Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada


Abstract
:1. Introduction
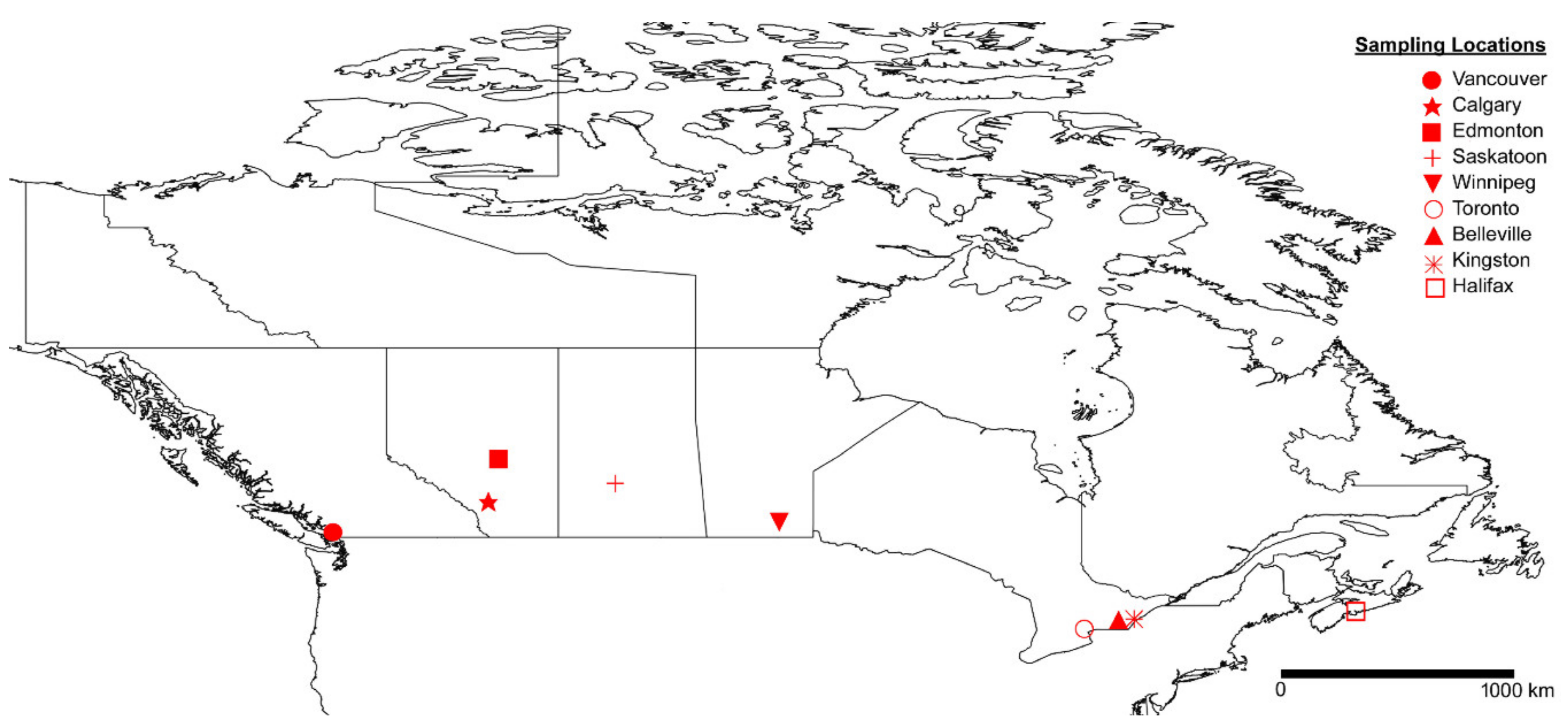
2. Materials and Methods
2.1. Collection and Identification of Ectosymbionts
2.2. Ectosymbiont Assemblages
2.3. Potential Effects of Local Climate on Ectosymbiont Assemblages
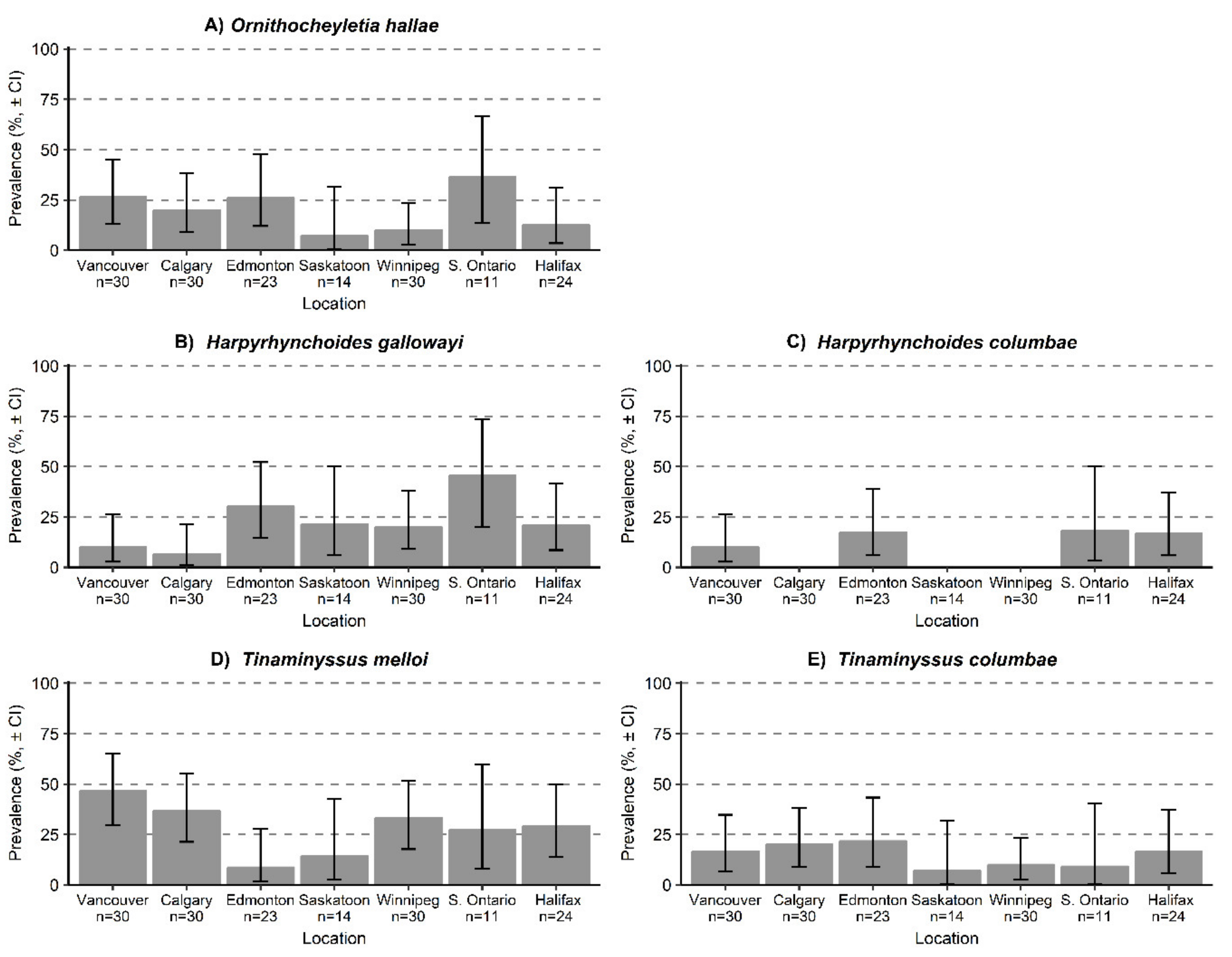
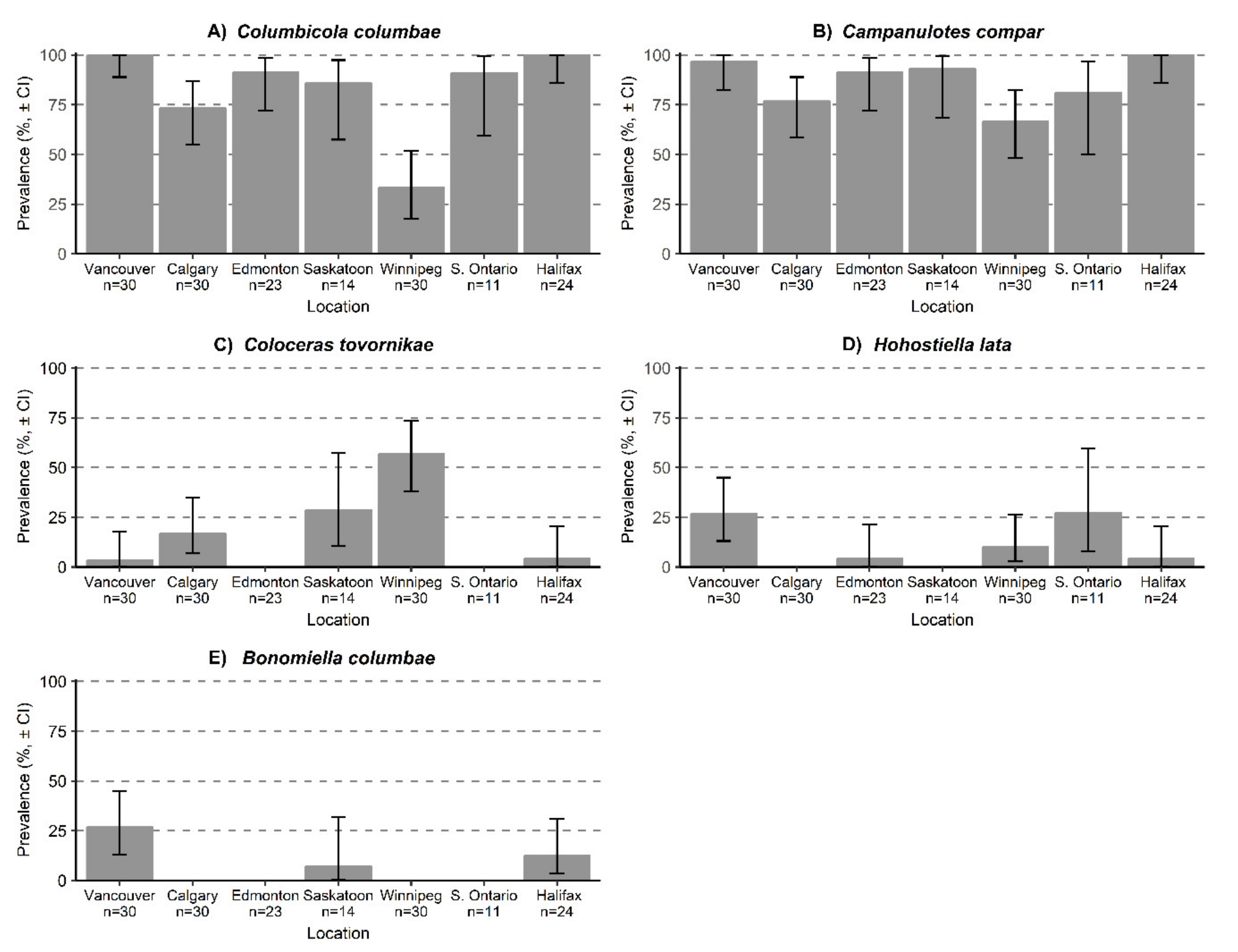
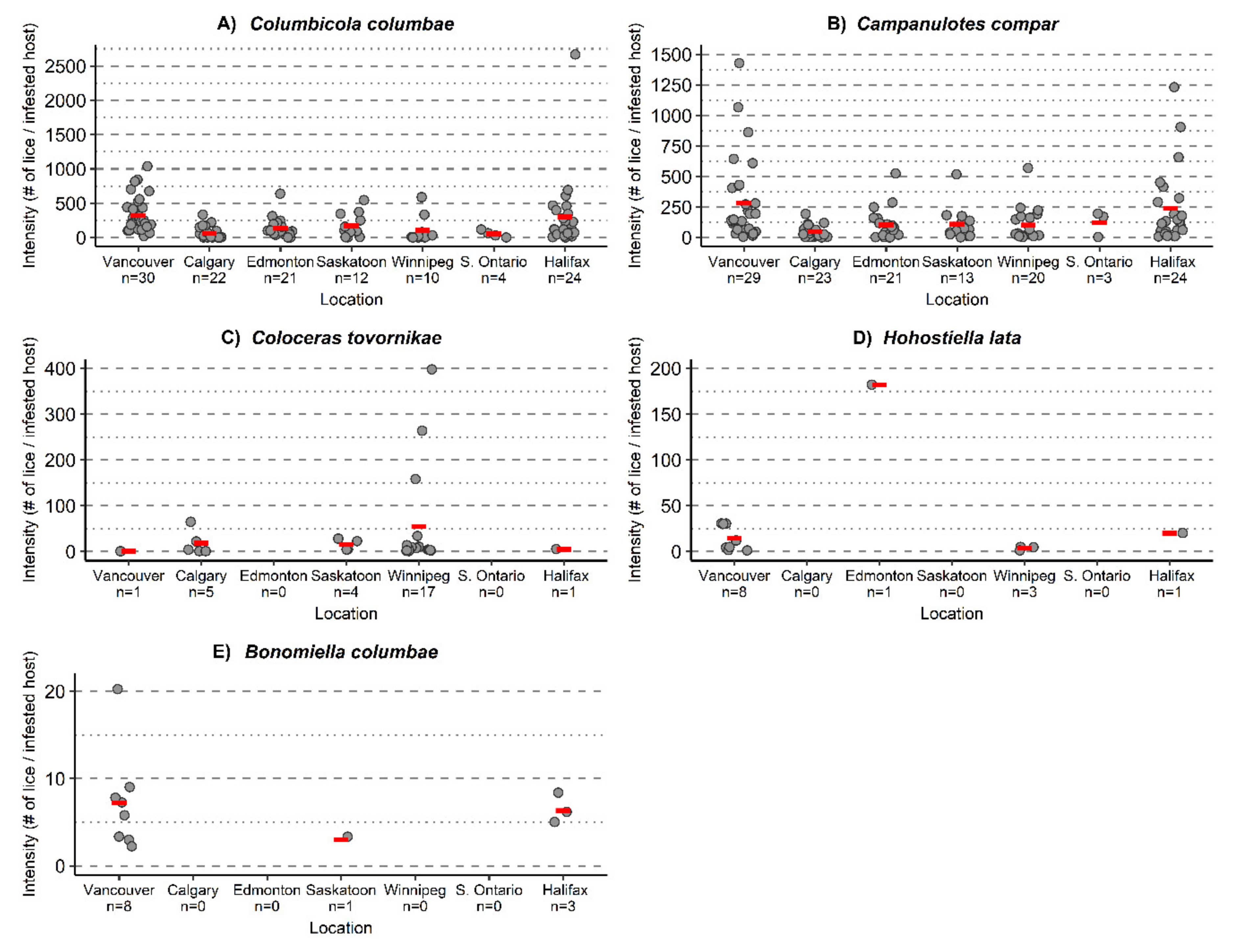
3. Results
3.1. Ectosymbiont Diversity, Prevalence, and Intensity
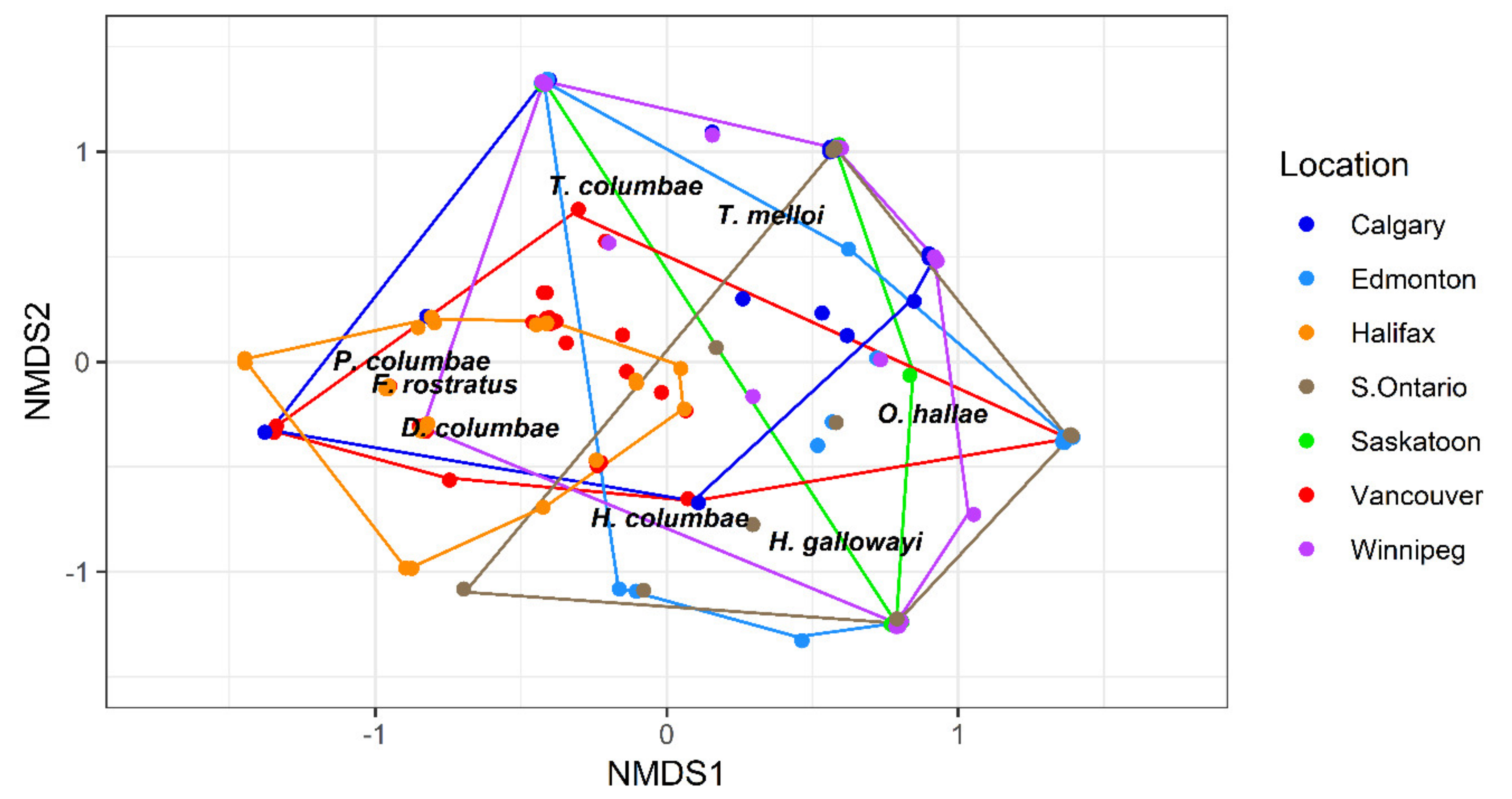
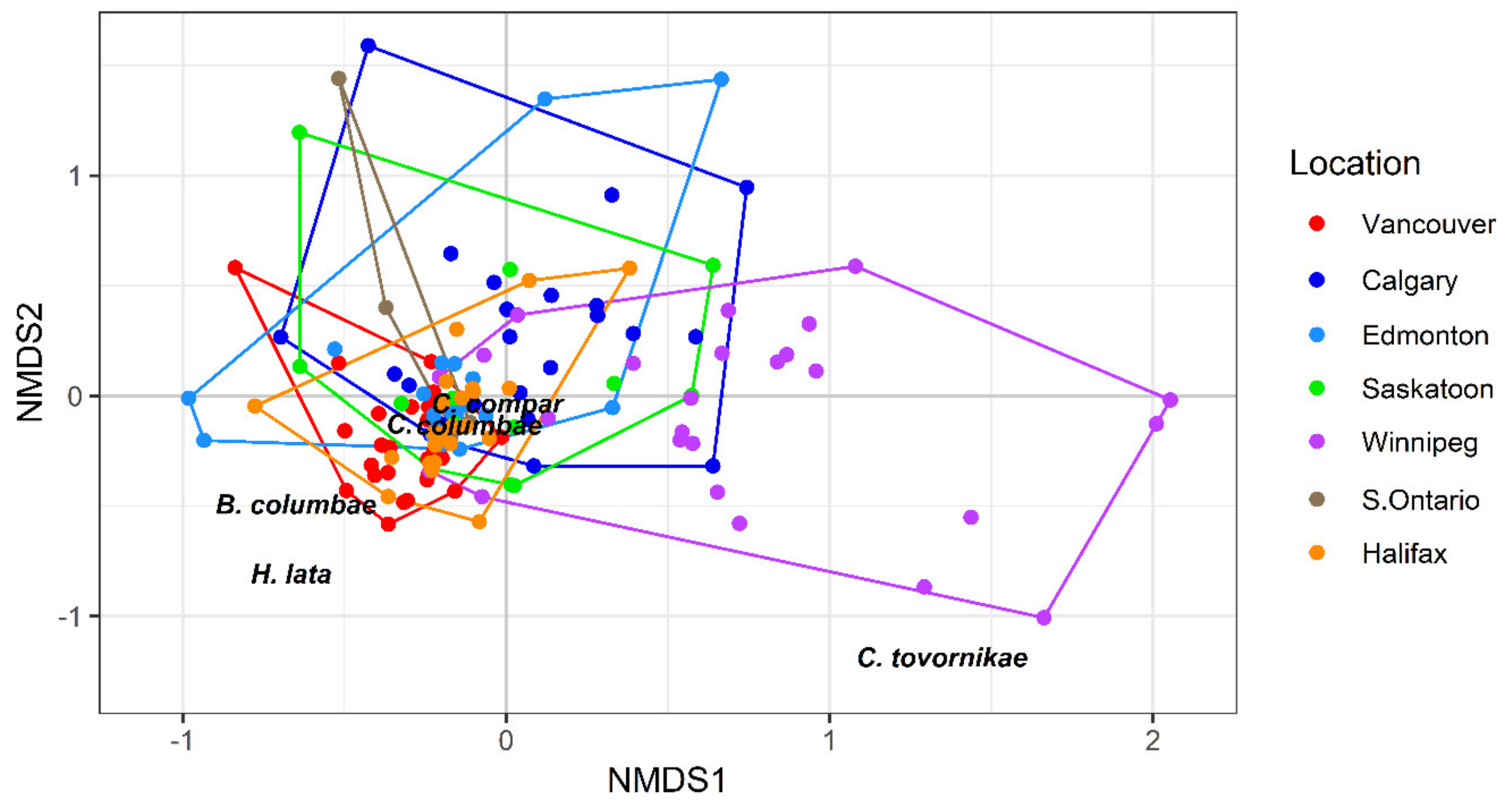
3.2. Assemblage Structure
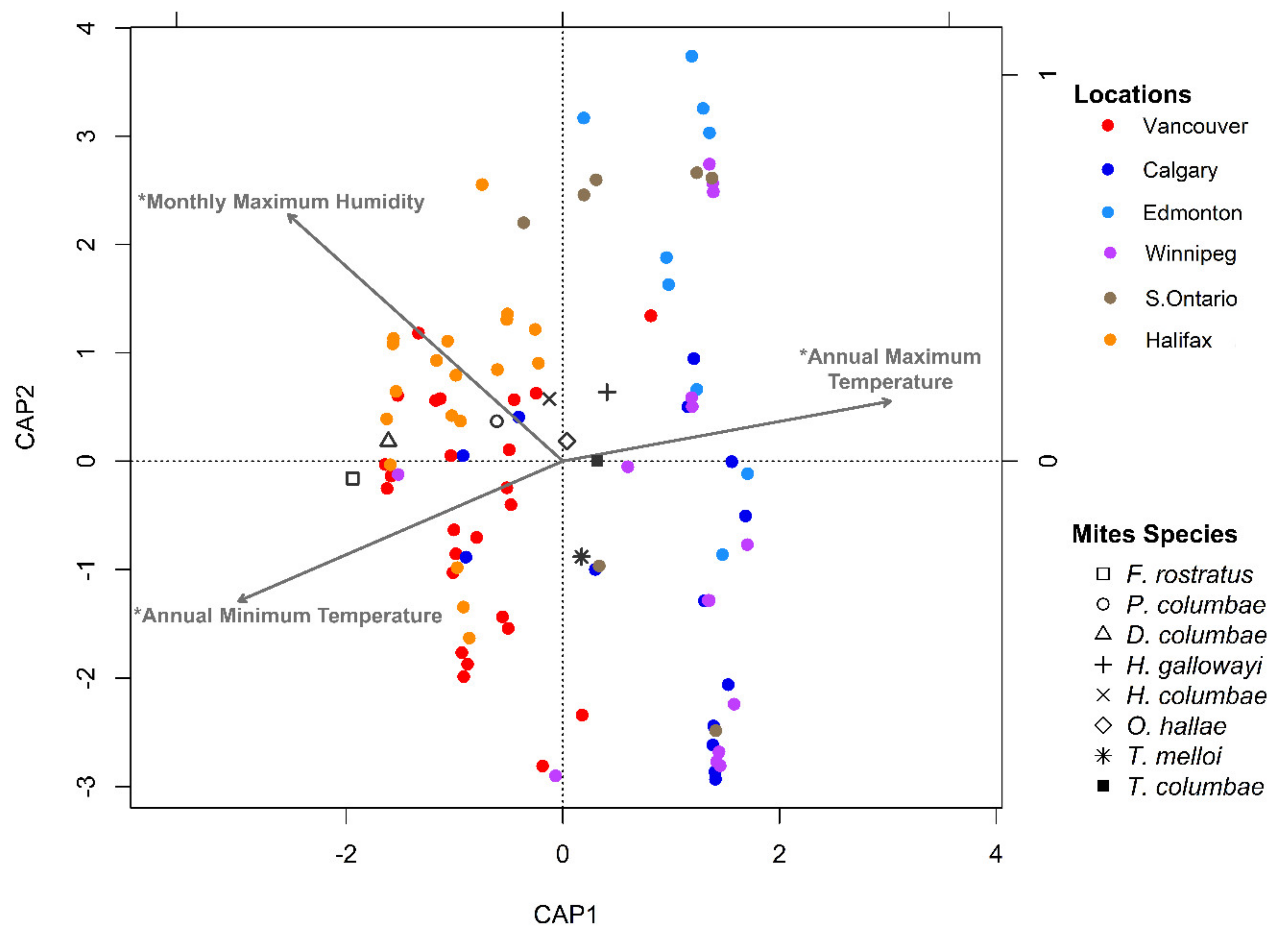
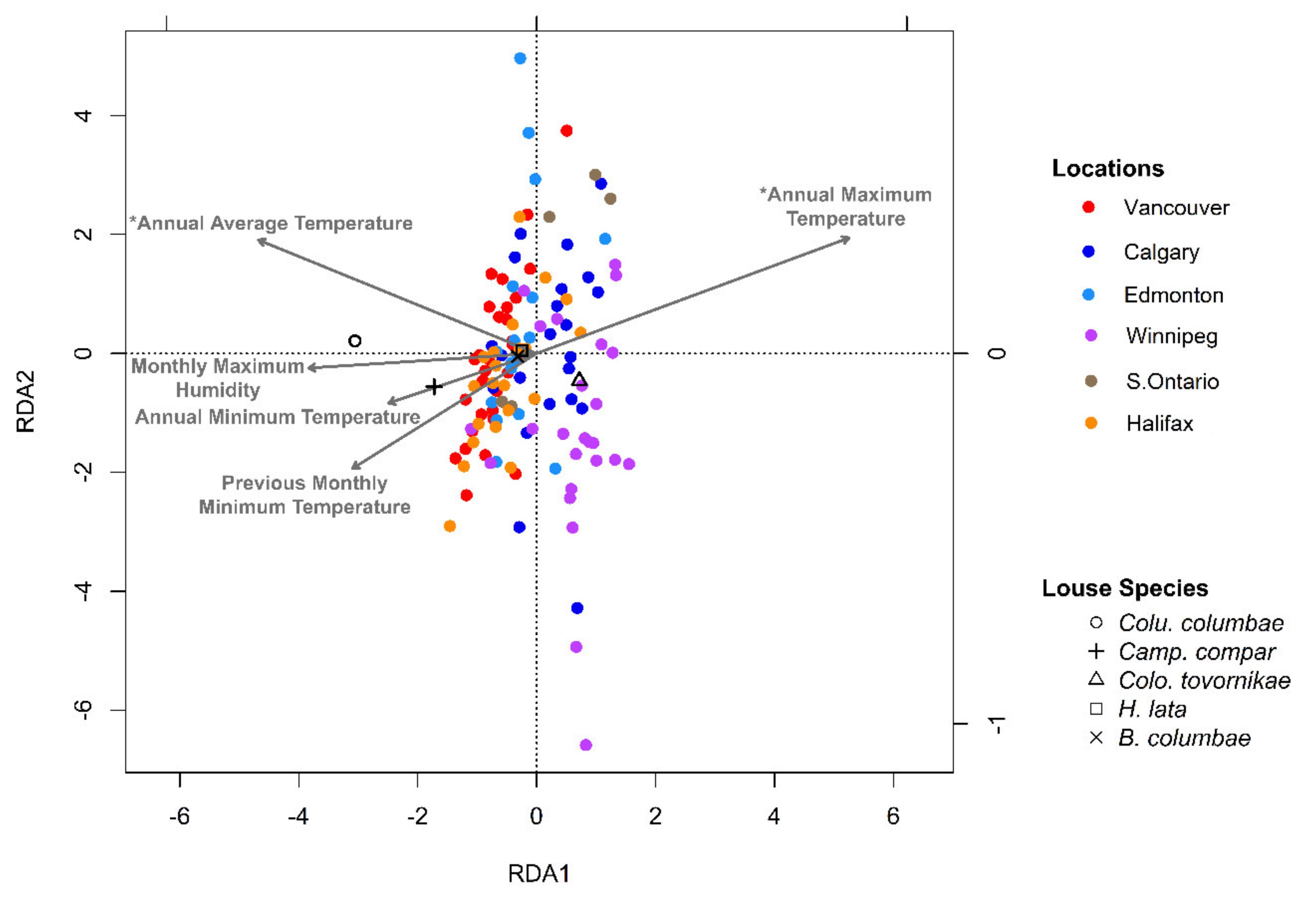
3.3. Potential Effects of Local Climate on Ectosymbiont Assemblages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torchin, M.E.; Mitchell, C.E. Parasites, pathogens, and invasions by plants and animals. Front. Ecol. Environ. 2004, 2, 183–190. [Google Scholar] [CrossRef]
