Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information


Abstract
:1. Introduction
2. Materials and Methods
3. Results
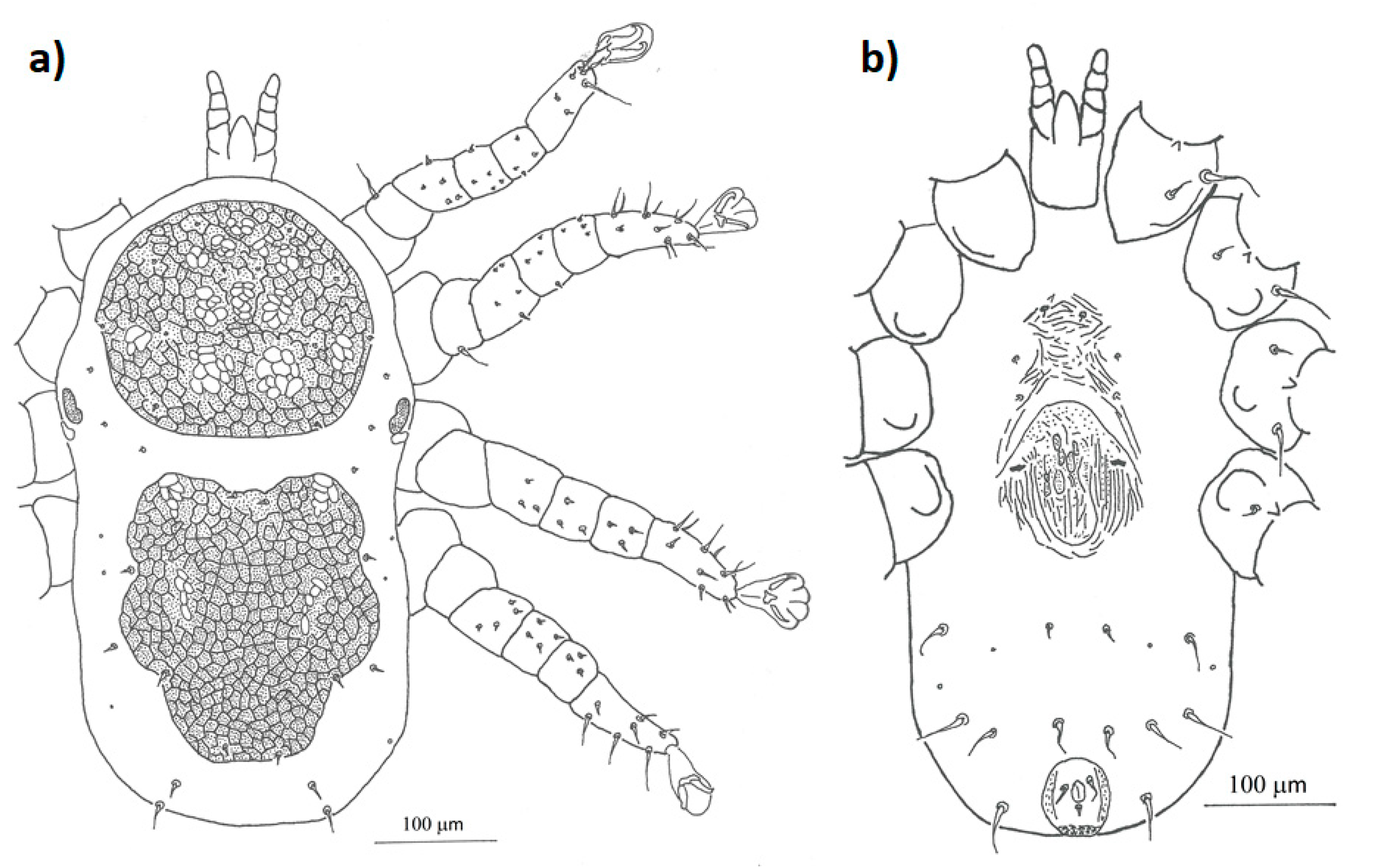
3.1. Morphological Analysis
- Genus Mesonyssus Fain, 1960 [35]. Mesonyssus [14,21,23,35,36,37,38]. Tinaminyssus [24,25,39,40,41]. Type species: Neonyssus treronis [42].Diagnosis: Idiosoma oval, 450–630 μm long. Dorsal side of idiosoma with podosomal and opisthosomal shields. Pygidial shield absent. Stigmata with oblong peritrems located dorsolaterally. Mesosomal shields absent. Poststigmal sclerites present or absent. Gnathosoma located terminally. Deutosternal denticles absent. Tritosternum absent. Sternal shield absent. Genital shield present. Anal shield surrounding anus, located ventrally. Cribrum (Aspero) present. Convexities and spines on coxae present or absent.
