On the Challenge to Correctly Identify Rasboras (Teleostei: Cyprinidae: Danioninae) Inhabiting the Mesangat Wetlands, East Kalimantan, Indonesia

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Design
2.2. Material Examined
2.3. Classical Species Determination
2.4. DNA Barcoding
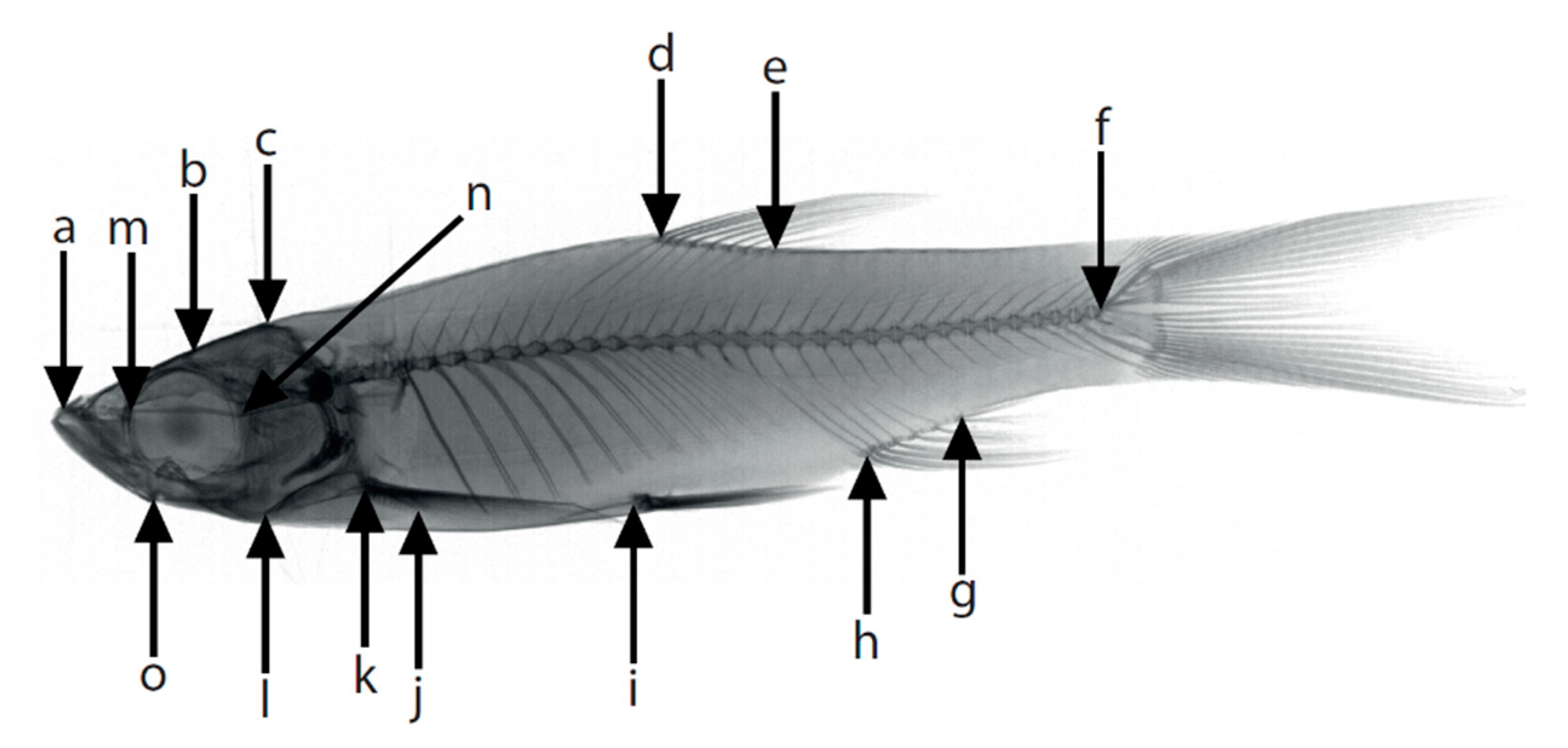
2.5. Morphometric Analysis
3. Results
3.1. Classical Species Determination
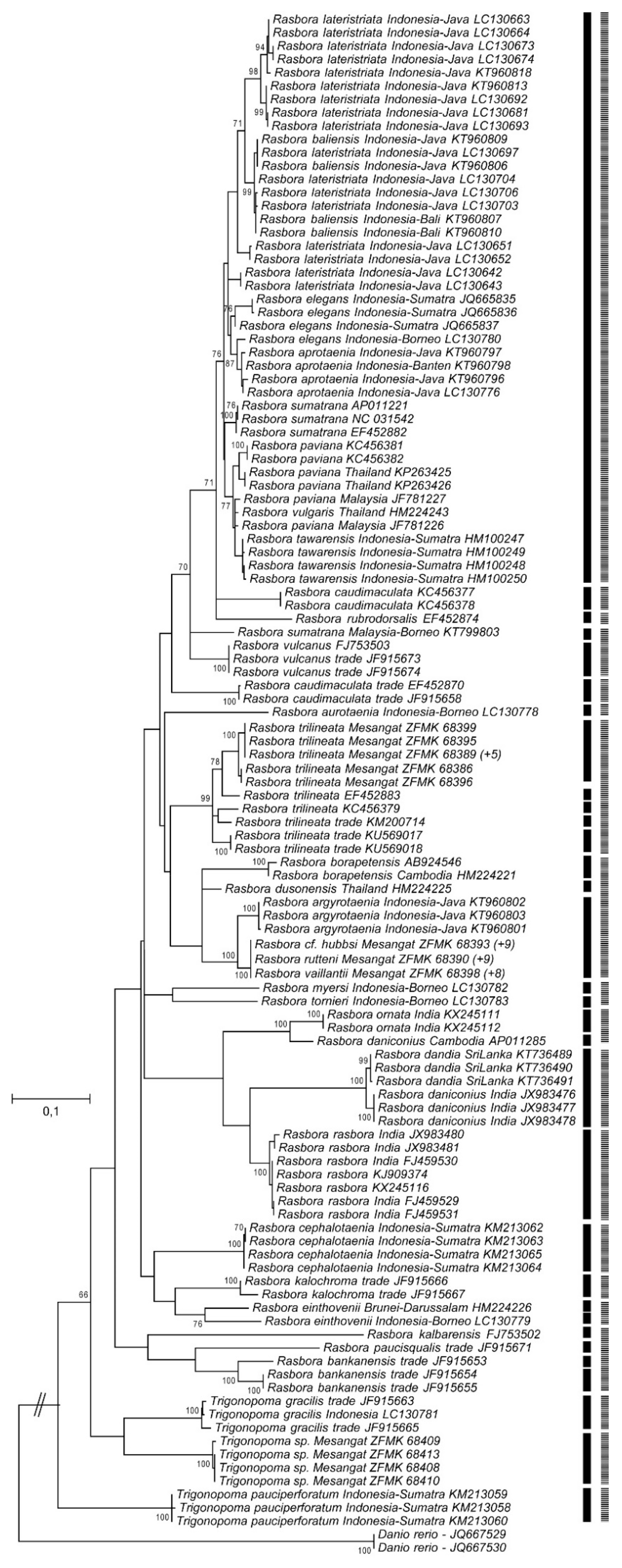
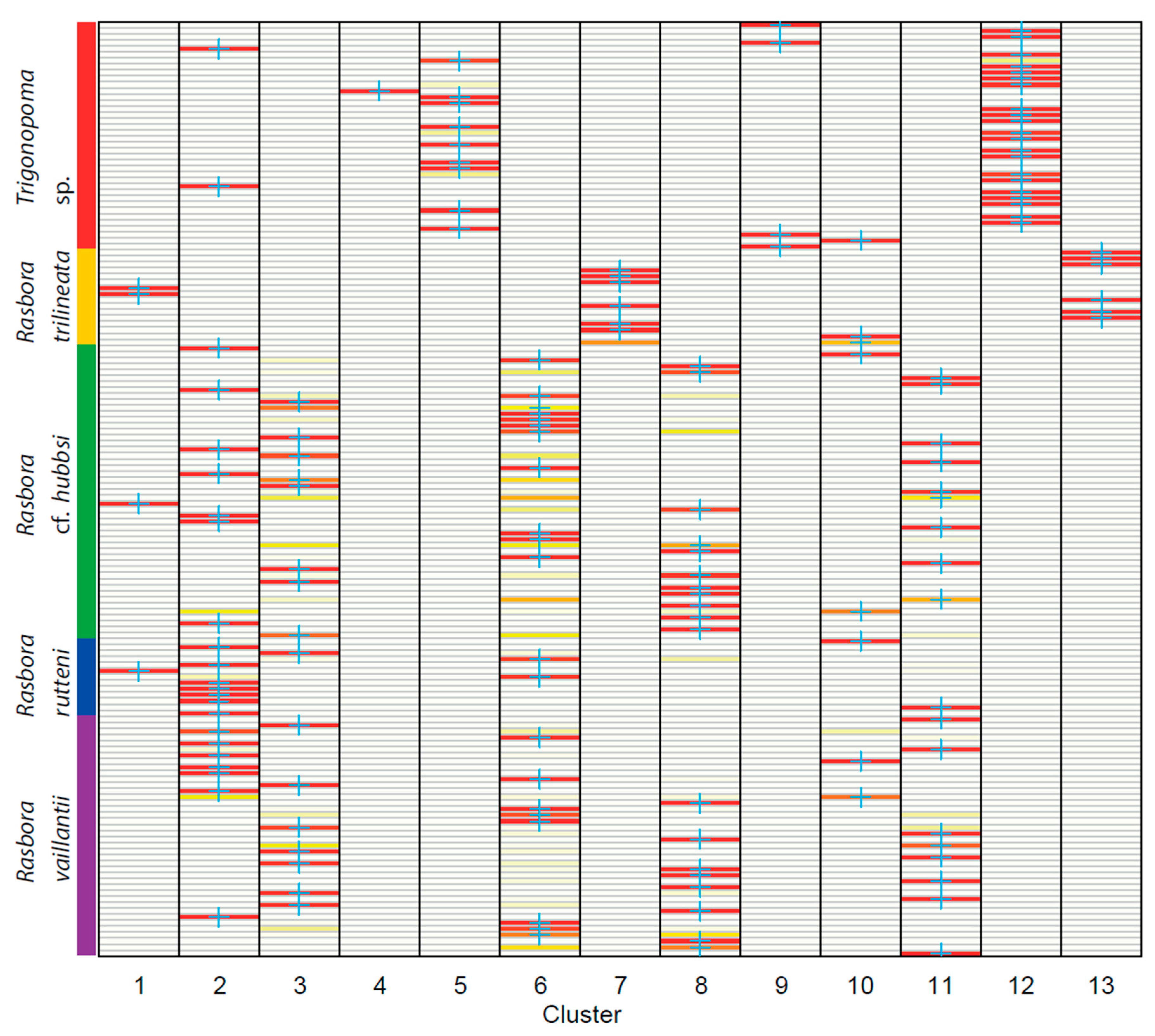
3.2. DNA Barcoding
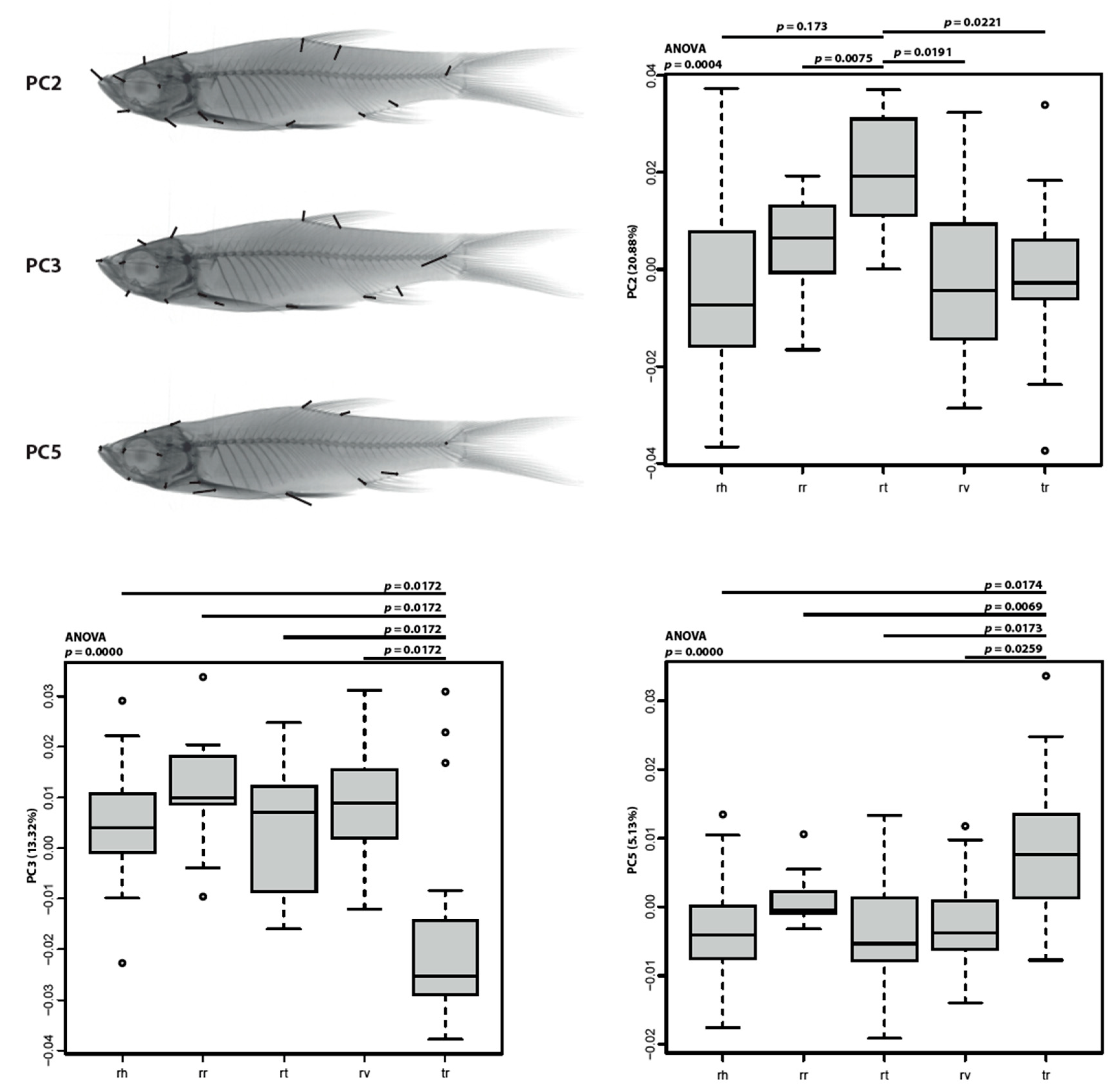
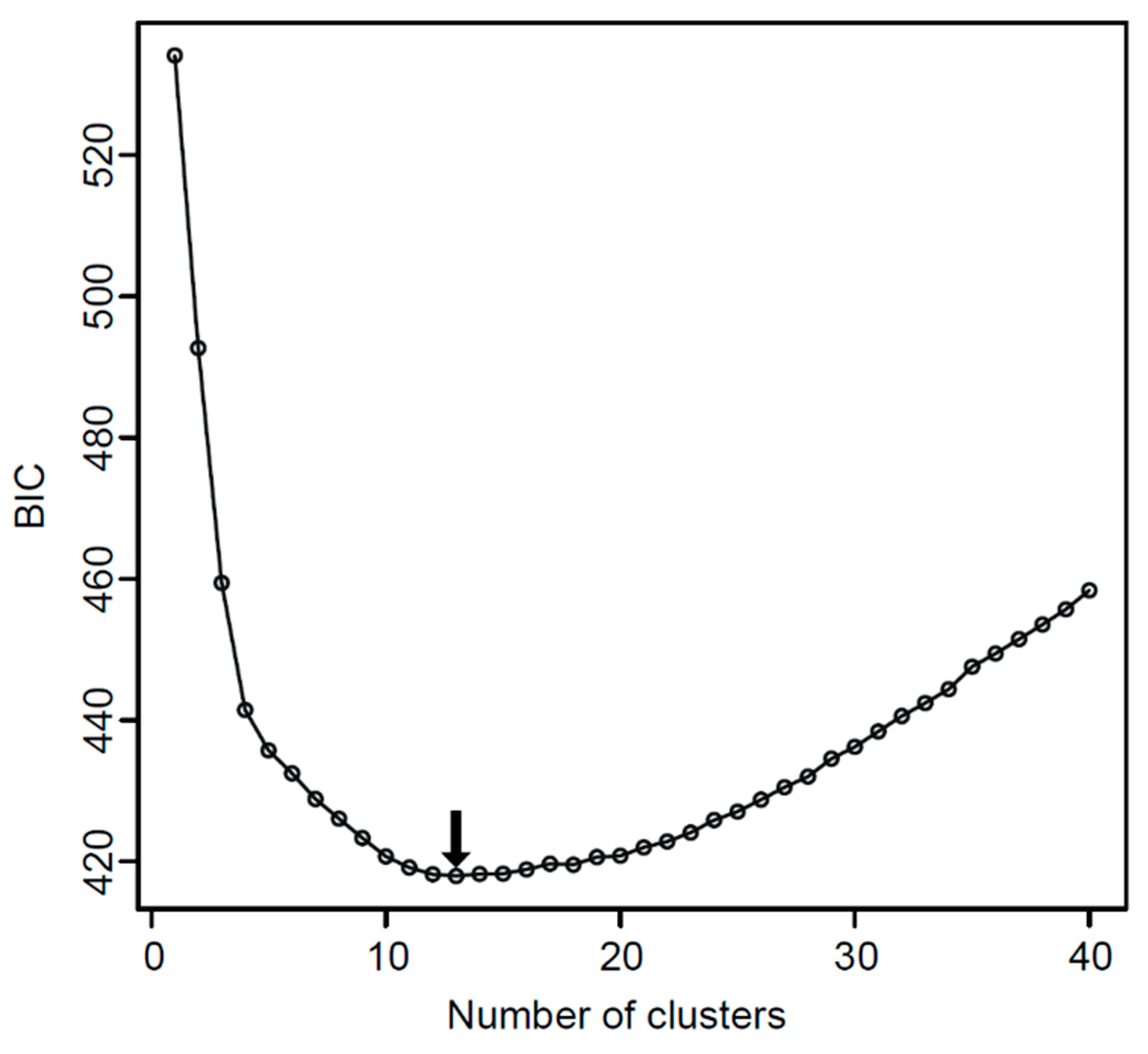
3.3. Morphometric Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fronseca, G.A.B.; Kent, J. Biodiversity hotspots and conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Hubert, N.; Kadarusman; Wibowo, A.; Busson, F.; Caruso, D.; Sulandari, S.; Nafiqoh, N.; Pouyaud, L.; Rüber, L.; Avarre, J.-V.; et al. DNA barcoding indonesian freshwater fishes: Challenges and prospects. DNA Barcodes 2015, 3, 144–169. [Google Scholar]
- Decru, E.; Moelants, T.; de Gelas, K.; Vreven, E.; Verheyen, E.; Snoeks, J. Taxonomic challenges in freshwater fishes: A mismatch between morphology and DNA barcoding in fish of the north-eastern part of the Congo basin. Mol. Ecol. Resour. 2015, 16, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Chapin, S.F.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Laurila-Pant, M.; Lehikoinen, A.; Uusitalo, L.; Venesjärvi, R. How to value biodiversity in environmental management? Ecol. Indic. 