- Lymbery, A.J.; Morine, M.; Kanani, H.G.; Beatty, S.J.; Morgan, D.L. Co-invaders: The effects of alien parasites on native hosts. Int. J. Parasitol. Parasites Wildl. 2014, 3, 171–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheath, D.J.; Williams, C.F.; Reading, A.J.; Robert Britton, J. Parasites of non-native freshwater fishes introduced into England and Wales suggest enemy release and parasite acquisition. Biol. Invasions 2015, 17, 2235–2246. [Google Scholar] [CrossRef] [Green Version]
- Macleod, C.J.; Paterson, A.M.; Tompkins, D.M.; Duncan, R.P. Parasites lost-do invaders miss the boat or drown on arrival? Ecol. Lett. 2010, 13, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.M.; Palma, R.L.; Gray, R.D. Drowning on arrival, missing the boat, and x-events: How likely are sorting events? In Tangled Trees: Phylogeny, Cospeciation and Coevolution; Page, R.D., Ed.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 287–309. [Google Scholar]
- Torchin, M.E.; Lafferty, K.D.; Dobson, A.P.; Mckenzie, V.J.; Kuris, A.M. Introduced species and their missing parasites. Lett. Nat. 2003, 421, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R. Evolutionary Ecology of Parasites, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Dabert, J. Feather mites (Astigmata; Pterolichoidea, Analgoidea) and birds as models for cophylogenetic studies. Phytophaga 2004, 14, 409–424. [Google Scholar]
- Dabert, J.; Dabert, M.; Mironov, S.V. Phylogeny of feather mite subfamily avenzoariinae (Acari: Analgoidea: Avenzoariidae) inferred from combined analyses of molecular and morphological data. Mol. Phylogenet. Evol. 2001, 20, 124–135. [Google Scholar] [CrossRef]
- Dabert, J. The feather mite family Syringobiidae Trouessart, 1896 (Acari, Astigmata, Pterolichoidea). 2. Phylogeny and host-parasite evolutionary relationships. Acta Parasitol. 2003, 48, 185–233. [Google Scholar]