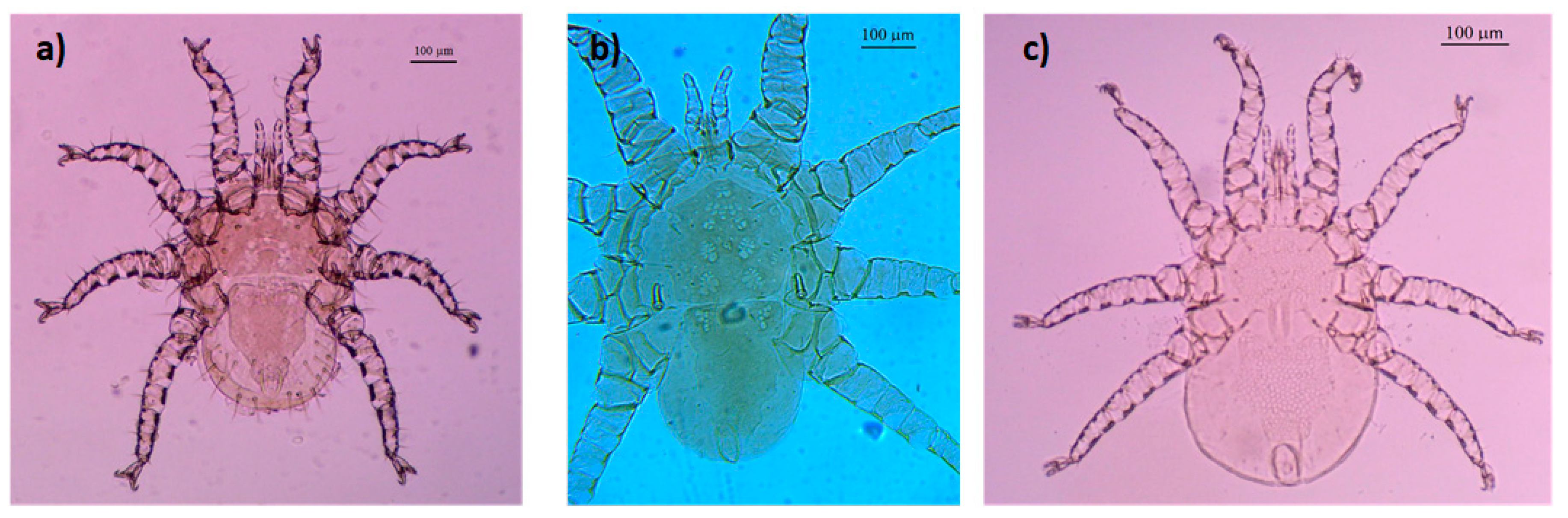
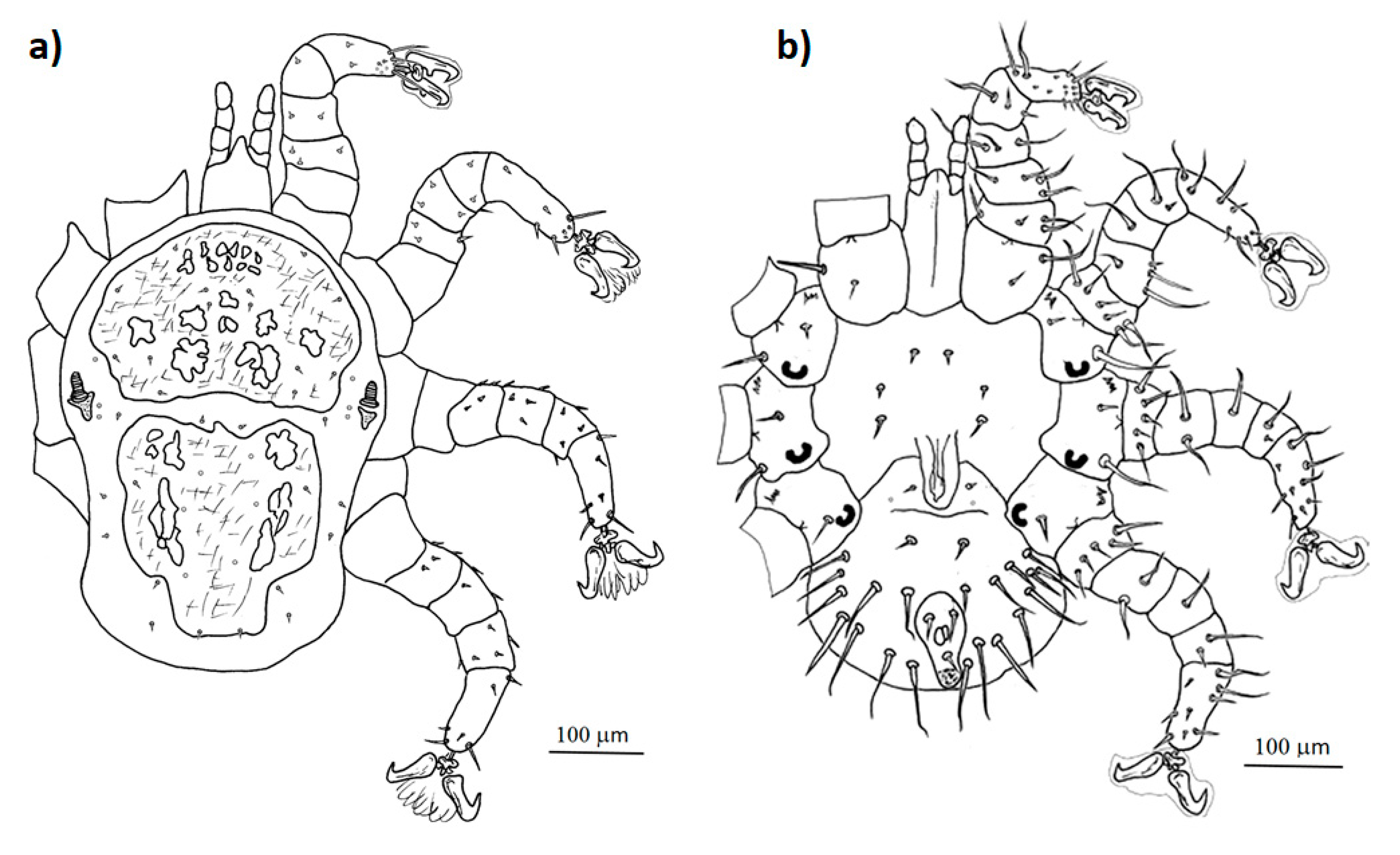
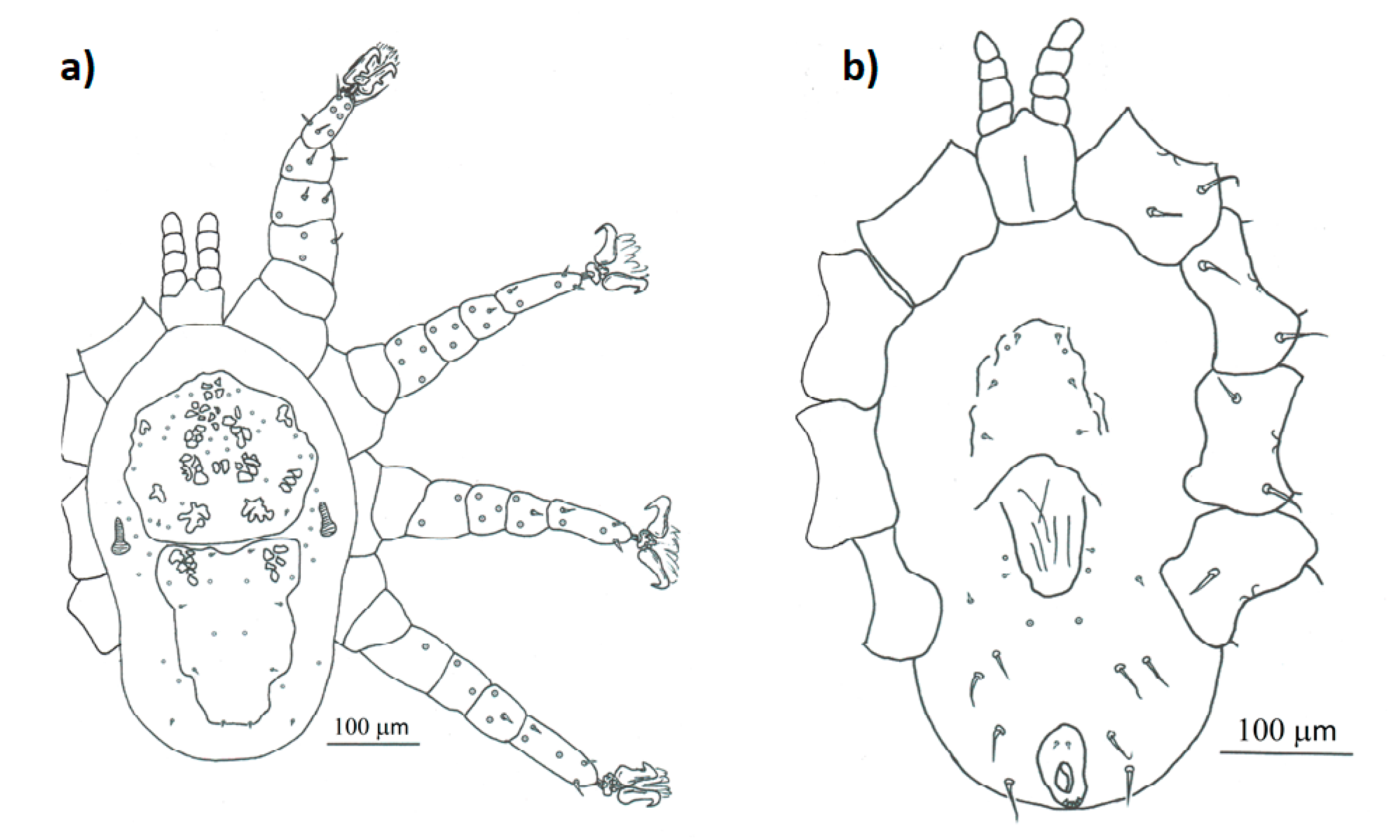
- Mesonyssus melloi (Castro, 1948) [43] (Figure 2a). Neonyssus melloi [22,43,44]. Mesonyssus melloi [14,21,27,45]. Tinaminyssus melloi [24,25,26,40]. Mesonyssus melloi melloi [23]. Neonyssus hirsutus [46].Dorsum (Figure 3a): Podosomal shield wide, strongly sclerotized, with anterior margin convex, almost semi-circular. Podosomal shield shows seven pairs of setae. Stigmata with short peritrems, located dorsolaterally, at level of coxae III. Post-stigmatic sclerites of triangular shape. On podosoma cuticle three pairs of mesolateral setae can be observed. Opisthosomal cuticle presents six pairs of fine setae. Opisthosomal shield wide, large, occupies almost entire opisthosoma. Anterior margin of this shield sinuous, and includes three pairs of pores and two pairs of setae.Ventrum: (Figure 3b) Sternal shield absent. three pairs of sternal setae present; Genital shield of medium size, thin and narrow. A pair of genital setae situated lateral to genital shield. Ventral opisthosoma with a pair of short setae Jv1 and 10 pairs of