Is the Distribution of Two Rare Orchis Sister Species Limited by Their Main Mycobiont?


Abstract
:
{kind=link}
{kind=link}
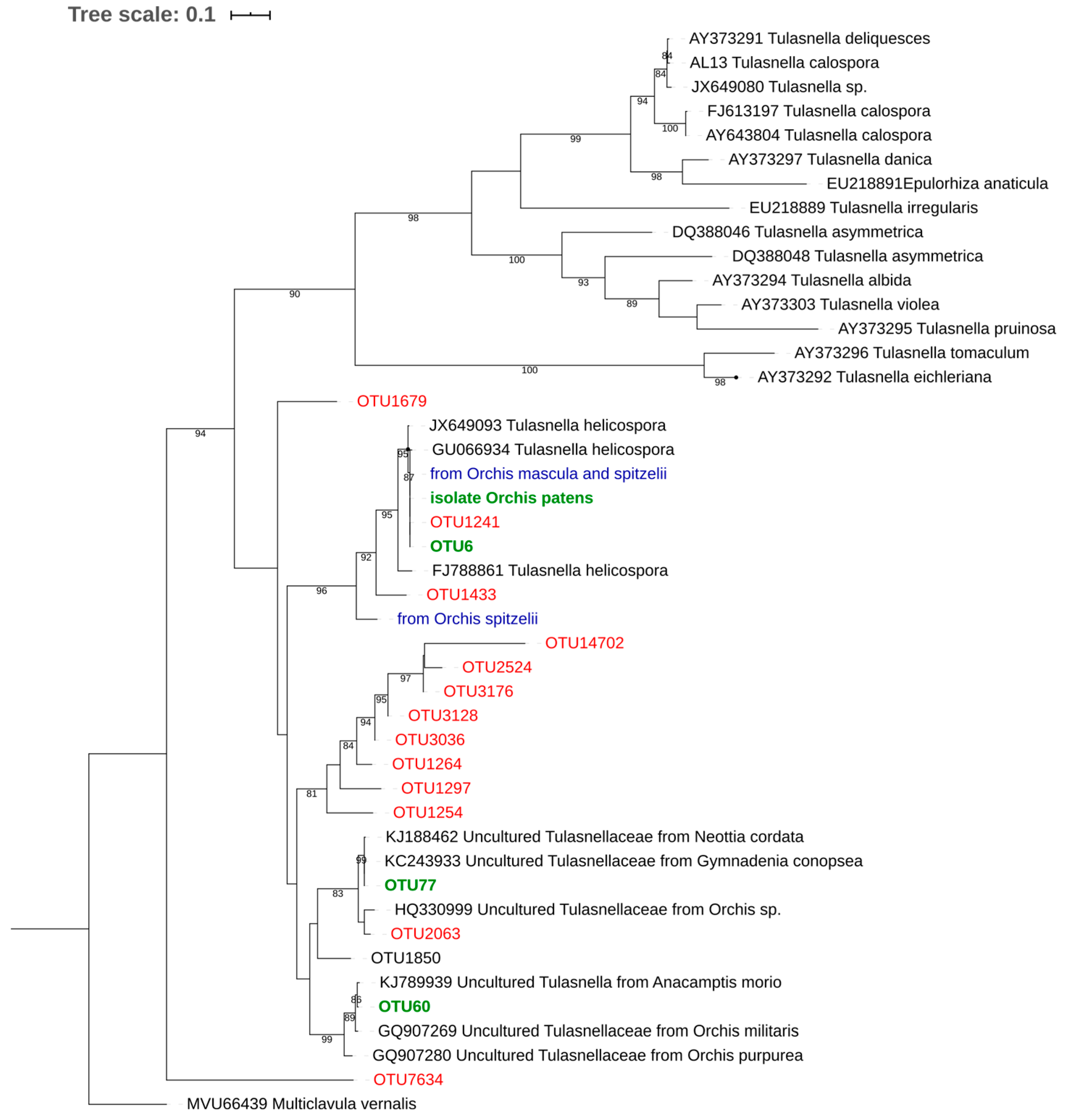{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Species and Material
2.2. Molecular Analysis of Mycorrhizal Fungi
2.2.1. DNA Extraction from Roots
2.2.2. Cloning and Fungal Amplification from Roots
2.2.3. Sampling, DNA Extraction and Amplification from Soil
2.2.4. Amplicon Purification and Illumina Sequencing
2.3. Fungal Isolation and Germination Trials
2.4. Data analysis and Bioinformatics
2.4.1. Bioinformatic Analysis
2.4.2. Phylogenetic Analysis
2.4.3. Statistical Analyses
3. Results
3.1. Identification of OMF in O. patens and O. canariensis
3.2. OM Fungal Diversity in the Different Orchid Populations
3.3. Amplification of Fungi From Soil
3.4. In Vitro Trials
4. Discussion
4.1. Specificity of Mycorrhizal Associations of the Two Orchis Sister Species
4.2. Distribution of Tulasnella helicospora in Soil
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jaquemyn, H.; Duffy, K.J.; Selosse, M.A. Biogeography of orchid Mycorrhizas. In Biogeography of Mycorrhizal Symbiosis. Ecological Studies; Tedersoo, L., Ed.; Springer: Cham, Switzerland, 2017; p. 230. [Google Scholar] [CrossRef]
- Ettinger, A.K.; HilleRisLambers, J. Climate isn’t everything: Competitive interactions and variation by life stage will also affect range shifts in a warming world. Am. J. Bot. 2013, 100, 1344–1355. [Google Scholar] [CrossRef] [Green Version]
- Afkhami, M.E.; McIntyre, P.J.; Strauss, S.Y. Mutualist-mediated effects on specie’s range limits across large geographic scales. Ecol. Lett. 2014, 17, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Duffy, K.J.; Johnson, S.D. Male interference with pollination efficiency in a hermaphroditic orchid. J. Evol. Biol. 2014, 27, 1751–1756. [Google Scholar] [CrossRef] [PubMed]
- Gioria, M.; Osborne, B.A. Resource competition in plant invasions: Emerging patterns and research needs. Front. Plant Sci. 2014, 5, 501. [Google Scholar] [CrossRef] [Green Version]
- Swarts, N.D.; Dixon, K.W. Conservation Methods for Terrestrial Orchids; J. Ross Publishing: Plantation, FL, USA, 2017. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: Cambridge, UK, 2008. [Google Scholar]
- Dearnaley, J.D.W.; Martos, F.; Selosse, M.A. Orchid mycorrhizas: Molecular ecology, physiology, evolution, and conservation aspects. In Fungal ASSociations, 2rd ed.; Hock, B., Ed.; Springer: Berlin, Germany, 2012; pp. 207–230. [Google Scholar]
- Waud, M.; Brys, R.; Van Landuyt, W.; Lievens, B.; Jacquemyn, H. Mycorrhizal specificity does not limit the distribution of an endangered orchid species. Mol. Ecol. 2017, 26, 1687–1701. [Google Scholar] [CrossRef] [PubMed]