2015, 55, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lohbeck, M.; Bongers, F.; Martinez-Ramos, M.; Poorter, L. The importance of biodiversity and dominance for multiple ecosystem functions in a human-modified tropical landscape. Ecology 2016, 97, 2772–2779. [Google Scholar] [CrossRef]
- Staniewicz, A.; Behler, N.; Dharmasyah, S.; Hones, G. Niche partitioning between juvenile sympatric crocodilians in Mesangat Lake, East Kalimantan, Indonesia. Raffles Bull. Zool. 2018, 66, 528–537. [Google Scholar]
- Sudrajat; Saleh, H.A. The potential of Mesangat Lake, East Kutai, Indonesia as an essential ecological area for habitat conservation of critically endangered Crocodylus siamensis. Biodiversitas 2019, 22, 3126–3133. [Google Scholar] [CrossRef]
- Stuebing, R.; Sommerlad, R.; Staniewicz, A. Conservation of the Sunda gharial Tomistoma schlegelii in Lake Mesangat, Indonesia. Int. Zoo Yearb. 2015, 49, 137–149. [Google Scholar] [CrossRef]
- Wijayanti, A.; Suripto, B.A.; Sancayaningsih, R.P. Bird of mesangat in East Kutai, East Kalimantan. AIP Conf. Proc. 2017, 1813, 1–7. [Google Scholar]
- Ahl, E. Einige neue süsswasserfische des indo-malayischen archipels. Ges. Nat. Freunde Berl. 1922, 1992, 30–36. [Google Scholar]
- Kottelat, M. The Fishes of the Mahakam River, East Borneo: An example of the limitations of zoogeographic analysis and the need for extensive fish surveys in Indonesia. Trop. Biodivers. 1995, 2, 401–426. [Google Scholar]