long setae. Anal shield pear-shaped, broadened anteriorly, bearing a pair of preanal setae, located anterior to anus, and one posterior postanal seta Cribrum (Aspero) present. Coxae I–III with two setae of different length. Posterior parts of coxae II–IV convexitas coxae. Anterior part of coxae II–IV with spina coxae.Gnathosoma: Oblong. Gnathosomal formula: 2-4-2. Three pairs of hypostomal setae and one pair of subcapitular setae present.
- Mesonyssus columbae (Crossley, 1950) [47] (Figure 2b). Neonyssus columbae [22,47]. Mesonyssus columbae [14,21,23,45]. Tinaminyssus columbae [24,25,40].Dorsum (Figure 4a): Podosomal and opisthosomal shields strongly sclerotized, which present on their surface 26 setae. Stigmata with peritrems, located at level of coxae III. The opisthosomal shield shows four pairs of setae and three pairs of pores. Dorsal podosomal cuticle includes three pairs of mesolateral setae. Dorsal opisthosomal cuticle presents four pairs of setae.Ventrum (Figure 4b): Sternal shield absent. Three pairs of sternal setae present. Genital shield wide, one pair of genital setae and one pair of pores situated lateral to this shield. Ventral opisthosomal idiosoma with six pairs of setae. Anal shield narrow oval and includes one pair of preanal setae. Aspero present.Gnathosoma: Wide. Gnathosomal formula (2-4-0). Subcapitular setae absent.
- Mesonyssus streptopeliae Fain, 1962 [18] (Figure 2c). Mesonyssus melloi streptopeliae [18]. Mesonyssus streptopeliae [20,21,27,28].Dorsum (Figure 5a): Podosomal and opisthosomal shields strongly sclerotized. Podosomal shield with round form with seven pairs of setae. Stigmata with peritrems, located at level of coxae III. Post-stigmatic sclerites of triangular shape. The opisthosomal shield wave-shaped with four pairs of setae. Dorsal podosomal cuticle includes three pairs of mesolateral setae. Dorsal opisthosomal cuticle presents four long pairs and three short pairs of setae.Ventrum (Figure 5b): There is a subtle rudiment of sternal shield. Three pairs of short sternal setae present. Genital shield wide. Ventral opisthosomal idiosoma with six pairs of setae and four pores. Anal shield round and includes one pair of preanal setae. Aspero absent.Gnathosoma: Thin and oblong. Gnathosomal formula (2-4-2). Palps long, with short and thin setae.