- Banks, J.C.; Palma, R.L.; Paterson, A.M. Cophylogenetic relationships between penguins and their chewing lice. J. Evol. Biol. 2006, 19, 156–166. [Google Scholar] [CrossRef]
- Johnson, K.P.; Kennedy, M.; McCracken, K.G. Reinterpreting the origins of flamingo lice: Cospeciation or host-switching? Biol. Lett. 2006, 2, 275–278. [Google Scholar] [CrossRef]
- Gustafsson, D.R.; Zou, F. Gallancyra gen. nov. (Phthiraptera: Ischnocera), with an overview of the geographical distribution of chewing lice parasitizing chicken. Eur. J. Taxon. 2020, 685, 1–36. [Google Scholar] [CrossRef]
- Price, R.D.; Hellenthal, R.A.; Palma, R.L.; Johnson, K.P.; Clayton, D.H. The Chewing Lice: World Checklist and Biological Overview; Illinois Natural History Survey Special Publication: Champaign, IL, USA, 2003. [Google Scholar]
- Gaud, J.; Atyeo, W.T. Feather mites of the world (Acarina, Astigmata): The supraspecific taxa. Mus. R. Afr. Cent. Ann. Sci. Zool. 1996, 277, 1–193. [Google Scholar]
- Marshall, A. The Ecology Ectoparasitic Insects; Academic Press: London, UK, 1981. [Google Scholar]
- Proctor, H.C. Feather mites (Acari: Astigmata): Ecology, behavior, and evolution. Annu. Rev. Entomol. 2003, 48, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha Amaral, H.L.; Bergmann, F.B.; Silveira, T.; dos Santos, P.R.S.; Krüger, R.F. Pseudolynchia canariensis (Diptera: Hippoboscidae): Distribution pattern and phoretic association with skin mites and chewing lice of Columba livia (Aves: Columbidae). J. Nat. Hist. 2013, 47, 2927–2936. [Google Scholar] [CrossRef]
- Goodwin, D. Pigeons and Doves of the World, 2nd ed.; British Museum (Natural History): London, UK, 1977. [Google Scholar]
- EBird eBird: An online database of bird distribution and abundance (web application). eBird, Cornell Lab of Ornithology: Ithaca, NY, USA. Available online: http://www.ebird.org. (accessed on 10 December 2019).