- Oktalira, F.T.; Whitehead, M.R.; Linde, C.C. Mycorrhizal specificity in widespread and narrow-range distributed Caladenia orchid species. Fungal Ecol. 2019. [Google Scholar] [CrossRef]
- McCormick, M.K.; Whigham, D.F.; Sloan, D.; O’Malley, K.; Hodkinson, B. Orchid–fungus fidelity: A marriage meant to last? Ecology 2006, 87, 903–911. [Google Scholar] [CrossRef]
- Bidartondo, M.I.; Read, D.J. Fungal specificity bottlenecks during orchid germination and development. Mol. Ecol. 2008, 17, 3707–3716. [Google Scholar] [CrossRef] [Green Version]
- Jacquemyn, H.; Merckx, V.; Brys, R.; Tyteca, D.; Cammue, B.P.A.; Honnay, O.; Lievens, B. Analysis of network architecture reveals phylogenetic constraints on mycorrhizal specificity in the genus Orchis (Orchidaceae). New Phytol. 2011, 192, 518–528. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Dixon, K.W.; Jersakova, J.; Tesitelova, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [Green Version]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- McCormick, M.K.; Whigham, D.F.; Canchani-Viruet, A. Mycorrhizal fungi affect orchid distribution and population dynamics. New Phytol. 2018, 219, 1207–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, M.F. Orchid conservation: How can we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shefferson, R.P.; Bunch, W.; Cowden, C.C.; Lee, Y.I.; Kartzinel, T.R.; Yukawa, T.; Downing, J.; Jiang, H. Does evolutionary history determine specificity in broad ecological interactions? J. Ecol. 2019, 107, 1582–1593. [Google Scholar] [CrossRef]
- Combes, C. Parasitism: The Ecology and Evolution of Intimate Inter-Actions; University of Chicago Press: Chicago, IL, USA, 2004. [Google Scholar]
- Davis, B.J.; Phillips, R.D.; Wright, M.; Linde, C.C.; Dixon, K.W. Continent-wide distribution in mycorrhizal fungi: Implications for the biogeography of specialized orchids. Ann. Bot. 2015, 116, 413–421. [Google Scholar] [CrossRef]
- Otero, J.T.; Flanagan, N.S.; Herre, E.A.; Ackerman, J.D.; Bayman, P. Widespread mycorrhizal specificity correlates to mycorrhizal function in the neotropical, epiphytic orchid Ionopsis utricularioides (Orchidaceae). Am. J. Bot. 2007, 94, 1944–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selosse, M.A.; Weiß, M.; Jany, J.L.; Tillier, A. Communities and populations of sebacinoid basidiomycetes associated with the achlorophyllous orchid Neottia nidus-avis (L.) L.C.M. Rich. and neighboring tree ectomycorrhizae. Mol. Ecol. 2002, 11, 1831–1844. [Google Scholar] [CrossRef]