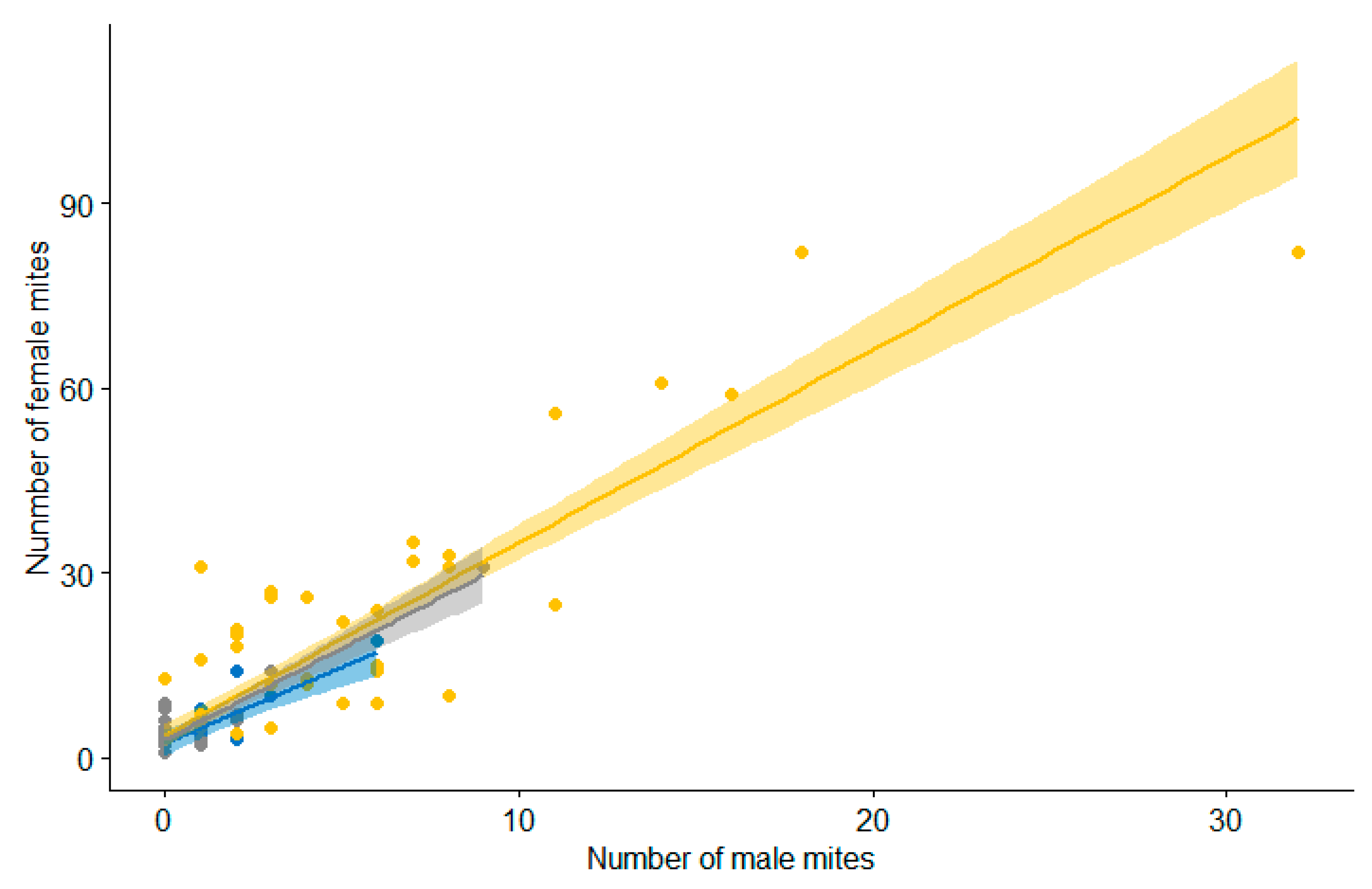
3.2. Epidemiological Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitzthum, H.G. Milben aus der Nasenhöhle von Vögeln. J. Ornithol. 1935, 83, 563–587. [Google Scholar] [CrossRef]
- Dimov, I.D. Morphology of Nasal Mites of the Family Rhinonyssidae—Scanning Electron Microscopy Saint Petersburg 2020; LLC Zhigulin: Saint Petersburg, Russia, 2020; ISBN 9785604468708. [Google Scholar]
- Dimov, I.D. Rhinonyssidosis avium. Vetpharma 2011, 3, 88–90. [Google Scholar]
- Stephan, S.A.R.; Kaschula, V.R.; Canham, A.S. Respiratory acariasis of canaries. J. S. Afr. Vet. Assoc. 1950, 21, 103–107. [Google Scholar]
- Tidemann, S.C.; McOrist, S.; Woinarski, J.C.Z.; Freeland, W.J. Parasitism of wild Gouldian finches (Erythrura gouldiae) by the air-sac mite Sternostoma tracheacolum. J. Wildl. Dis. 1992, 28, 80–84. [Google Scholar] [CrossRef]
- Moro, C.V.; Chauve, C.; Zenner, L. Experimental infection of Salmonella Enteritidis by the poultry red mite, Dermanyssus gallinae. Vet. Parasitol. 2007, 146, 329–336. [Google Scholar] [CrossRef]
- Sommer, D.; Heffels-Redmann, U.; Köhler, K.; Lierz, M.; Kaleta, E.F. Role of the poultry red mite (Dermanyssus gallinae) in the transmission of avian influenza A virus. Tierarztliche Praxis Ausgabe G Grosstiere/Nutztiere 2016, 44, 26–33. [Google Scholar] [CrossRef]
- Osuna-Mascaró, C.; Doña, J.; Johnson, K.P.; Esteban, R.; De Rojas, M. Complete mitochondrial genomes and bacterial metagenomic data from two species of parasitic avian nasal-mites (Rhinonyssidae: Mesostigmata). Front. Ecol. Evol. 2020, 8, 142. [Google Scholar] [CrossRef]
- Feider, Z.; Mironescu, I. Rhinonyssides parasites des oiseaux des genres Podiceps et Tringa. Anaele Stiint. Univ. Al. I. Cuza Iasi Sect. II A 1972, 18, 317–330. [Google Scholar]
- Guevara Benítez, D.C.; Úbeda Ontiveros, J.M. Ácaros del género Ptilonyssus Berlese y Trouessart, 1889 (mesostigmata: Rhynonyssidae) parásitos de fosas nasales de paseriformes españolas. I. Ptilonyssus hirsti (Castro y Pereira, 1947) Pereira y Castro, 1949. Rev. Iber. Parasitol. 1978, 38, 591–614. [Google Scholar]
- Kadosaka, T.; Kaneko, K.; Asanuma, K. A new species and new records of avian nasal mites (Acarina: Rhinonyssidae) from Japan. Jpn. J. Sanit. Zool. 1987, 38, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Spicer, G.S. Nasal mites from birds of a Guatemalan cloud forest (Acarina: Rhinonyssidae). J. Parasitol. 1984, 70, 794–802. [Google Scholar] [CrossRef]
- Da Silva, D.S.; Scheer, S.; Muller, G. Nasal mites (Mesostigmata, Rhinonyssidae) in Sternidae (Aves: Charadriiformes) on the southern coast of Brazil. Rev. Bras. Parasitol. Veterinária 2018, 27, 109–111. [Google Scholar] [CrossRef] [Green Version]
- Dimov, I.D. Kleshchi-rinonissidy ptic Severo-Zapada Ross; LLC Zhigulin: Saint Petersburg, Russia, 2018; ISBN 9785600021778. [Google Scholar]