- Nieuwenhuis, A.W. Die durchquerung borneos durch die niederländische Expedition, 1896–1897. Petermanns Geogr. Mitt. 1898, 44, 9–19. [Google Scholar]
- Nieuwenhuis, A.W. Centraal Borneo. Reis von Pontianak naar Samarinda 2; Brill: Leiden, The Netherlands, 1900; pp. 1–520. [Google Scholar]
- Nieuwenhuis, A.W. Quer Durch Borneo, Ergebnisse Seiner Reisen in Den Jahren 1894, 1896–1987 und 1898–1900 Band 2; Brill: Leiden, The Netherlands, 1904–1907; pp. 1–340. [Google Scholar]
- Popta, C.M.L. Descriptions préliminaires des nouvelles espèces de poissons recueillies au Bornéo central par M. le Dr A.W. Nieuwenhuis en 1898 et en 1900. Notes Leyden Mus. 1904, 24, 179–202. [Google Scholar]
- Popta, C.M.L. Suite des descriptions préliminaires des nouvelles espèces de poissons recueillies au Bornéo central par M. le Dr. A. Nieuwenhuis en 1989 et an 1900. Notes Leyden Mus. 1905, 25, 171–186. [Google Scholar]
- Popta, C.M.L. Résultats ichthyologique des voyages scientifiques de Monsieur le professeur Dr. A. W. Nieuwenhuis dans le centre de Bornéo (1898 et 1900). Notes Leyden Mus. 1906, 27, 1–304. [Google Scholar]
- Vaillant, L.L. Résultats zoologiques de l’expédition scientifique Néerlandaise au Bornéo central. Poissons. Notes Leyden Mus. 1902, 24, 1–166. [Google Scholar]
- Christensen, M.S. Investigations on the ecology and fish fauna of the Mahakam River in east Kalimantan (Borneo), Indonesia. Int. Rev. Hydrobiol. 1992, 77, 593–608. [Google Scholar] [CrossRef]
