Red Imported Fire Ants Reduce Invertebrate Abundance, Richness, and Diversity in Gopher Tortoise Burrows


Abstract
:1. Introduction
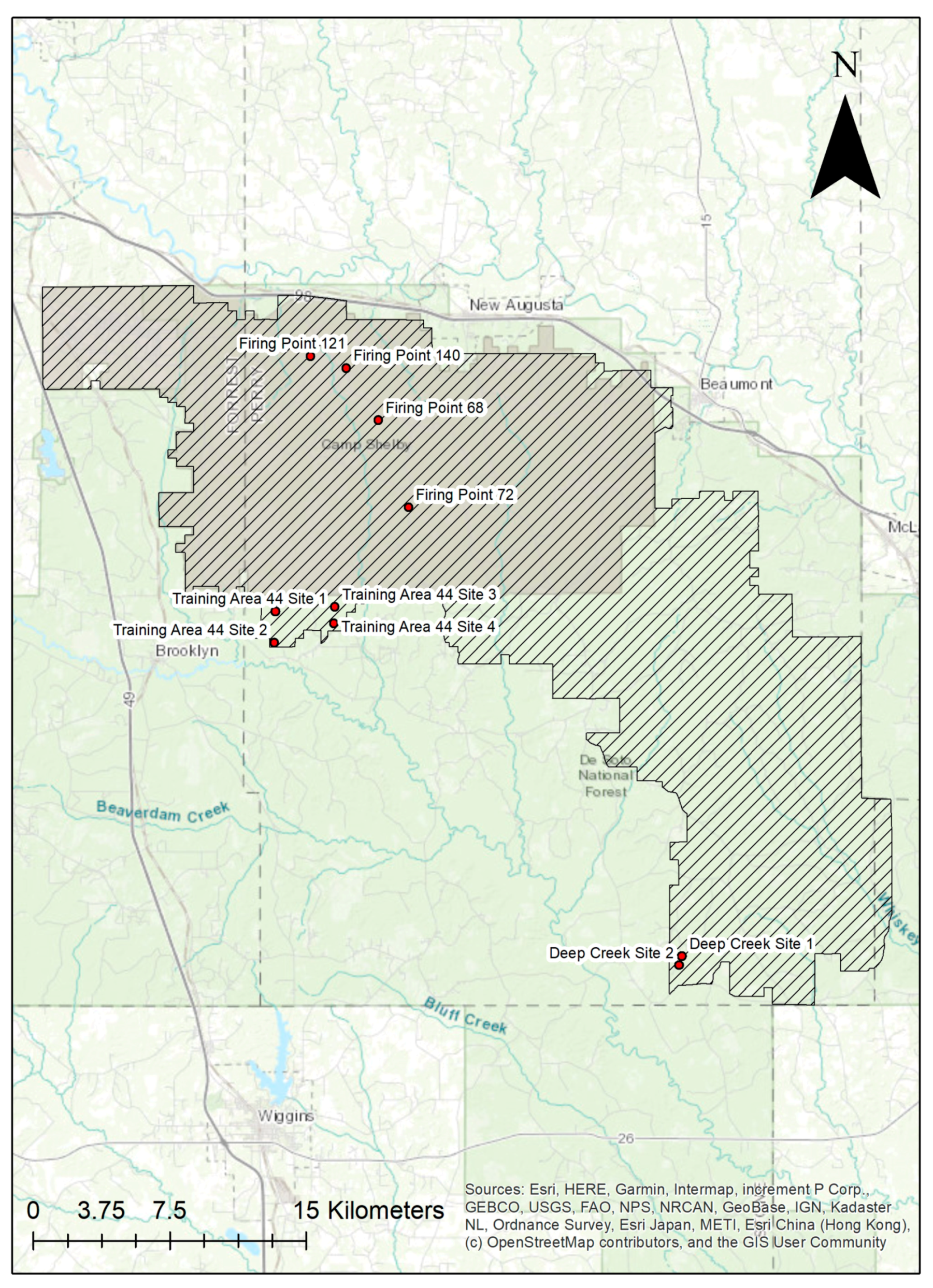
2. Materials and Methods
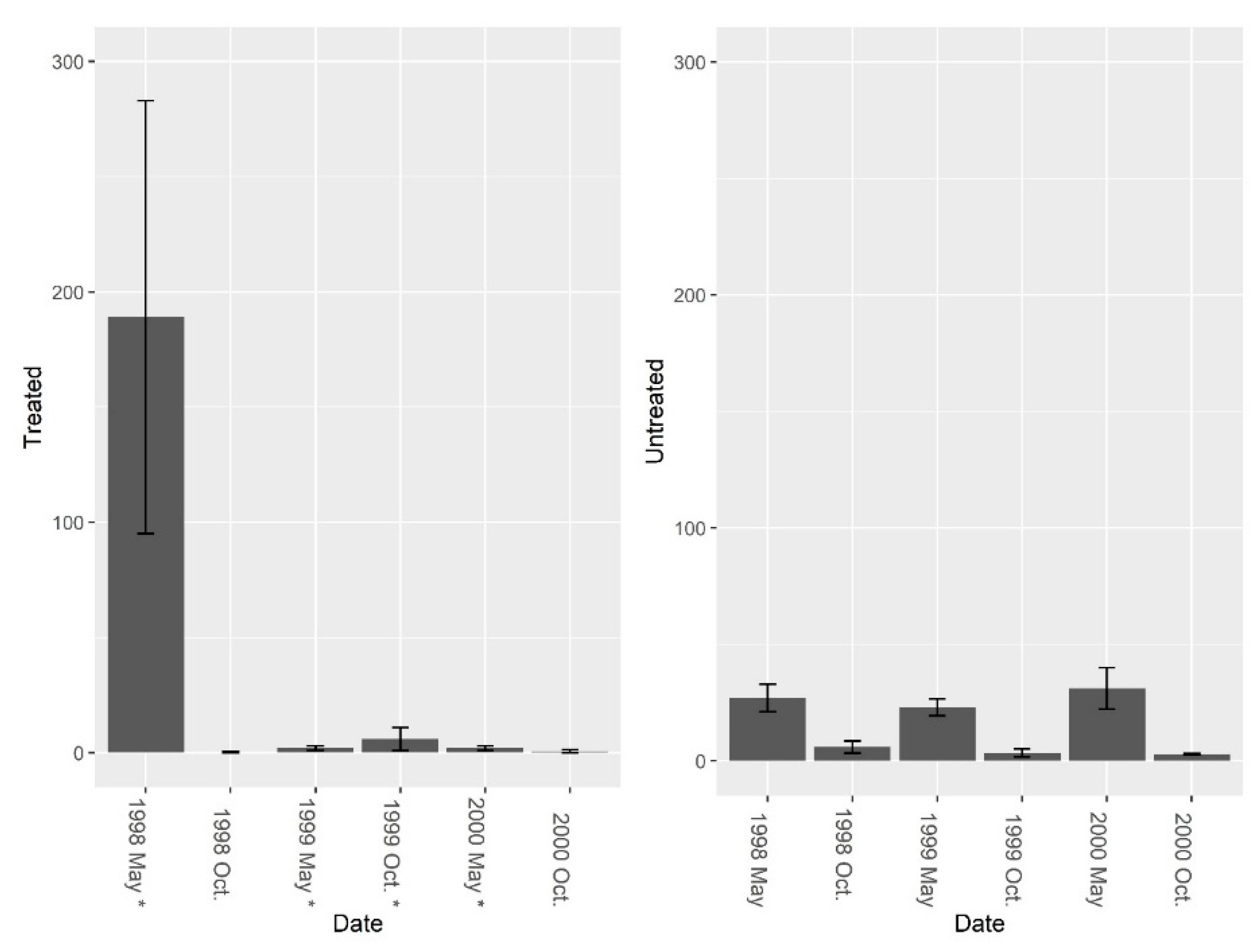
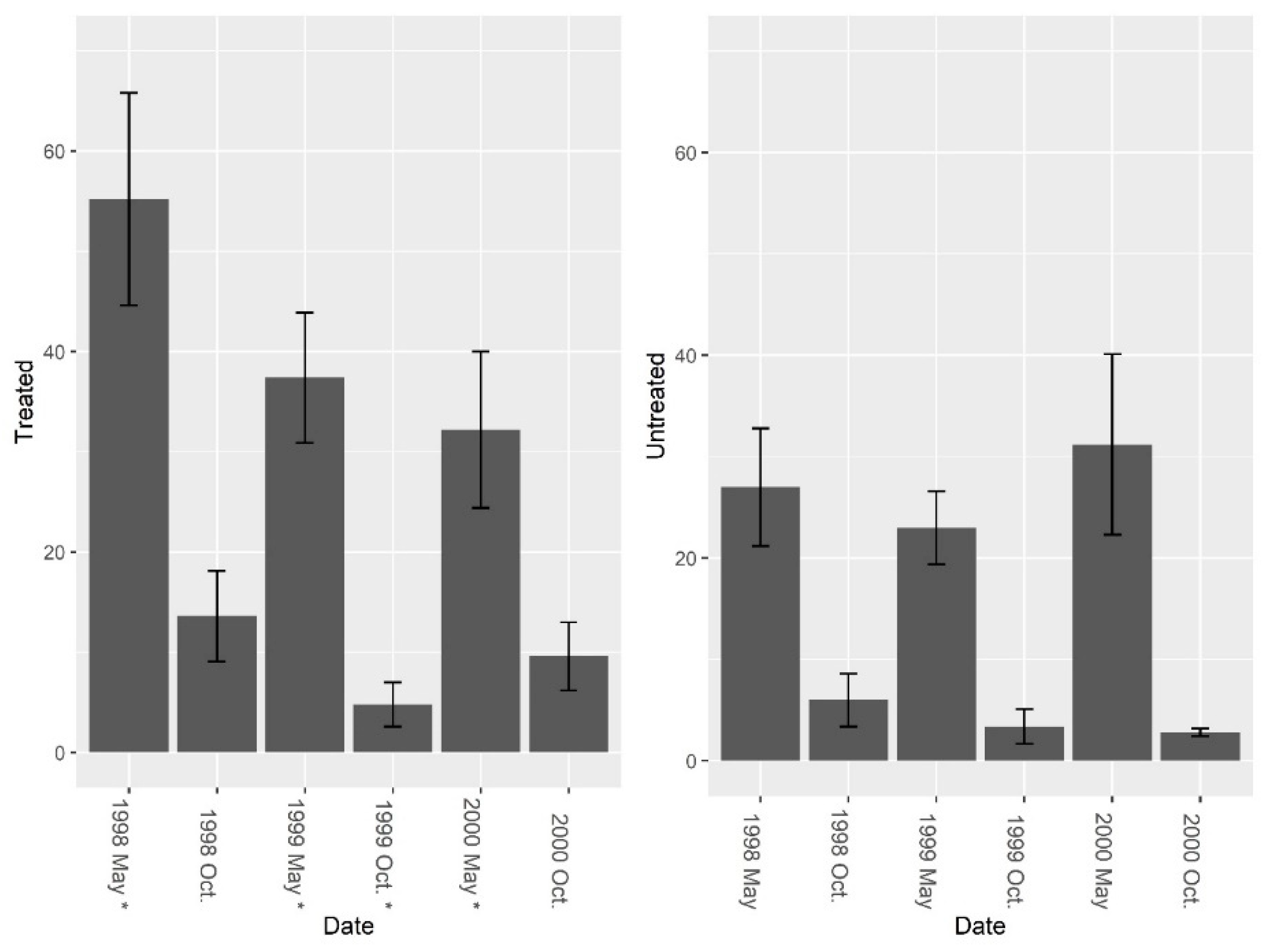
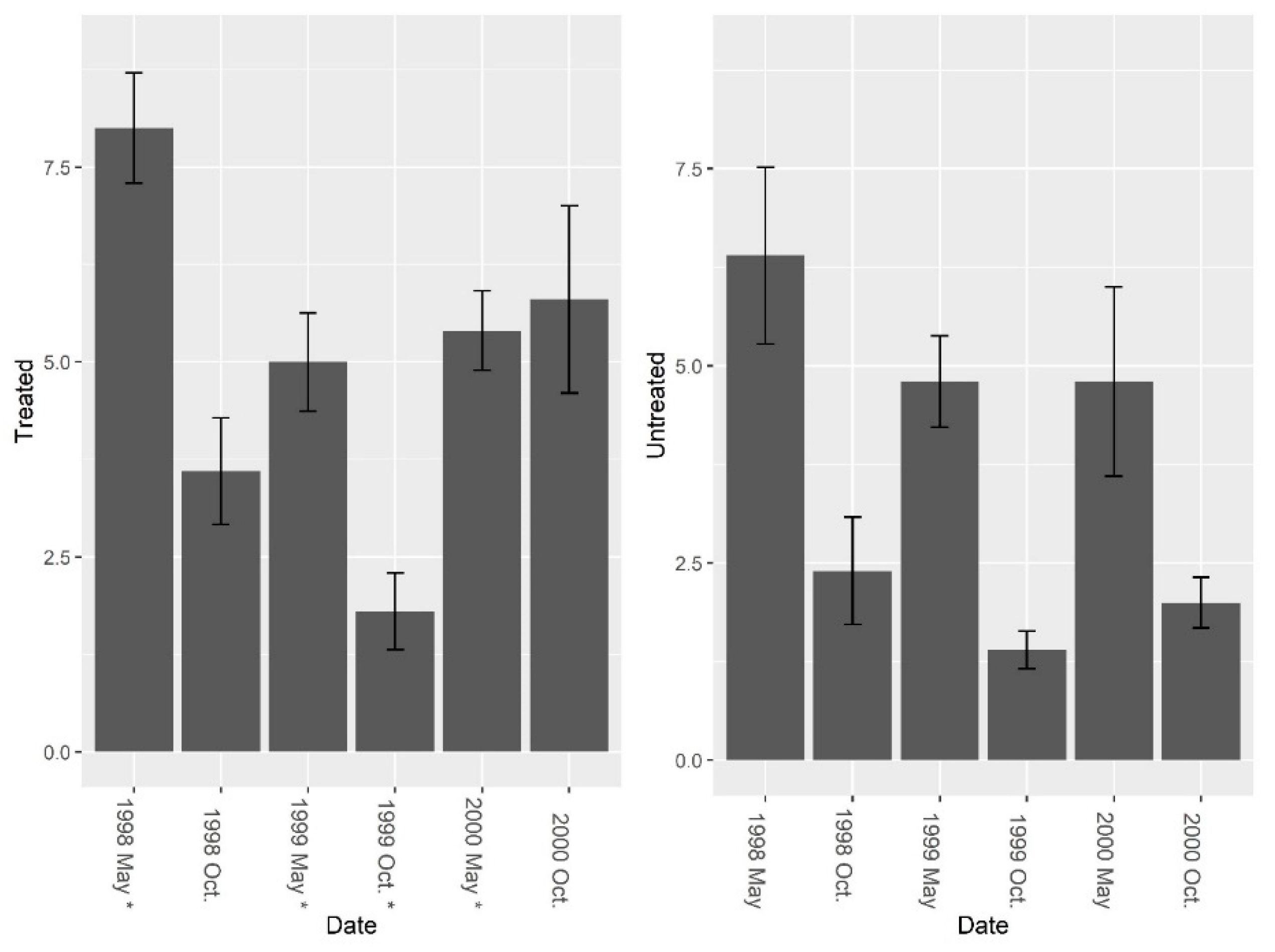
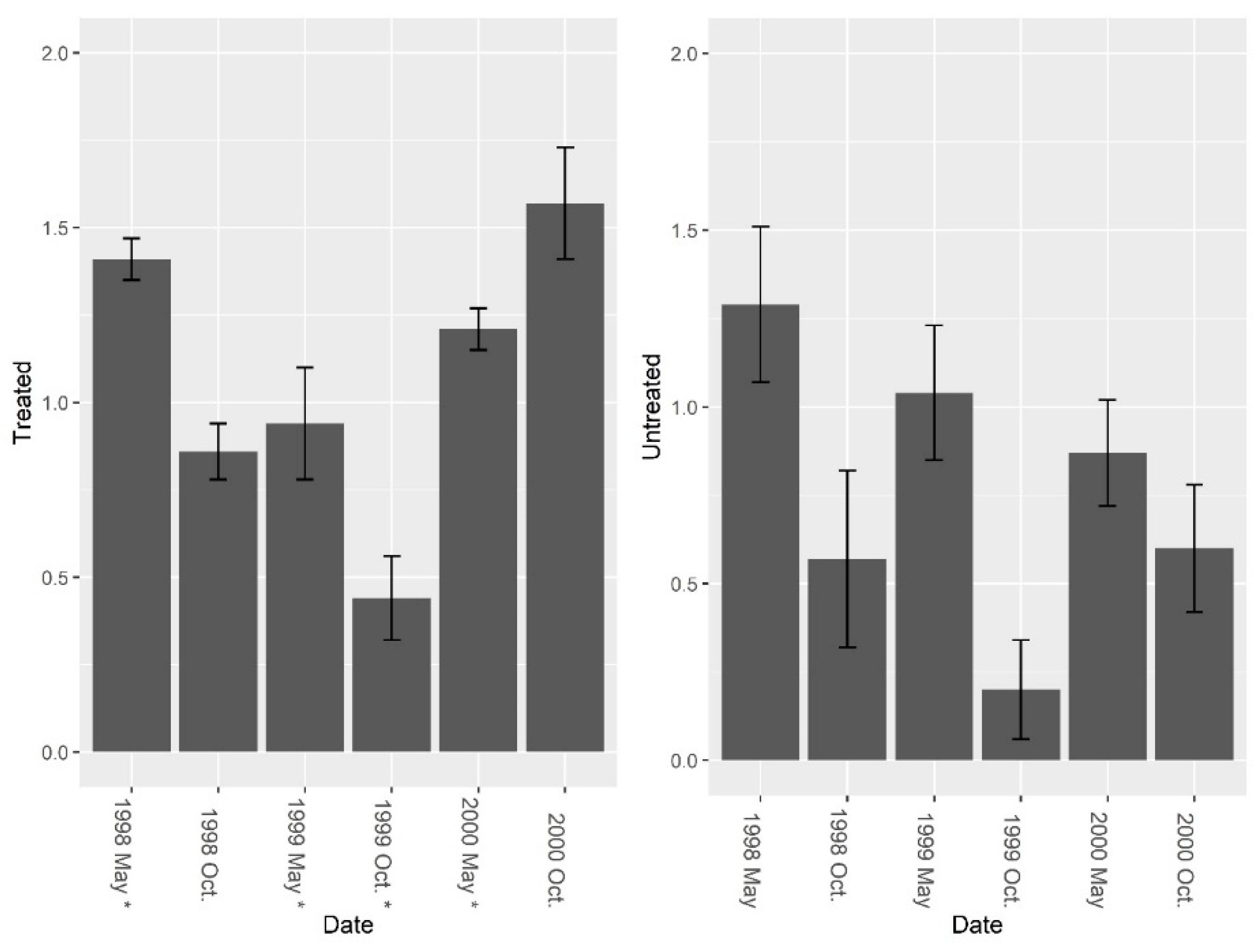
3. Results
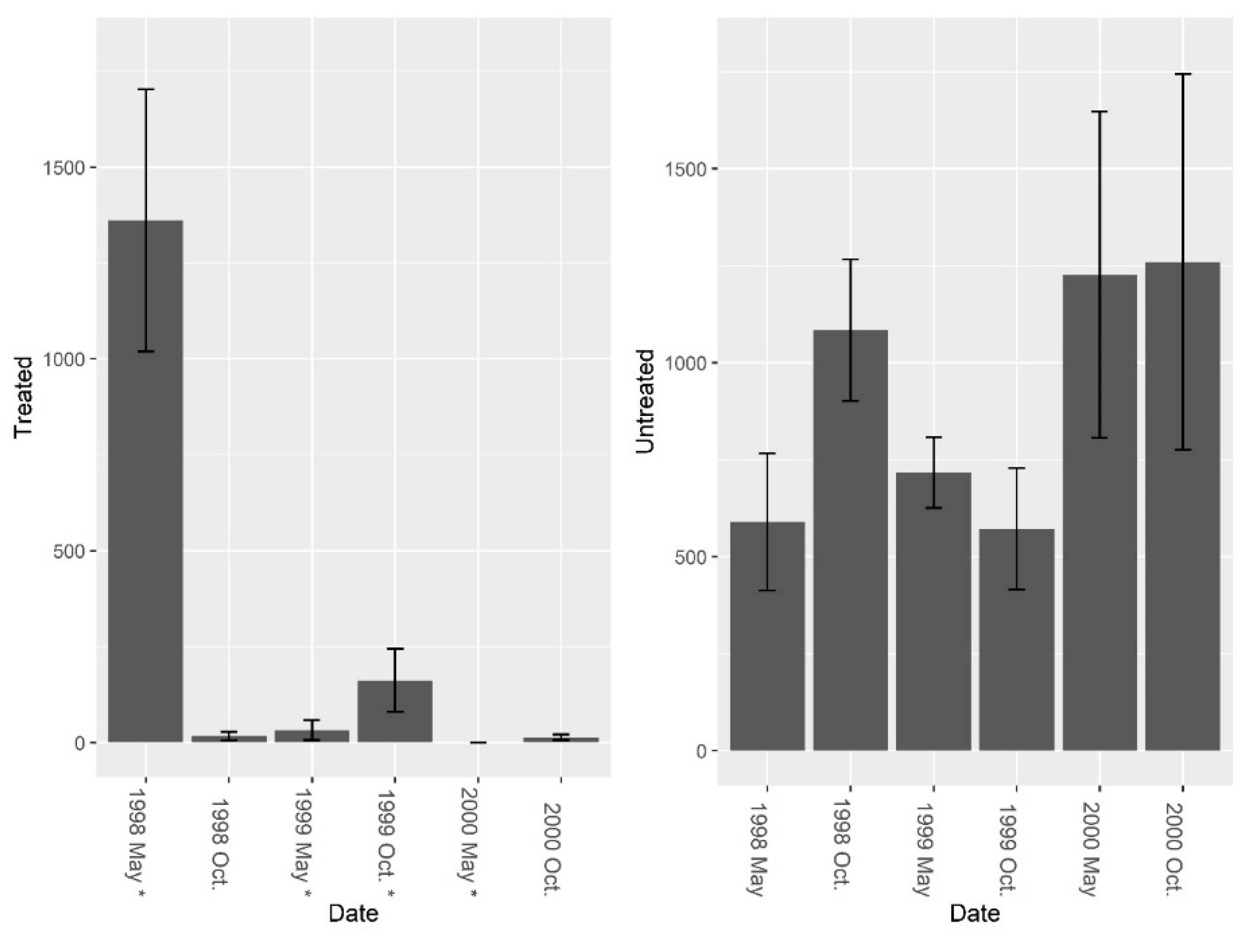
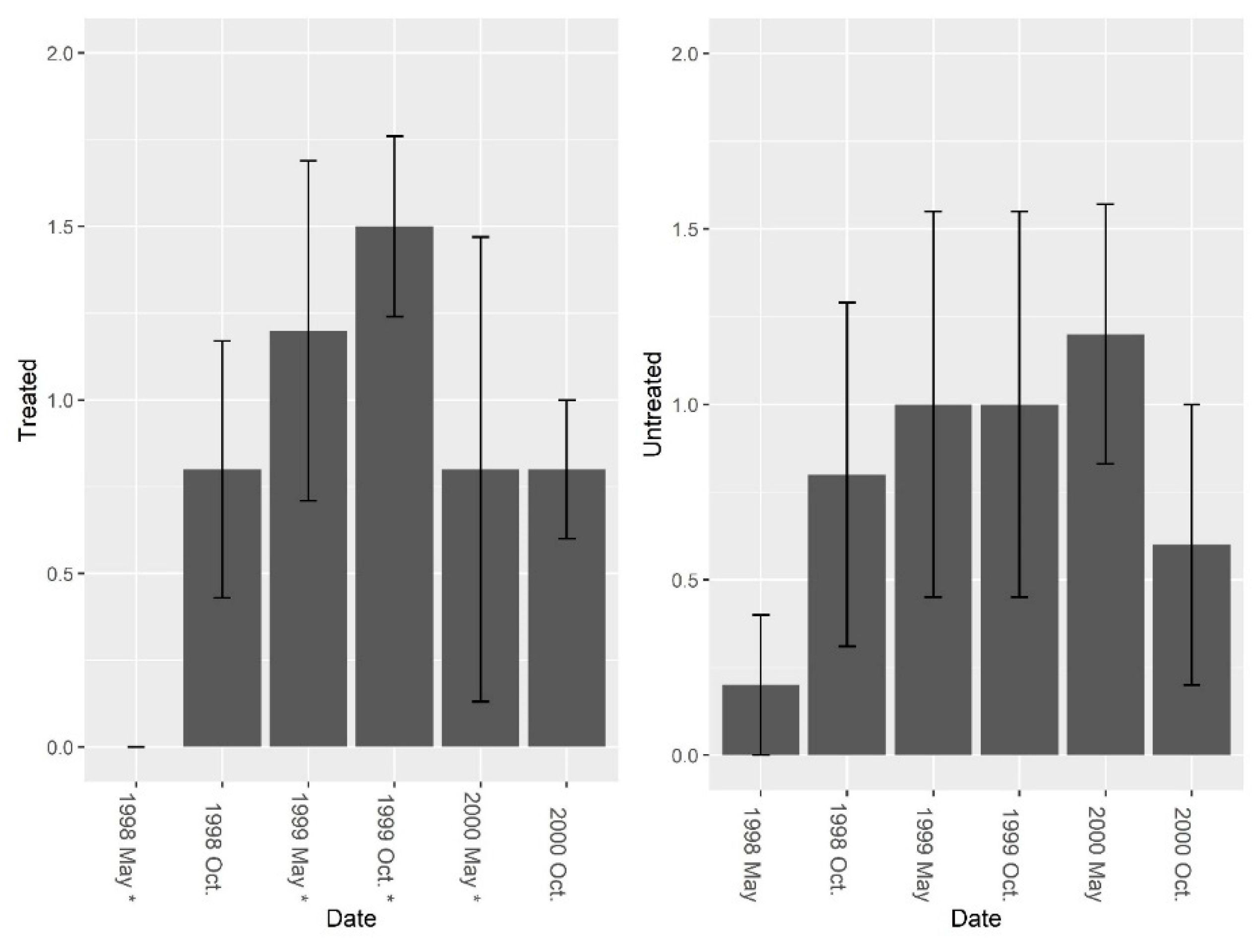
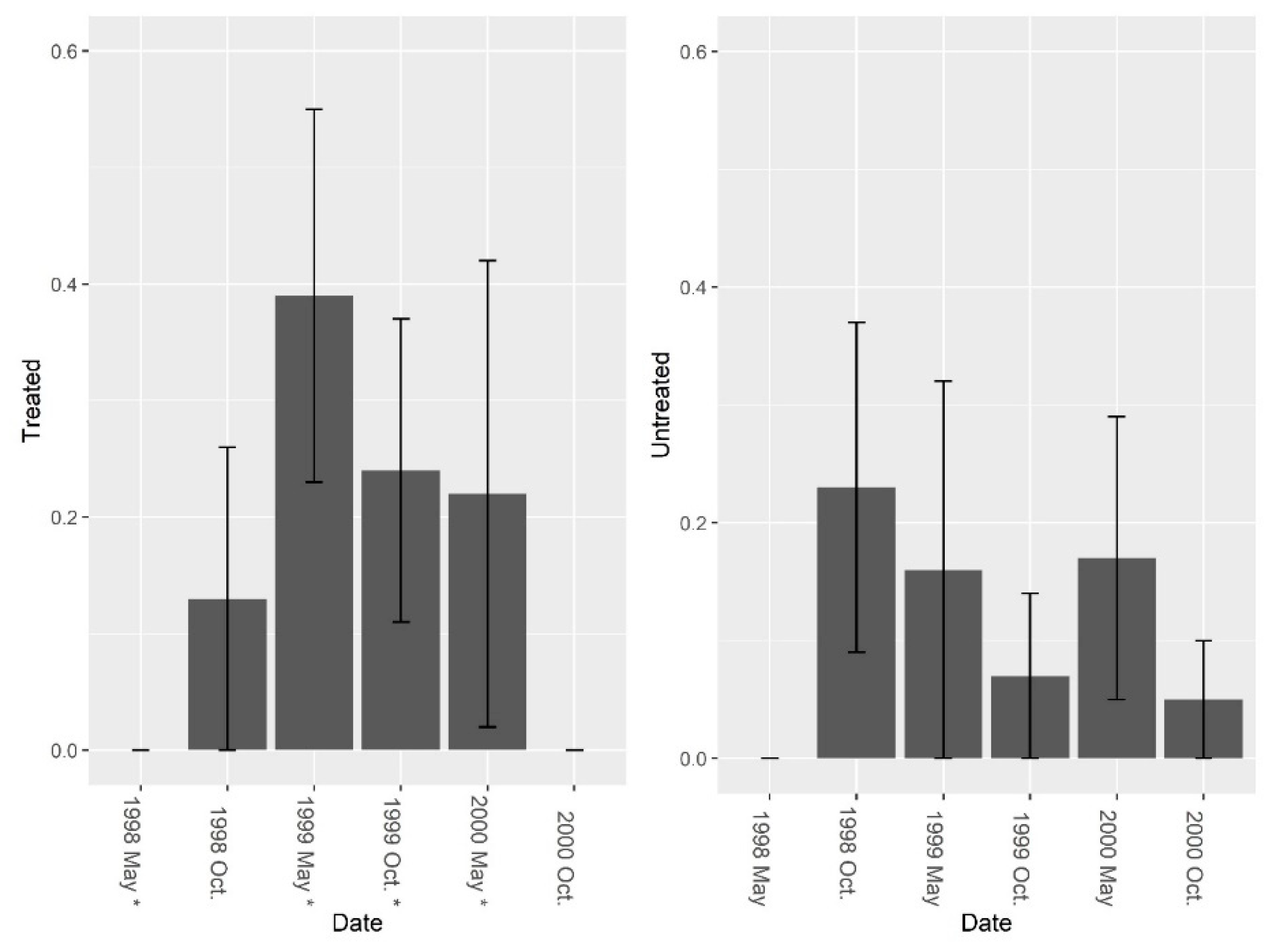
3.1. Burrow Baits
3.2. Burrow Vacuums
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Northing | Easting |
|---|---|---|
| Deep Creek Site 1 | 3423586 | 315624 |
| Deep Creek Site 2 | 3423153 | 315479 |
| Training Area 44 Site 1 | 3440128 | 296754 |