- Schorger, A.W. Introduction of the domestic pigeon. Auk 1952, 69, 462–463. [Google Scholar] [CrossRef]
- Galloway, T.D.; Lamb, R.J. Infestation parameters for chewing lice (Phthiraptera: Amblycera, Ischnocera) infesting owls (Aves: Strigidae, Tytonidae) in Manitoba, Canada. Can. Entomol. 2019, 151, 608–620. [Google Scholar] [CrossRef]
- Lamb, R.J.; Galloway, T.D. Abundance and stability of populations of chewing lice (Phthiraptera: Amblycera and Ischnocera) infesting two species of woodpeckers (Aves: Piciformes: Picidae). Can. Entomol. 2018, 150, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Galloway, T.D. Phthiraptera of Canada. Zookeys 2019, 819, 301–310. [Google Scholar] [CrossRef]
- Knee, W.; Proctor, H.; Galloway, T. Survey of nasal mites (Rhinonyssidae, Ereynetidae, and Turbinoptidae) associated with birds in Alberta and Manitoba, Canada. Can. Entomol. 2008, 140, 364–379. [Google Scholar] [CrossRef]
- Mitchell, W.R. Observations on poultry diseases in Ontario. Ontario Vet. Coll. Rep. 1953, 1952, 78–79. [Google Scholar]
- Moyer, B.R.; Drown, D.M.; Clayton, D.H. Low humidity reduces ectoparasite pressure: Implications for host life history evolution. Oikos 2002, 97, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, C.M.; Valera, F.; Barbosa, A.; Moreno, E. Thriving in an arid environment: High prevalence of avian lice in low humidity conditions. Écoscience 2007, 12, 241–249. [Google Scholar] [CrossRef]
- Dowling, D.K.; Richardson, D.S.; Blaakmeer, K.; Komdeur, J. Feather mite loads influenced by salt exposure, age and reproductive stage in the Seychelles Warbler Acrocephalus sechellensis. J. Avian Biol. 2001, 32, 364–369. [Google Scholar] [CrossRef]
- Meléndez, L.; Laiolo, P.; Mironov, S.; García, M.; Magaña, O.; Jovani, R. Climate-driven variation in the intensity of a host-symbiont animal interaction along a broad elevation gradient. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabiyi, T.P. Association between duration of humid season and geographical distribution patterns of different species of chewing lice (Mallophaga: Insecta) infesting domestic chickens in Nigeria. J. Parasitol. 1996, 82, 1034–1036. [Google Scholar] [CrossRef]
- Shorthouse, D.P. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. Available online: www.simplemappr.net (accessed on 25 September 2020).
- Grossi, A.A.; Sharanowski, B.J.; Galloway, T.D. Anatoecus species (Phthiraptera: Philopteridae) from Anseriformes in North America and taxonomic status of Anatoecus dentatus and Anatoecus icterodes. Can. Entomol. 2014, 146, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Petryszak, A.; Rościszewska, M.; Bonczar, Z.; Szwalec, R. Observations on the vitality of lice from dead pigeons. Wiad. Parazytol. 1996, 42, 337–347. (In Polish) [Google Scholar]
- Gaud, J. Acariens Sarcoptiformes plumicoles parasites sur les oiseaux Lariformes et Columbiformes d’Afrique. Ann. Mus. R. Afr. Cent. Sér. 8 Sci. Zool. 1976, 214, 1–101. (In French) [Google Scholar]
- Smiley, R.L. A review of the family Cheyletiellidae (Acarina). Ann. Entomol. Soc. Am. 1970, 63, 1056–1078. [Google Scholar] [CrossRef]
- Bochkov, A.V.; Galloway, T.D. Parasitic cheyletoid mites (Acari: Cheyletoidea) associated with passeriform birds (Aves: Passeriformes) in Canada. Can. J. Zool. 2001, 79, 2014–2028. [Google Scholar] [CrossRef]
- Emerson, K.C. A new species of Mallophaga from the Pigeon. Florida Entomol. 1957, 40, 63–64. [Google Scholar] [CrossRef]
- Galloway, T.D.; Palma, R.L. Serendipity with chewing lice (Phthiraptera: Menoponidae, Philopteridae) infesting rock pigeons and mourning doves (Aves: Columbiformes: Columbidae) in Manitoba, with new records for North America and Canada. Can. Entomol. 2008, 140, 208–218. [Google Scholar] [CrossRef]
- Adams, R.J.; Price, R.D.; Clayton, D.H. Taxonomic revision of Old World members of the feather louse genus Columbicola (Phthiraptera: Ischnocera), including descriptions of eight new species. J. Nat. Hist. 2005, 39, 3545–3618. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own term: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Peter, S.; et al. Vegan: Community Ecology Package. R package v 2.4–6 2018. Available online: https://cran.ism.ac.jp/web/packages/vegan/vegan.pdf (accessed on 1 September 2020).