- Hadiaty, R.K.; Kottelat, M. Rasbora lacrimula, a new species of cyprinid fish from eastern Borneo (Teleostei: Cyprinidae). Ichthyol. Explor. Freshw. 2009, 20, 105–109. [Google Scholar]
- Hadiaty, R.K.; Kottelat, M. Pangio lidi, a new species of loach from eastern Borneo, Indonesia (Teleostei: Cobitidae). Zootaxa 2009, 2171, 65–68. [Google Scholar] [CrossRef]
- Kottelat, M. Rasbora rheophila, a new species of fish from northern Borneo (Teleostei: Cyprinidae). Rev. Suisse Zool. 2012, 119, 77–87. [Google Scholar] [CrossRef]
- Kottelat, M.; Tan, H.H. Rasbora atranus, a new species of fish from central Borneo (Teleostei: Cyprinidae). Ichthyol. Explor. Freshw. 2011, 22, 215–220. [Google Scholar]
- Siebert, D.J.; Richardson, P.J. Rasbora latoclava, a new cyprinid from Kalimantan, Indonesia, and lectotype designation for R. vaillantii. Ichthyol. Explor. Freshw. 1997, 8, 89–95. [Google Scholar]
- Lumbantobing, D.N. Four new species of Rasbora of the Sumatrana group (Teleostei: Cyprinidae) from northern Sumatra, Indonesia. Zootaxa 2014, 3764, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Sholihah, A.; Delrieu-Trottin, E.; Sukmono, T.; Dahruddin, H.; Risdawati, R.; Elvyra, R.; Wibowo, A.; Kustiati, K.; Busson, F.; Sauri, S.; et al. Disentangling the taxonomy of the subfamily Rasborinae (Cypriniformes, Danionidae) in Sundaland using DNA barcodes. Sci. Rep. 2020, 10, 2818. [Google Scholar] [CrossRef] [Green Version]