| Training Area 44 Site 2 | 3438659 | 296666 |
| Training Area 44 Site 3 | 3440288 | 299548 |
| Training Area 44 Site 4 | 3439513 | 299489 |
| Firing Point 68 | 3449013 | 301776 |
| Firing Point 72 | 3444891 | 303116 |
| Firing Point 121 | 3452081 | 298644 |
| Firing Point 140 | 3451485 | 300308 |
References
- Wilson, E.O. (Ed.) Biodiversity; The National Academies Press: Washington, DC, USA, 1988. [Google Scholar]
- Porter, S.D.; Savignano, D.A. Invasion of Polygyne Fire Ants Decimates Native Ants and Disrupts Arthropod Community. Ecology 1990, 71, 2095–2106. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks-Leite, C.; Pardini, R.; Tambosi, L.R.; Pearse, W.D.; Bueno, A.A.; Bruscagin, R.T.; Condez, T.H.; Dixo, M.; Igari, A.T.; Martensen, A.C.; et al. Using ecological thresholds to evaluate the costs and benefits of set-asides in a biodiversity hotspot. Science 2014, 345, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Poynton, J.C.; Loader, S.P.; Sherratt, E.; Clarke, B.T. Biodiversity hotspots for conservation priorities. Biodivers. Conserv. 2007, 16, 853–858. [Google Scholar] [CrossRef]
- Roberts, C.M.; McClean, C.J.; Veron, J.E.N.; Hawkins, J.P.; Allen, G.R.; McAllister, D.E.; Mittermeier, C.G.; Schueler, F.W.; Spalding, M.; Wells, F.; et al. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 2002, 295, 1280–1284. [Google Scholar] [CrossRef] [Green Version]