- Bell, P.J. The life history and transmission biology of Sternostoma tracheacolum Lawrence (Acari: Rhinonyssidae) associated with the Gouldian finch Erythrura gouldiae. Exp. Appl. Acarol. 1996, 20, 323–341. [Google Scholar] [CrossRef]
- Pence, D.B. The nasal mites of birds from Louisiana. IX. Synopsis. J. Parasitol. 1973, 881–892. [Google Scholar] [CrossRef]
- Baulieu, F.; Knee, W.; Nowell, V.; Schwarzfeld, M.; Lindo, Z.; Behan-Pelletier, V.M.; Lumley, L.; Young, M.R.; Smith, I.; Proctor, H.C. Acari of Canada. Zookeys 2019, 77. [Google Scholar] [CrossRef]
- Fain, A. Les Rhinonyssides parasites des Pigeons. Rev. Zool. Bot. Afr. 1962, 65, 305–324. [Google Scholar]
- Bonnefoy, X.; Kampen, H.; Sweeney, K. Public Health Significance of Urban Pests; World Health Organization: Geneva, Switzerland, 2008; ISBN 9289071885. [Google Scholar]
- Fain, A. Diagnoses d’acariens parasites nouveaux. Rev. Zool. Bot. Afr 1965, 72, 152–160. [Google Scholar]
- Butenko, O.M. Rhinonyssid Mites of Non-Passerine Birds of the USSR; Academy of Sciences of USSR: Moscow, Russia, 1984. [Google Scholar]
- Fain, A. Les acariens des familles Epidermoptidae et Rhinonyssidae parasites des fosses nasales d’oiseaux au Ruanda-Urundi et au Congo belge. Proc. Ann. Musée R. Congo Belge 1957, 60, 1–176. [Google Scholar]
- Wilson, N. New records and descriptions of Rhinonyssidae mostly from New Guinea. Pac. Insects 1964, 6, 357–388. [Google Scholar]
- Pence, D.B. Keys, Species and Host List, and Bibliography for Nasal Mites of North. American Birds (Acarina-Rhinonyssinae, Turbinoptinae, Speleognathinae, and Cytoditidae); Texas Tech Press: Lubbock, TX, USA, 1975. [Google Scholar]
- Knee, W.; Proctor, H.; Galloway, T. Survey of nasal mites (Rhinonyssidae, Ereynetidae, and Turbinoptidae) associated with birds in Alberta and Manitoba, Canada. Can. Entomol. 2008, 140, 364–379. [Google Scholar] [CrossRef]
- Domrow, R. Bird nasal mites in New Zealand. Tuatara 1972, 19, 99–100. [Google Scholar]
- Shumilo, R.; Lunkashu, M. Rhinonyssid Mites from Southeastern URSS; Editorial Academy of Sciences of MSSR; Academy of Sciences of USSR: Moscow, Russia, 1970. [Google Scholar]
- Fain, A.; Herin, A.; Puylaert, F. Notes sur les Acariens nasicoles d’Oiseaux du Cameroun. Rev. Zool. Afr. 1977, 91, 385–396. [Google Scholar]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Found. Stat. Comput. Vienna Austria. 2020. Available online: http://www.R-project.org/ (accessed on 28 December 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 3319242776. [Google Scholar]
- Kassambara, A. ggpubr: “ggplot2” Based Publication Ready Plots. Available online: https://cran.r-project.org/package=ggpubr (accessed on 28 December 2020).
- Devleesschauwer, B.; Torgerson, P.; Charlier, J.; Levecke, B.; Praet, N.; Roelandt, S.; Smit, S.; Dorny, P.; Berkvens, D.; Speybroeck, N. Prevalence: Tools for prevalence assessment studies. R Packag. Version. 2015. Available online: http://cran.r-project.org/package=prevalence (accessed on 28 December 2020).