- Kottelat, M.; Tan, H.H. Rasbora cryptica, a new species of fish from Sarawak, Borneo (Teleostei: Cyprinidae). Ichthyol. Explor. Freshw. 2012, 23, 37–44. [Google Scholar]
- Brittan, M.R. A Revision of the Indo-Malayan Fresh-Water Fish Genus Rasbora; Monograph Institute of Science and Technology: Manila, Philippines, 1954; pp. 1–224. [Google Scholar]
- Kottelat, M.; Vidthayanon, C. Boraras micros, a new genus and species of minute freshwater fish from Thailand (Teleostei: Cyprinidae). Ichthyol. Explor. Freshw. 1993, 4, 161–176. [Google Scholar]
- Liao, T.Y.; Kullander, S.O.; Fang, F. Phylogenetic analysis of the genus Rasbora (Teleostei: Cyprinidae). Zool. Scr. 2011, 39, 155–176. [Google Scholar] [CrossRef]
- Tang, K.L.; Agnew, M.K.; Hirt, M.V.; Sado, T.; Schneider, L.M.; Freyhof, J.; Sulaiman, Z.; Swartz, E.; Vidthayanon, C.; Miya, M.; et al. Systematics of the subfamily Danioninae (Teleostei: Cypriniformes: Cyprinidae). Mol. Phylogenet. Evol. 2010, 57, 189–214. [Google Scholar] [CrossRef]
- Inger, R.F.; Ching, P.K. The freshwater fishes of North Borneo. Fieldiana Zool. 1962, 45, 1–268. [Google Scholar]
- Kottelat, M.; Whitten, A.J.; Kartikasari, S.N.; Wirjoatmodjo, S. Freshwater Fishes of Western Indonesia and Sulawesi, Indonesia; Periplus Editions Ltd.: Hong Kong, China, 1993; pp. 1–259. [Google Scholar]
- Weber, M.; de Beaufort, L.F. The fishes of the Indo-Australian Archipelago. III. Ostariophysi: II Cyprinoidea, Apodes, Synbranchi; Brill: Leiden, The Netherlands, 1916; pp. 1–455. [Google Scholar]