- Kinlaw, A.; Grasmueck, M. Evidence for and geomorphologic consequences of a reptilian ecosystem engineer: The burrowing cascade initiated by the Gopher Tortoise. Geomorphology 2012, 157–158, 108–121. [Google Scholar] [CrossRef]
- Alexy, K.J.; Brunjes, K.J.; Gassett, J.W.; Miller, K.V.; Alex, K.J.; Brunjes, K.J.; Gassett, J.W.; Miller, K.V. Continuous Remote Monitoring of Gopher Tortoise Burrow Use. Wildl. Soc. Bull. 2003, 31, 1240–1243. [Google Scholar]
- Jackson, D.R.; Milstrey, E.G. The fauna of gopher tortoise burrows. In Proceedings of the Gopher Tortoise Relocation Symposium, Tallahassee, FL, USA; 1989; pp. 86–98. [Google Scholar]
- Guyer, C.; Hermann, S.M. Patterns of size and longevity of gopher tortoise (Gopherus polyphemus) burrows: Implications for the longleaf pine ecosystem. Chelonian Conserv. Biol. 1997, 2, 507–513. [Google Scholar]
- Milstrey, E.G. Ticks and invertebrate commensals in Gopher Tortoise burrows: Implications and importance. In Proceedings of the 5th Annual. Meeting Gopher Tortoise Council, Gainesville, FL, USA; 1986; pp. 4–15. [Google Scholar]
- Pike, D.A.; Mitchell, J.C. Burrow-dwelling ecosystem engineers provide thermal refugia throughout the landscape. Anim. Conserv. 2013, 16, 694–703. [Google Scholar] [CrossRef]
- Ennen, J.R.; Kreiser, B.R.; Qualls, C.P. Low Genetic Diversity in Several Gopher Tortoise (Gopherus polyphemus) Populations in the Desoto National Forest, Mississippi. Herpetologica 2010, 66, 31–38. [Google Scholar] [CrossRef]
- Clostio, R.W.; Martinez, A.M.; Leblanc, K.E.; Anthony, N.M. Population genetic structure of a threatened tortoise across the south-eastern United States: Implications for conservation management. Anim. Conserv. 2012, 15, 613–625. [Google Scholar] [CrossRef]
- Lago, P.K. A survey of arthropods associated with Gopher Tortoise burrows in Mississippi. Entomol. News 1991, 102, 1–13. [Google Scholar]
- Callcott, A.-M.A.; Collins, H.L. Invasion and Range Expansion of Imported Fire Ants (Hymenoptera: Formicidae) in North America from 1918–1995. Florida Entomol. 1996, 79, 240–251. [Google Scholar] [CrossRef]
- The University of Georgia EDDMapS: Red Imported Fire Ant Solenopsis Invicta (Buren). Available online: https://www.eddmaps.org/distribution/viewmap.cfm?sub=4675 (accessed on 23 November 2020).