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/. (accessed on 1 September 2020).
- Bland, M.; Altman, D.G. Multiple significance tests: The Bonferroni method. BMJ 1995, 310, 170. [Google Scholar] [CrossRef] [Green Version]
- Simon, S. A Modern Approach to Regression with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Ròzsa, L. Speciation patterns of ectoparasites and “straggling” lice. Int. J. Parasitol. 1993, 23, 859–864. [Google Scholar] [CrossRef]
- Kerness Whiteman, N.; Santiago-Alarcon, D.; Johnson, K.P.; Parker, P.G. Differences in straggling rates between two genera of dove lice (Insecta: Phthiraptera) reinforce population genetic and cophylogenetic patterns. Int. J. Parasitol. 2004, 34, 1113–1119. [Google Scholar] [CrossRef]
- Rayner, J.A. Parasites of wild birds in Quebec. Sci. Agric. 1932, 12, 307–309. [Google Scholar] [CrossRef]
- Bochkov, A.V.; O’Connor, B.M.; Klompen, H. A review of the mite subfamily Harpirhynchinae (Acariformes: Harpirhynchidae)—Parasites of New World birds (Aves: Neognathae). Zootaxa 2015, 4023, 1–130. [Google Scholar] [CrossRef] [Green Version]
- Banks, N. New Canadian mites. Proc. Entomol. Soc. Washingt. 1909, 11, 133–143. [Google Scholar]
- Hill, W.W.; Tuff, D.W. A review of the Mallophaga parasitizing the Columbiformes of North America North of Mexico. J. Kans. Entomol. Soc. 1978, 51, 307–327. [Google Scholar]
- Harbison, C.W.; Bush, S.E.; Malenke, J.R.; Clayton, D.H. Comparative transmission dynamics of competing parasite species. Ecology 2008, 89, 3186–3194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaud, J.; Petitot, M.L. Sacroptides plumicoles des oiseaux du Maroc. Ann. Parasitol. Hum. Comparée 1948, 23, 35–46. (In French) [Google Scholar] [CrossRef]
- Millthorpe, A.P.; Eves, N. Pigeon mites and human infestation. New Zeal. Entomol. 1971, 5, 90–92. [Google Scholar] [CrossRef]
- Oudemans, A.C. Description du Myialges anchora Sergent et Trouessart 1907 (Acarien). Ann. Parasitol. Hum. Comp. 1935, 13, 5–11. [Google Scholar] [CrossRef]
- Dubinin, V. Studies in the adaptation of ectoparasites. 2. Ecological adaptations of the mallophagans and feather mites. Parazitol. Sb. 1947, 9, 191–222. (In Russian) [Google Scholar]
- Gaud, J.; Barré, N. Les genres Pterophagoides et Byersalges (Falculiferidae, Pterolichoidea) parasites plumicoles des Columbidae. Acarologia 1988, 29, 63–71. (In French) [Google Scholar]
- Dubinin, V. Fauna of the USSR—Arachnida. Part 3. Family Pterolichidae 1. Zool. Inst. Acad. Sci. USSR 1956, 6, 1–363. (In Russian) [Google Scholar]
- Cerny, V. Die parasitischen Arthropoden der synanthropen Taubenvögel einer Grossstadt. Angew. Parasitol. 1970, 11, 244–248. [Google Scholar]
- Naz, S.; Sychra, O.; Rizvi, S.A. New records and a new species of chewing lice (Phthiraptera, Amblycera, Ischnocera) found on Columbidae (Columbiformes) in Pakistan. Zookeys 2012, 174, 79–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knab, F. Four European Diptera established in North America. Insecutor Inscitiae Menstruus 1916, 4, 1–4. [Google Scholar]
- Martin, M. Life history and habits of the Pigeon Louse (Colimbicola columbae L.). Can. Entomol. 1934, 66, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Clayton, D.H.; Moyer, B.R.; Bush, S.E.; Jones, T.G.; Gardiner, D.W.; Rhodes, B.B.; Goller, F. Adaptive significance of avian beak morphology for ectoparasite control. Proc. Biol. Sci. 2005, 272, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doña, J.; Proctor, H.; Serrano, D.; Johnson, K.P.; Oploo, A.O.; Huguet-Tapia, J.C.; Ascunce, M.S.; Jovani, R. Feather mites play a role in cleaning host feathers: New insights from DNA metabarcoding and microscopy. Mol. Ecol. 2019, 28, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.C.; Murray, M.D. The distribution of Mallophaga on the domestic pigeon (Columba livia). Int. J. Parasitol. 1971, 1, 21–29. [Google Scholar] [CrossRef]
- Galloway, T.D.; Lamb, R.J. Seasonal population dynamics of four species of chewing lice (Phthiraptera: Menoponidae, Philopteridae) on feral pigeons (Aves: Columbiformes: Columbidae). Can. Entomol. 2015, 147, 712–722. [Google Scholar] [CrossRef] [Green Version]
- Pap, P.L.; Vágási, C.I.; Osváth, G.; Mureşan, C.; Barta, Z. Seasonality in the uropygial gland size and feather mite abundance in house sparrows Passer domesticus: Natural covariation and an experiment. J. Avian Biol. 2010, 41, 653–661. [Google Scholar] [CrossRef]
- Blanco, G.; Frías, O. Symbiotic feather mites synchronize dispersal and population growth with host sociality and migratory disposition. Ecography 2001, 24, 113–120. [Google Scholar] [CrossRef]
- Hamstra, T.L.; Badyaev, A.V. Comprehensive investigation of ectoparasite community and abundance across life history stages of avian host. J. Zool. 2009, 278, 91–99. [Google Scholar] [CrossRef]
- Brown, N.S. A survey of the arthropod parasites of pigeons (Columba livia) in Boston. J. Parasitol. 1971, 57, 1379–1380. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Anser Rizvi, S.; Sychra, O. The high rate of infestation of chewing lice (Phthiraptera) in Rock Pigeons (Columba livia Gmelin 1789) in Pakistan. Trop. Zool. 2010, 23, 21–28. [Google Scholar]