- Taylor, D.L.; Bruns, T.D.; Hodges, S.A. Evidence for mycorrhizal races in a cheating orchid. Proc. Biol. Sci. 2004, 271, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Girlanda, M.; Selosse, M.A.; Cafasso, D.; Brilli, F.; Delfine, S.; Fabbian, R.; Ghihnone, P.; Pinelli, R.; Segreto, F.; Loreto, S.; et al. Inefficient photosynthesis in the Mediterranean orchid Limodorum abortivum (L.) Swartz is mirrored by specific association to ectomycorrhizal Russulaceae. Mol. Ecol. 2006, 15, 491–504. [Google Scholar] [CrossRef]
- Irwin, M.J.; Bougoure, J.J.; Dearnaley, J.D.W. Pterostylis nutans (Orchidaceae) has a specific association with two Ceratobasidium root-associated fungi across its range in eastern Australia. Mycoscience 2007, 48, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Roy, M.; Yagame, T.; Yamato, M.; Iwase, K.; Heinz, C.; Faccio, A.; Bonfante, P.; Selosse, M.A. Ectomycorrhizal Inocybe species associate with the mycoheterotrophic orchid Epipogium aphyllum but not its asexual propagules. Ann. Bot. 2009, 104, 595–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Těšitelová, T.; Kotilínek, M.; Jersáková, J.; Joly, F.X.; Košnar, J.; Tatarenko, I.; Selosse, M.A. Two widespread green Neottia species (Orchidaceae) show mycorrhizal preference for Sebacinales in various habitats and ontogenetic stages. Mol. Ecol. 2015, 24, 1122–1134. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Gao, Y.; Zhao, Z.; Waud, M.; Duffy, K.J.; Selosse, M.A.; Jakalski, M.; Liu, N.; Jacquemyn, H.; Guo, S. Similarity in mycorrhizal communities associating with two widespread terrestrial orchids decays with distance. J. Biogeogr. 2019. [Google Scholar] [CrossRef]
- Duffy, K.J.; Waud, M.; Schatz, B.; Petanidou, T.; Jacquemyn, H. Latitudinal variation in mycorrhizal diversity associated with a European orchid. J. Biogeogr. 2019, 46, 968–980. [Google Scholar] [CrossRef]
- Pandey, M.; Sharma, J.; Taylor, D.L.; Yadon, V.L. A narrowly endemic photosynthetic orchid is non-specific in its mycorrhizal associations. Mol. Ecol. 2013, 22, 2341–2354. [Google Scholar] [CrossRef]
- Kaur, J.; Andrews, L.; Sharma, J. High specificity of a rare terrestrial orchid toward a rare fungus within the North American tallgrass prairie. Fungal Ecol. 2019, 123, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M.; Hollingsworth, P.M.; Preston, J.; Yi-Bo, L.; Pridgeon, A.M.; Chase, M.W. Molecular phylogenetics and evolution of Orchidinae and selected Habenariinae (Orchidaceae). Bot. J. Linn. Soc. 2003, 142, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Rankou, H. Orchis patens, green spotted Orchid. In The IUCN Red List of Threatened Species 2018; Available online: https://dx.doi.org/10.2305/IUCN.UK.2018-2.RLTS.T175961A84665334.en (accessed on 10 December 2019).