- Epperson, D.M.; Heise, C.D. Nesting and hatchling ecology of Gopher Tortoises (Gopherus polyphemus) in southern Mississippi. J. Herpetol. 2003, 37, 315–324. [Google Scholar] [CrossRef]
- Jusino-Atresino, R.; Phillips, S.A. Impact of Red Imported Fire Ants on the Ant Fauna of Texas; Westview Press: Boulder, CO, USA, 1994. [Google Scholar]
- Morris, J.R.; Steigman, K.L. Southwestern Association of Naturalists Effects of Polygyne Fire Ant Invasion on Native Ants of a Blackland Prairie in Texas. Southwest. Nat. 1993, 38, 136–140. [Google Scholar] [CrossRef]
- Epperson, D.M.; Allen, C.R. Red Imported Fire Ant Impacts on Upland Arthropods in Southern Mississippi. Am. Midl. Nat. 2010, 163, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Kenis, M.; Auger-Rozenberg, M.A.; Roques, A.; Timms, L.; Péré, C.; Cock, M.J.W.; Settele, J.; Augustin, S.; Lopez-Vaamonde, C. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Holway, D.A.; Lach, L.; Suarez, A.V.; Tsutsui, N.D.; Case, T.J. The causes and consequences of ant invasions. Annu. Rev. Ecol. Syst. 2002, 33, 181–233. [Google Scholar] [CrossRef] [Green Version]
- Boivin, N.L.; Zeder, M.A.; Fuller, D.Q.; Crowther, A.; Larson, G.; Erlandson, J.M.; Denhami, T.; Petraglia, M.D. Ecological consequences of human niche construction: Examining long-term anthropogenic shaping of global species distributions. Proc. Natl. Acad. Sci. USA 2016, 113, 6388–6396. [Google Scholar] [CrossRef] [Green Version]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic Invasions: Causes, Epidemiology, Global Consequences, and Control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Dziadzio, M.C.; Long, A.K.; Smith, L.L.; Chandler, R.B.; Castleberry, S.B. Presence of the red imported fire ant at gopher tortoise nests. Wildl. Soc. Bull. 2016, 40, 202–206. [Google Scholar] [CrossRef]
- Jennings, R.; Fritts, T. The Status of the Gopher Tortoise, Gopherus polyphemus Daudin; Unpublished Final Report; United States Fish and Wildlife Service: Jackson, MS, USA, 1983. [Google Scholar]
- Wetterer, J.K.; Moore, J.A. Red Imported Fire Ants (Hymenoptera: Formicidae) At Gopher Tortoise (Testudines: Testudinidae) Burrows. Fla. Entomol. 2005, 88, 349–354. [Google Scholar] [CrossRef]
- Allen, C.R.; Epperson, D.M.; Garmestani, A.S. Red Imported Fire Ant Impacts on Wildlife: A Decade of Research. Am. Midl. Nat. 2004, 152, 88–103. [Google Scholar] [CrossRef] [Green Version]
- Stoker, R.L.; Grant, W.E.; Vinson, S.B. Solenopsis invicta (Hymenoptera: Formicidae) effect on invertebrate decomposers of carrion in central Texas. Environ. Entomol. 1995, 24, 817–822. [Google Scholar] [CrossRef]
- Vinson, S.B. Impact of the invasion of the imported fire ant. Insect Sci. 2013, 20, 439–455. [Google Scholar] [CrossRef]
- Wojcik, D.P.; Allen, C.R.; Brenner, R.J.; Forys, E.A.; Jouvenaz, D.P.; Lutz, R.S. Red Imported Fire Ants: Impact on Biodiversity. Am. Entomol. 2001, 47, 16–23. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Hubbard, H.G. Additional Notes on the insect guests of the Florida land tortoise. Proc. Entomol. Soc. Wash. 1896, 3, 299–302. [Google Scholar]
- Cox, J.; Inkley, D.; Kautz, R. Ecology and Protection needs of Gopher Tortoise (Gopherus polyphemus) Populations Found on Lands Slated for Large-Scale Development in Florida; Nongame Wildlife Program Technical Report No. 4; Florida Game and Fresh Water Fish Commission: Tallahassee, FL, USA, 1987. [Google Scholar]
- Calvert, W.H. Fire ant predation on monarch larvae (Nymphalidae: Danainae) in a central Texas prairie. J. Lepid. Soc. 1996, 50, 149–151. [Google Scholar]
- Calvert, W.H. Patterns in the spatial and temporal use of the Texas milkweeds (Asclepiadaceae) by the monarch butterfly (Danus plexippus L.) during fall, 1996. J. Lepid. Soc. 1999, 53, 37–44. [Google Scholar]
- Forys, E.A.; Quistorff, A.; Allen, C.R. Potential Fire Ant (Hymenoptera: Formicidae) Impact on the Endangered Schaus Swallowtail. Fla. Entomol. 2001, 84, 254–258. [Google Scholar] [CrossRef]
- Morrison, L.W. Long-term impacts of an arthropod-community invasion by the imported fire ant, Solenopsis invicta. Ecology 2002, 83, 2337–2345. [Google Scholar] [CrossRef]
- Dziadzio, M.C.; Smith, L.L. Vertebrate Use of Gopher Tortoise Burrows and Aprons. Southeast. Nat. 2016, 15, 586–594. [Google Scholar] [CrossRef]
