for the lipid chains in the binding that was observed here for the two truncated enzymes can certainly not be excluded and deserves to be investigated in future studies.
Our results indicate that the bacterial EcDHODH displays a higher degree of binding to POPC and TOCL compared to the truncated human enzyme (HsΔ29DHODH). The interaction between the bacterial enzyme and the lipids is both stronger and more stable. The presence of ubiquinone in the lipid bilayer does not significantly increase the binding of EcDHODH. The lipid complexity does have an effect on EcDHODH binding, as its binding to bilayers, mimicking the composition of the bacterial plasma membrane (i.e., containing POPE and POPG), is even higher compared to only POPC and TOCL.
In order to examine the structural features of Class II DHOHDs, we searched the UniProt Database (accessed 16 December 2020) for Class II DHODHs with available X-ray crystal structures using the following query search in PDB: “dihydroorotate dehydrogenase quinone database: (type:pdb)”. From the returned eight hits, three (the DHODHs from
Eimeria tenella,
Helicobacter pylori, and
Mycobacterium tuberculosis) had obsolete (not available) PDB entries, or did not cover the full α1-α2 microdomain. The remaining five sequences were used to perform a multiple sequence alignment (
Figure 10 and
Figure S3).
We hypothesize that the higher relative binding displayed by the bacterial enzyme may arise from the presence of an abundance of positively charged residues on the outer surface of the α1-α2 microdomain of
EcDHODH (Arg7, Lys8, Arg17, Arg27, and Arg28), which are likely to be in direct contact with the lipid bilayer. This is supported by the comparison of the amino acid sequences in
Figure 10. The truncated human enzyme also possesses cationic residues in the corresponding region, but they are fewer in number (Arg35, Arg56, and Arg60). Thus, the human enzyme may rely to a greater extent on the presence of the transmembrane domain in order to achieve a stable interaction with the lipid bilayer, as opposed to the bacterial enzyme, which lacks such a structure and is also in need of using other electron acceptors besides membrane-embedded ubiquinones.
It is, however, interesting to note that the distribution of the cationic residues in the α1-α2 microdomain does not seem highly conserved in the Class II DHODHs in
Figure 10, spanning a wide evolutionary distance. This conclusion is in line with a recent comprehensive bioinformatics study by Sousa et al. [
76]. None of the positively charged residues on the outer surface of the α1-α2 microdomain were pointed out as highly conserved in an alignment of 1062 Class II DHODH sequences, or the whole α1-α2 microdomain, was one of the least conserved regions of the enzymes. It might be that the N-terminal part of DHODH has evolved to match the lipid composition of its respective cellular environment. In eukaryotes, together with the mitochondrial location, Class II DHODH have acquired a transmembrane domain, that anchors their position to the outer IMM. What other functions and benefits this transmembrane domain might have, is still open for discovery.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}