- Kohl, M. MKinfer: Inferential Statistics. R Packag. Version 0.5. 2020. Available online: http://cran.r-project.org/package=prevalence (accessed on 28 December 2020).
- Alec, F. Sur le statut de quelques espèces de Rhinonyssidae décrites par Hirst. Rev. Zool. Bot. Afr. 1960, 59, 310–314. [Google Scholar]
- Domrow, R. Some mite paraites of Australian birds. Proc. Linn. Soc. N. S. W. 1966, 90, 190–217. [Google Scholar]
- Fain, A.; Aitken, T.H.G. Les acariens parasites nasicoles des oiseaux de Trinidad (Indes Occidentales): I. Rhinonyssidae mesostigmates. Bull. Ann. Soc. R. D’Entomologie Belg. 1967, 43, 1–44. [Google Scholar]
- Sixl, W. Nasale Milben einheimischer Vögel. Mitteilungen Naturwissenschaftlichen Vereines Steiermark 1969, 99, 217–218. [Google Scholar]
- Strandtmann, R.W.; Wharton, G.W. A Manual of Mesostigmatid Mites Parasitic on Vertebrates; Institute of Acarology, University Maryland: College Park, MD, USA, 1958. [Google Scholar]
- Domrow, R. The nasal mites of Queensland birds (Acari: Dermanyssidae, Ereynetidae, and Epidermoptidae). Proc. Linnean Soc. N. S. W. 1969, 93, 297–426. [Google Scholar]
- Úbeda, J.M.; De Rojas, M.; Mora, M.D.; Guevara, D.C. Nasal mites (Mesostigmata, Rhinonyssidae) parasitic on columbiform birds from the Iberian Peninsula. Comparison of ribosomal ITS 1 region for species differentiation. Boletín la Real Soc. Española Hist. Nat. Sección Biológica 2018, 98, 65–75. [Google Scholar]
- Fain, A. Les acariens de la famille Rhinonyssidae Vitzthum 1935 parasites des fosses nasales des oiseaux au Ruanda-Urundi. Rev. Zool. Bot. Afri. 1956, 53, 131–157. [Google Scholar]
- Castro, M.P. de Reestruturação genérica da familia Rhinonyssidae Vitzthum, 1935 (Acari: Mesostigmata: Gamasides) e descrição de algumas espécies novas. Arq. Inst. Biol. (Sao Paulo) 1948, 18, 253–284. [Google Scholar]
- Pereira, C.; Castro, M.P. Revisão da subfamília Ptilonyssinae Castro, 1948 (Acari: Mesostigmata: Rhinonyssidae) com a descrição de algumas espécies novas. Arq. Inst. Biol (Sao. Paulo) 1949, 19, 218–235. [Google Scholar]
- Fain, A.; Sixl, W.; Moritsch, C. The nasal mites of the family Rhinonyssidae with description of a new species (Acarina). Mitteilungen der Abteilung Zool. Bot. Landesmuseum Joanneum Graz 1974, 3, 1–9. [Google Scholar]
- Feider, Z. Neonyssus hirsutus n.sp. (Rhinonyssidae, Parasitiformes), un nou parazit in cavitatea nazala la porumbelul domestic. Stidii. Cerc. Sti. Iasi. 1962, 13, 57–68. [Google Scholar]
- Crossley, D.A. A new species of nasal mite, Neonyssus (Neonyssus) columbae, from the pigeon (Acarina, Mesostigmata, Rhinonyssidae). Proc. Entomol. Soc. Wash 1950, 52, 309–312. [Google Scholar]
- De Rojas, M.; Doña, J.; Dimov, I. A comprehensive survey of Rhinonyssid mites (Mesostigmata: Rhinonyssidae) in Northwest Russia: New mite-host associations and prevalence data. Biodivers. Data J. 2020, 8, e49535. [Google Scholar] [CrossRef]