- Britz, R.; Tan, H.H. Rasbora simonbirchi, a new species of cyprinid fish from Central Kalimantan, Borneo. Ichthyol. Explor. Freshw. 2018, 28, 337–344. [Google Scholar]
- Hui, T.H.; Kottelat, M. Rasbora marinae, a new species of cyprinid fish from northwestern Borneo (Teleostei: Danionidae). Raffles Bull. Zool. 2020, 68, 750–758. [Google Scholar]
- Lumbantobing, D.N. Four new species of the Rasbora trifasciata-group (Teleostei: Cyprinidae) from Northwestern Sumatra, Indonesia. Copeia 2010, 4, 644–670. [Google Scholar] [CrossRef]
- Bengtson, P. Open nomenclature. Palaeontology 1988, 16, 713–719. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 8 November 2020).
- Ivanova, N.V.; Zemlak, T.S.; Hanner, T.H.; Hebert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega 6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T.J. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1958, 22, 160–174. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Masters, B.C.; Fan, V.; Ross, H.A. Species delimitation—A geneious plugin for the exploration of species boundaries. Mol. Ecol. Resour. 2011, 11, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Kapli, P.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate poisson tree processes for single-locus species delimitation under Maximum Likelihood and Markov Chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Exelixis Lab. MPTP. Available online: https://mptp.h-its.org/#/tree (accessed on 5 December 2020).