- Forond, A.P.; Valladares, B.; Rivera-Medin, A.J.A.; Figueruel, O.E.; Abre, U.N.; Casanov, A.J.C. Parasites of Columba livia (Aves: Columbiformes) in Tenerife (Canary Islands) and their role in the conservation biology of the Laurel pigeons. Parasite 2004, 11, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adang, K.L.; Oniye, S.J.; Ezealor, A.U.; Abdu, P.A.; Ajanusi, O.J. Ectoparasites of domestic pigeon (Columba livia domestica, Linnaeus) in Zaria, Nigeria. Res. J. Parasitol. 2008, 3, 79–84. [Google Scholar] [CrossRef]
- Bochkov, A.; Williams, G.; Proctor, H. First record of Picobia zumpti (Acari: Syringophilidae) from quills of the rock pigeon in North America and description of the male. Belg. J. Entomol. 2006, 7, 121–127. [Google Scholar]
- Kaszewska, K.; Skoracki, M.; Hromada, M. A review of the quill mites of the genus Gunabopicobia Skoracki and Hromada (Acariformes: Prostigmata: Syringophilidae) associated with birds of the order Columbiformes. Int. J. Acarol. 2018, 44, 288–299. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vancouver (n = 30) | Calgary (n = 30) | Edmonton (n = 23) | Saskatoon (n = 14) | Winnipeg (n = 30) | S. Ontario (n = 11) | Halifax (n = 24) | |
|---|---|---|---|---|---|---|---|
| Falculiferidae | |||||||
| Falculifer rostratus | 93.3 (78.7–98.8) | 10.0 (2.8–26.3) | 0 | 0 | 6.7 (1.2–21.3) | 9.1 (0.5–40.4) | 83.3 (62.8–94.1) |
| Pterophagus columbae | 13.3 (4.7–29.8) | 0 | 0 | 0 | 0 | 0 | 41.7 (23.4–62.8) |
| Analgidae | |||||||
| Diplaegidia columbae | 76.7 (58.4–88.8) | 6.7 (1.2–21.3) | 8.7 (1.6–27.8) | 0 | 6.7 (1.2–21.3) | 27.3 (7.9–59.6) | 83.3 (62.8–94.1) |
| Harpirhynchidae | |||||||
| Harpyrhynchoides gallowayi | 10.0 (2.8–26.3) | 6.7 (1.2–21.3) | 30.4 (14.5–52.2) | 21.4 (6.1–50.0) | 20.0 (9–38) | 45.5 (20.0–73.5) | 20.8 (8.6–41.5) |
| Harpyrhynchoides columbae | 10.0 (2.8–26.3) | 0 | 17.3 (6.2–38.9) | 0 | 0 | 8.2 (3.3–50.0) | 16.7 (5.9–37.2) |
| Cheyletidae | |||||||
| Ornithocheyletia hallae | 26.7 (13.1–44.9) | 20.0 (9.1–38.2) | 26.1 (12.0–47.8) | 7.1 (0.4–31.7) | 10.0 (2.8–23.6) | 36.4 (13.5–66.7) | 12.5 (3.5–31.0) |
| Rhinonyssidae | |||||||
| Tinaminyssus melloi | 46.7 (29.8–65.2) | 36.7 21.4–55.1) | 8.7 (1.6–27.8) | 14.2 (2.6–42.6) | 33.3 (17.7–51.7) | 27.3 (7.9–59.6) | 29.2 (13.9–50.0) |
| Tinaminyssus columbae | 16.7 (6.8–34.7) | 20.0 (9.1–38.2) | 21.7 (9.0–43.3) | 7.1 (0.4–31.7) | 10.0 (2.8–23.4) | 9.1 (0.5–40.4) | 16.7 (5.9–37.2) |
| Vancouver (n = 30) | Calgary (n = 30) | Edmonton (n = 23) | Saskatoon (n = 14) | Winnipeg (n = 30) | S. Ontario (n = 11) | Halifax (n = 24) | |
|---|---|---|---|---|---|---|---|
| Philopteridae | |||||||
| Columbicola columbae | 100 (88.9–100) | 73.3 (55.1–86.9) | 91.3 (72.2–98.4) | 85.7 (57.4–97.4) | 33.3 (17.7–51.7) | 90.9 (59.6–99.5) | 100 (86.1–100) |
| Campanulotes compar | 96.7 (82.3–99.8) | 76.7 (58.4–88.8) | 91.3 (72.2–98.4) | 92.9 (68.3–99.6) | 66.7 (48.3–82.3) | 81.1 (50.0–96.7) | 100.0 (86.1–100) |
| Coloceras tovornikae | 3.3 (0.2–17.7) | 16.7 (6.8–34.8) | 0 | 28.6 (10.4–57.4) | 56.7 (38.2–73.7) | 0 | 4.2 (0.2–20.4) |
| Menoponidae | |||||||
| Hohorstiella lata | 26.7 (13.1–44.9) | 0 | 4.3 (0.2–21.3) | 0 | 10.0 (2.8–26.3) | 27.3 (7.9–59.6) | 4.2 (0.2–20.4) |
| Bonomiella columbae | 26.7 (13.1–44.9) | 0 | 0 | 7.1 (0.4–31.7) | 0 | 0 | 12.5 (3–31.0) |
| Vancouver | Calgary | Edmonton | Saskatoon | Winnipeg | S. Ontario | Halifax | |
|---|---|---|---|---|---|---|---|
| Philopteridae | |||||||
|
Columbicola columbae | 326.1 (246.7–427.7) [22–841] | 64.2 (33.9–112.0) [1–2026] | 136.2 (95.1–219.1) [1–640] | 173.6 (97.5–287.7) [4–542] | 107.5 (20.3–280.6) [1–9] | 53.8 (16.5–96.3) [2–118] | 303.1 (171.0–696.3) [1–2669] |
|
Campanulotes compar | 282.0 (186.0–441.6) [6–1428] | 47.8 (32.2–71.9) [1–194] | 104.8 (67.3–175.1) [1–524] | 110.1 (66.5–199.2) ^ [2–318] | 102.8 (61.7–183.6)[4–568] | 122.7× [4–195] | 239.2 (148.5–400.3) [1–2669] |
| Coloceras tovornikae | 1.0 * | 18.6 (2.2–44.2) [1–65] | - | 15.0 (4.3–25.5) [4–28] | 54.2 (22.3–117.6) ^ [1–404] | - | 5.0 * |
| Menoponidae | |||||||
| Hohorstiella lata | 14.5 (6.4–23.8) [1–31] | - | 182.0 * | - | 3.7 × [1–5] | - | 20.0 * |
|
Bonomiella columbae | 7.3 (4.5–12.4) [2–20] | - | - | 3.0 * | - | - | 6.3 (5.0–7.3) [5–8] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grossi, A.; Proctor, H. Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada. Diversity 2021, 13, 9. https://doi.org/10.3390/d13010009
Grossi A, Proctor H. Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada. Diversity. 2021; 13(1):9. https://doi.org/10.3390/d13010009
Chicago/Turabian StyleGrossi, Alexandra, and Heather Proctor. 2021. "Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada" Diversity 13, no. 1: 9. https://doi.org/10.3390/d13010009
APA StyleGrossi, A., & Proctor, H. (2021). Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada. Diversity, 13(1), 9. https://doi.org/10.3390/d13010009