- Beron, P. Acarorum Catalogus VI. Order Mesostigmata. Gamasina: Dermanyssoidea (Rhinonyssidae, Spinturnicidae). In National Museum of Natural History, Sofia Bulgarian Academy of Sciences; Pensoft Publishers: Sofia, Bulgaria, 2020. [Google Scholar] [CrossRef]
- Spicer, G.S. Prevalence and host-parasite list of some nasal mites from birds (Acarina: Rhinonyssidae, Speleognathidae). J. Parasitol. 1987, 73, 259–264. [Google Scholar] [CrossRef]
- Doña, J.; Sweet, A.D.; Johnson, K.P.; Serrano, D.; Mironov, S.; Jovani, R. Cophylogenetic analyses reveal extensive host-shift speciation in a highly specialized and host-specific symbiont system. Mol. Phylogenet. Evol. 2017, 115, 190–196. [Google Scholar] [CrossRef]
- Crossley, D.A. Two new nasal mites from columbiform birds. J. Parasitol. 1952, 38, 385–390. [Google Scholar] [CrossRef]
- Rózsa, L. The ectoparasite fauna of feral pigeon populations in Hungary. Parasitol. Hungarica 1990, 23, 115–119. [Google Scholar]
- Foronda, P.; Valladares, B.; Rivera-Medina, J.A.; Figueruelo, E.; Abreu, N.; Casanova, J.C. Parasites of Columba livia (Aves: Columbiformes) in Tenerife (Canary Islands) and their role in the conservation biology of the laurel pigeons. Parasite 2004, 11, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, W.D. Extraordinary sex ratios. Science 1967, 156, 477–488. [Google Scholar] [CrossRef]
- Herre, E.A. Optimality, plasticity and selective regime in fig wasp sex ratios. Nature 1987, 329, 627–629. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Species | Parasite Species (n) | Number of Males | Number of Females | Number of Infested Hosts | Prevalence (IC95%) | Mean intensity±SE (Range) |
| Columba livia domestica | M. melloi (n = 1223) | 219 | 1004 | 85 | 0.34 (0.28–0.40) | 14.4 ± 2.39 (0–114) |
| M. columbae (n = 110) | 22 | 88 | 14 | 0.06 (0.03–0.09) | 7.86 ± 2.08 (0–25) | |
| Streptopelia decaocto | M. streptopeliae (n = 158) | 20 | 138 | 26 | 0.10 (0.07–0.15) | 6.08 ± 1.52 (0–40) |
| M. columbae (n = 3) | - | 3 | 1 | 0.004 (0.0001–0.22) | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veiga, J.; Dimov, I.; de Rojas, M. Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information. Diversity 2021, 13, 11. https://doi.org/10.3390/d13010011
Veiga J, Dimov I, de Rojas M. Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information. Diversity. 2021; 13(1):11. https://doi.org/10.3390/d13010011
Chicago/Turabian StyleVeiga, Jesús, Ivan Dimov, and Manuel de Rojas. 2021. "Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information" Diversity 13, no. 1: 11. https://doi.org/10.3390/d13010011
APA StyleVeiga, J., Dimov, I., & de Rojas, M. (2021). Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information. Diversity, 13(1), 11. https://doi.org/10.3390/d13010011