- Calevo. Newly developed nuclear microsatellite markers provide molecular evidence for species and subspecies level distinctions in the genus Orchis. Manuscript in preparation.
- Taylor, D.L.; McCormick, M.K. Internal transcribed spacer primers and sequences for improved characterization of basidiomycetous orchid mycorrhizas. New Phytol. 2008, 177, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Voyron, S.; Ercole, E.; Ghignone, S.; Perotto, S.; Girlanda, M. Fine-scale spatial distribution of orchid mycorrhizal fungi in the soil of host-rich grasslands. New Phytol. 2017, 213, 1428–1439. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Ercole, E.; Rodda, M.; Girlanda, M.; Perotto, S. Establishment of a symbiotic in vitro system between a green meadow Orchid and a Rhizoctonia-like Fungus. Bioprotocol 2015, 5. [Google Scholar] [CrossRef]
- Zang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Hoiland, K.; Kjoller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 3030, 1312–1313. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. Available online: http://cran.r-project.org/package=vegan (accessed on 11 December 2019).
- The R Development Core Team. R: A Language and Environment for Statistical Computing (Version 2.6.2). Available online: http://www.R-project.org (accessed on 12 September 2019).
- Rasmussen, H.N.; Rasmussen, F.N. Seedling mycorrhiza: A discussion of origin and evolution in Orchidaceae. Bot. J. Linn. Soc. 2014, 175, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef]
- Orgiazzi, A.; Bianciotto, V.; Bonfante, P.; Daghino, S.; Ghignone, S.; Lazzari, A.; Lumini, E.; Mello, A.; Napoli, C.; Perotto, S.; et al. 454 pyrosequencing analysis of fungal assemblages from geographically distant, disparate soils reveals spatial patterning and a core mycobiome. Diversity 2013, 5, 73–98. [Google Scholar] [CrossRef] [Green Version]
- Orgiazzi, A.; Lumini, E.; Nilsson, R.H.; Girlanda, M.; Vizzini, A.; Bonfante, P.; Bianciotto, V. Unravelling soil fungal communities from different Mediterranean land-use backgrounds. PLoS ONE 2012, 7, e34847. [Google Scholar] [CrossRef]
- Schmidt, P.A.; Balint, M.; Greshake, B.; Bandow, C.; Römbke, J.; Schmitt, I. Illumina metabarcoding of a soil fungal community. Soil Biol. Biochem. 2013, 65, 128–132. [Google Scholar] [CrossRef]
- Waud, M.; Busschaert, P.; Lievens, B.; Jacquemyn, H. Specificity and localized distribution of mycorrhizal fungi in the soil may contribute to co-existence of orchid species. Fungal Ecol. 2016, 20, 155–165. [Google Scholar] [CrossRef]
- Oberwinkler, F.; Riess, K.; Bauer, R.; Selosse, M.A.; Weiss, M.; Garnica, S.; Zuccaro, A. Enigmatic Sebacinales. Mycol. Prog. 2013, 12, 1–27. [Google Scholar] [CrossRef]
- Phillips, R.D.; Barret, M.D.; Dalziell, E.L.; Dixon, K.W.; Swarts, N.D. Geographical range and host breadth of Sebacina orchid mycorrhizal fungi associating with Caladenia in South-Western Australia. Bot. J. Linn. Soc. 2016, 1, 140–151. [Google Scholar] [CrossRef]