- Rohlf, F.J. TPSDIG2: A Program for Landmark Development and Analysis. Available online: http://life.bio.sunysb.edu/morph/ (accessed on 8 November 2020).
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer; Elsevier: San Diego, CA, USA, 2004; pp. 1–443. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Sidlauskas, B.L.; Mol, J.H.; Vari, R.P. Dealing with allometry in linear and geometric morphometrics: A taxonomic case study in the Leporinus cylindriformis group (Characiformes: Anostomidae) with description of a new species from Suriname. Zool. J. Linn. Soc. 2011, 162, 103–130. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Collins, C. A Tutorial for Discriminant Analysis of Principal Components (DAPC) Using Adegenet 2.0.0. Available online: http://adegenet.r-forge.r-project.org/files/tutorial-dapc.pdf (accessed on 8 November 2020).
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.R-project.org (accessed on 8 November 2020).
- Kottelat, M. Notes on the taxonomy of some Sundaic and Indochinese species of Rasbora, with description of four new species (Pisces: Cyprinidae). Ichthyol. Explor. Freshw. 1991, 2, 177–191. [Google Scholar]
- Rainboth, W.J. Fishes of the Combodian Mekong; FAO: Rome, Italy, 1996; pp. 1–265. [Google Scholar]
- Mayden, R.L.; Conway, K.W.; Tang, K.; Freyhof, J.; Chamberlain, S.; Haskins, M.; Schneider, L.; Sudkamp, M.; Wood, R.M.; Agnew, M.; et al. Phylogenetic relationships of Danio within the order cypriniformes: A framework for comparative and evolutionary studies of a model species. J. Exp. Zool. B Mol. Dev. Evol. 2007, 308, 642–654. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D. DNA barcode divergence among species and genera of birds and fishes. Mol. Ecol. Resour. 2009, 9, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Herder, F.; Nolte, A.W.; Pfaender, J.; Schwarzer, J.; Hadiaty, R.K.; Schliewen, U.K. Adaptive radiation and hybridization in Wallace’s Dreamponds: Evidence from sailfin silversides in the Malili Lakes of Sulawesi. Proc. R. Soc. B 2006, 273, 2209–2217. [Google Scholar] [CrossRef] [PubMed]
- April, J.; Mayden, R.L.; Hanner, R.H.; Bernatchez, L. Genetic calibration of species diversity among North America’s freshwater fishes. Proc. Natl. Acad. Sci. USA 2011, 108, 10602–10607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Kapil, S.; Chawla, M.; Ansari, M.D. On k-means data clustering algorithm with genetic algorithm. In Proceedings of the Fourth International Conference on Parallel, Distributed and Grid Computing (PDGC), Solan, India, 22–24 December 2016; pp. 202–206. [Google Scholar]
- Fraley, C.; Raftery, A.E. How many clusters? Which clustering method? Answers via model-based cluster analysis. Comput. J. 1998, 41, 578–588. [Google Scholar] [CrossRef]
- Gasch, A.P.; Eisen, M.B. Exploring the conditional coregulation of yeast gene expression through fuzzy k-means clustering. Genome Biol. 2002, 3, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.X.; Zhang, W.J.; Kusalik, A.J. A genetic k-means clustering algorithm applied to gene expression data. In Proceedings of the Sixteenth Canadian Conference on Artificial Intelligence, Halifax, NS, Canada, 11–13 June 2003; Xiang, Y., Chaib-draa, B., Eds.; Springer: Heidelberg, Germany, 2003; pp. 520–526. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Coordinates | Date(s) in September 2014 | Number of Collected Rasborines |
|---|---|---|---|
| S. Mesangat Ulu | 0.502 116.698 | 2, 10, 11, 25 | 145 |
| Sungai Sebastian | 0.502 116.699 | 8, 22, 23 | 66 |
| Loa Blöma 1 | 0.503 116.697 | 4, 23 | 206 |
| Loa Kahang | 0.504 116.698 | 9, 21, 24 | 37 |
| Loa Lakum 1.1 | 0.502 116.698 | 2, 11 | 48 |
| Loa Lakum 1.3 | 0.500 116.699 | 2 | 9 |
| Loa Lakum 2.1 | 0.502 116.698 | 4, 22 | 118 |
| Loa Lakum 3.1 | 0.500 116.699 | 6 | 10 |
| Loa Lakum 3.3 | 0.500 116.699 | 16 | 100 |