- Waud, M.; Wiegand, T.; Brys, R.; Lievens, B.; Jacquemyn, H. Nonrandom seedling establishment corresponds with distance-dependent decline in mycorrhizal abundance in two terrestrial orchids. New Phytol. 2016, 211, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemyn, H.; Brys, R.; Waud, M.; Busschaert, P.; Lievens, B. Mycorrhizal networks and coexistence in species-rich orchid communities. New Phytol. 2015, 206, 1127–1134. [Google Scholar] [CrossRef]
- Egidi, E.; May, T.W.; Franks, A.E. Seeking the needle in the haystack: Undetectability of mycorrhizal fungi outside of the plant rhizosphere associated with an endangered Australian orchid. Fungal Ecol. 2018, 33, 13–23. [Google Scholar] [CrossRef]
- Oja, J.; Kohout, P.; Tedersoo, L.; Kull, T.; Koljalg, U. Temporal patterns of orchid mycorrhizal fungi in meadows and forests as revealed by 454 pyrosequencing. New Phytol. 2015, 205, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.A. The latest news from biological interactions in orchids: In love, head to toe. New Phytol. 2014, 202, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.A.; Martos, F. Do chlorophyllous orchids heterotrophically use mycorrhizal fungal carbon? Trends Plant Sci. 2014, 19, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Ruibal, M.P.; Peakall, R.; Smith, L.M.; Linde, C.C. Phylogenetic and microsatellite markers for Tulasnella (Tulasnellaceae) mycorrhizal fungi associated with Australian orchids. Appl. Plant Sci. 2013, 11. [Google Scholar] [CrossRef]
- Ruibal, M.P.; Triponez, Y.; Smith, L.M.; Peakall, R.; Linde, C.C. Population structure of an orchid mycorrhizal fungus with genus-wide specificity. Sci. Rep. 2017, 7, 5613. [Google Scholar] [CrossRef]
- Whitehead, M.R.; Catullo, R.A.; Ruibal, M.; Dixon, K.W.; Peakall, R.; Linde, C.C. Evaluating multilocus Bayesian species delimitation for discovery of cryptic mycorrhizal diversity. Fungal Ecol. 2017, 26, 74–84. [Google Scholar] [CrossRef]
- Duffy, K.J.; Johnson, S.D. Specialized mutualisms may constrain the geographical distribution of flowering plants. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Factors affecting the distribution and abundance of orchids in grasslands and herbaceous wetlands. Syst. Biodivers. 2016, 1414, 355–370. [Google Scholar] [CrossRef]
- Fuji, M.; Miura, C.; Yamamoto, T.; Komiyama, S.; Suetsugu, K.; Yahame, T.; Yamato, M.; Kaminaka, H. Relative effectiveness of Tulasnella fungal strains in orchid mycorrhizal symbioses between germination and subsequent seedling growth. Symbiosis 2020. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calevo, J.; Voyron, S.; Ercole, E.; Girlanda, M. Is the Distribution of Two Rare Orchis Sister Species Limited by Their Main Mycobiont? Diversity 2020, 12, 262. https://doi.org/10.3390/d12070262
Calevo J, Voyron S, Ercole E, Girlanda M. Is the Distribution of Two Rare Orchis Sister Species Limited by Their Main Mycobiont? Diversity. 2020; 12(7):262. https://doi.org/10.3390/d12070262
Chicago/Turabian StyleCalevo, Jacopo, Samuele Voyron, Enrico Ercole, and Mariangela Girlanda. 2020. "Is the Distribution of Two Rare Orchis Sister Species Limited by Their Main Mycobiont?" Diversity 12, no. 7: 262. https://doi.org/10.3390/d12070262
APA StyleCalevo, J., Voyron, S., Ercole, E., & Girlanda, M. (2020). Is the Distribution of Two Rare Orchis Sister Species Limited by Their Main Mycobiont? Diversity, 12(7), 262. https://doi.org/10.3390/d12070262