| Loa Sangkal 1 | 0.501 116.698 | 6, 7 | 18 |
| Loa Sangkal 2 | 0.502 116.698 | 15 | 17 |
| Loa Toh 1 | 0.497 116.697 | 7 | 1 |
| Long Potok 2 | 0.492 116.700 | 20 | 30 |
| Mesangat Ilir 1 | 0.493 166.685 | 17 | 5 |
| Mesangat River | 0.494 116.697 | 11 | 11 |
| Mesangat Swamp 2 | 0.508 116.697 | 8, 24 | 107 |
| unknown | - | - | 18 |
| Total sample number | 946 |
| Rasbora vaillantii | Rasbora rutteni | Rasbora hubbsi | ||||||
|---|---|---|---|---|---|---|---|---|
| Author | Popta [17] | Weber and de Beaufort [35] | Weber and de Beaufort [35] | Brittan [29] | Kottelat et al. [34] | Brittan [29] | Kottelat et al. [34] | |
| Coloration | Lateral Stripe | Dark, narrower anteriorly | Broader posteriorly | Broader and darker posteriorly | Broader and darker posteriorly | Indistinct anteriorly | Slightly darker and narrower posteriorly; ends in spot posteriorly | Indistinct anteriorly |
| Axial Streak | Separated from lateral stripe anteriorly | - | - | Above lateral stripe anteriorly; coincident posteriorly | - | Weaker anteriorly, above lateral stripe anteriorly | - | |
| Anal Streak | Dark, forms subpeduncular streak posteriorly | Dark, forms subpeduncular streak posteriorly | - | Some melanophores | - | Distinct; forms subpeduncular streal posteriorly | - | |
| Dorsal Stripe | Dorsal part of caudal peduncle | Dorsal part of caudal peduncle | Dorsal part of caudal peduncle | Distinct | - | Distinct | - | |
| Fins | Hyaline | - | Hyaline | Hyaline; in life sometimes rosy | - | Hyaline | - | |
| Caudal-Fin | Dark margin, blotch at lobes | Blackish border | - | - | - | yellowish at base of lobes | - | |
| Finrays | Dorsal | II,7 | II,7 | II,7 | II,7 | - | II,7 | - |
| Pectoral | I,15 | I,15 | I,12; not reaching pelvics | I,11-12; not reaching pelvics | - | I,11-13 | - | |
| Pelvic | II,8 | II,8; not reaching anal | II,7; reaching anal | II,7; not reaching anal | - | II,7-8 | - | |
| Anal | III,5-6 | III,5 | III,5 | III,5 | - | III,5 | - | |
| Caudal | 1/17/1 | - | - | 6/1/17/1/8 | - | 8/1/17/1/8 | - | |
| Scales | Lateral Line | 27–30 | 27–30 | 26 | 26–29; 24–26 perforated | 26–29; 24–26 perforated | 28–31; 25–29 perforated | 29–30; 25–26 perforated |
| Transverse Row | 4,5/1/3,5 | 4,5/1/3,5 | 4,5/1/3,5 | - | 4,5/1/2,5-3,5 | - | ||
| Predorsal | - | - | 10 | 10 | - | 10-12 | - | |
| Caudal Peduncle | - | - | - | 12 | - | 12-14 | 14 | |
| Measurements | Dorsohypural | Not reaching snout | - | - | Fallen on anterior margin of pupil | - | Fallen on snout tip | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hüllen, S.; Mandl, C.; Geiger, M.F.; Hadiaty, R.K.; Wahyudewantoro, G.; Ziegler, T.; Herder, F. On the Challenge to Correctly Identify Rasboras (Teleostei: Cyprinidae: Danioninae) Inhabiting the Mesangat Wetlands, East Kalimantan, Indonesia. Diversity 2021, 13, 8. https://doi.org/10.3390/d13010008
Hüllen S, Mandl C, Geiger MF, Hadiaty RK, Wahyudewantoro G, Ziegler T, Herder F. On the Challenge to Correctly Identify Rasboras (Teleostei: Cyprinidae: Danioninae) Inhabiting the Mesangat Wetlands, East Kalimantan, Indonesia. Diversity. 2021; 13(1):8. https://doi.org/10.3390/d13010008
Chicago/Turabian StyleHüllen, Sebastian, Chiara Mandl, Matthias F. Geiger, Renny K. Hadiaty, Gema Wahyudewantoro, Thomas Ziegler, and Fabian Herder. 2021. "On the Challenge to Correctly Identify Rasboras (Teleostei: Cyprinidae: Danioninae) Inhabiting the Mesangat Wetlands, East Kalimantan, Indonesia" Diversity 13, no. 1: 8. https://doi.org/10.3390/d13010008
APA StyleHüllen, S., Mandl, C., Geiger, M. F., Hadiaty, R. K., Wahyudewantoro, G., Ziegler, T., & Herder, F. (2021). On the Challenge to Correctly Identify Rasboras (Teleostei: Cyprinidae: Danioninae) Inhabiting the Mesangat Wetlands, East Kalimantan, Indonesia. Diversity, 13(1), 8. https://doi.org/10.3390/d13010008