- White, K.N.; Tuberville, T.D. Birds and Burrows: Avifauna Use and Visitation of Burrows of Gopher Tortoises at Two Military Sites in the Florida Panhandle. Wilson J. Ornithol. 2017, 129, 792–803. [Google Scholar] [CrossRef]
- Richter, S.C.; Jackson, J.A.; Hinderliter, M.; Epperson, D.; Theodorakis, C.W.; Adams, S.M. Conservation genetics of the largest cluster of federally threatened gopher tortoise (gopherus polyphemus) colonies with implications for species management. Herpetologica 2011, 67, 406–419. [Google Scholar] [CrossRef]
- Collins, C.D.; Banks-Leite, C.; Brudvig, L.A.; Foster, B.L.; Cook, W.M.; Damschen, E.I.; Andrade, A.; Austin, M.; Camargo, J.L.; Driscoll, D.A.; et al. Fragmentation affects plant community composition over time. Ecography 2017, 40, 119–130. [Google Scholar] [CrossRef]








| Class | Order | Family | Genus | Species | Treatment |
|---|---|---|---|---|---|
| CHILOPODA | Both | ||||
| DIPOPLODA | Both | ||||
| ARACHNIDA | Araneae | Agelenidae | Agelenopsis | naevia | Treated |
| Agelenopsis | Both | ||||
| Coras | Untreated | ||||
| Neoscona | . | Untreated | |||
| Tegeraria | domestica | Treated | |||
| Araneidae | Araneus | . | Treated | ||
| Genus A | Untreated | ||||
| Argiope | aurantia | Treated | |||
| Gnaphosidae | Gnaphosa | . | Both | ||
| Hahniidae | Genus A | Untreated | |||
| Linphiidae | Genus A | Treated | |||
| Lycosidae | Lycosa | helluo | Both | ||
| Lycosa | avida | Treated | |||
| Lycosa | rabida | Treated | |||
| Lycosa | . | Untreated | |||
| Schizocosa | Untreated | ||||
| Sossipus | . | Both | |||
| Mimetidae | Mimetus | Treated | |||
| Philodromidae | Apollophanes | Treated | |||
| Salticidae | Evarcha | Untreated | |||
| Phidippus | Treated | ||||
| Opiliones | Phalangidae | Genus A | Treated | ||
| INSECTA | Blattaria | Blatellidae | Genus A | Treated | |
| Coleoptera | Scarabaeidae | Aphodius | Both | ||
| Staphylinidae | Genus A | Both | |||
| Carabidae | Agonum | Treated | |||
| Clivinia | Untreated | ||||
| Curclionidae | Pantomorus | cervinus | Untreated | ||
| Nitidulidae | Genus A | Untreated | |||
| Elateridae | Genus A | Treated | |||
| Diptera | Anthomyidae | Eutrichota | gopheri * | Both | |
| Chironomidae | Genus A | Untreated | |||
| Culicidae | Genus A | Both | |||
| Dolichopodidae | Hercostoma | Both | |||
| Hercostoma | Both | ||||
| Sphaeroceridae | Genus A | Both | |||
| Hemiptera | Lygaeidae | Genus A | Untreated | ||
| Miridae | Genus A | Treated | |||
| Pentatomidae | Thylantea | calceata | Untreated | ||
| Cicadellidae | Genus A | Both | |||
| Genus B | Treated | ||||
| Hymenoptera | Braconidae | Genus A | Untreated | ||
| Formicidae | Aphenogaster | carolinense | Treated | ||
| Aphenogaster | lamellidans | Both | |||
| Aphenogaster | Both | ||||
| Brachymyrmex | depilis | Treated | |||
| Cyphomyrmex | rimosus | Both | |||
| Dorymyrmex | bureni | Both | |||
| Dorymyrmex | medeis | Both | |||
| Paratrechina | Both | ||||
| Solenopsis | invicta | Both | |||
| Halictidae | Genus A | Treated | |||
| Mutillidae | Dasymutilla | Treated | |||
| Both | |||||
| Lepidoptera | Treated | ||||
| Orthoptera | Gryllacridae | Ceutophilis | divergens* | Both | |
| Gryllidae | Gryllus | Treated | |||
| Tettigoniidae | Both |
| Scientific Name | Treatment |
|---|---|
| Aphenogaster near rudis | both |
| Brachymyrmex depilis | treated |
| Dorymyrmex bureni | both |
| Paratrechina concinna | untreated |
| Paratrechina longicornis | both |
| Paratrechina phantasma | untreated |
| Pheidole dentata | treated |
| Pheidole dentigula | untreated |
| Pheidole floridana | both |
| Pheidole metallescens | both |
| Pheidole moerens | both |
| Solenopsis sp. | untreated |
| Solenopsis invicta | both |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Epperson, D.M.; Allen, C.R.; Hogan, K.F.E. Red Imported Fire Ants Reduce Invertebrate Abundance, Richness, and Diversity in Gopher Tortoise Burrows. Diversity 2021, 13, 7. https://doi.org/10.3390/d13010007
Epperson DM, Allen CR, Hogan KFE. Red Imported Fire Ants Reduce Invertebrate Abundance, Richness, and Diversity in Gopher Tortoise Burrows. Diversity. 2021; 13(1):7. https://doi.org/10.3390/d13010007
Chicago/Turabian StyleEpperson, Deborah M., Craig R. Allen, and Katharine F. E. Hogan. 2021. "Red Imported Fire Ants Reduce Invertebrate Abundance, Richness, and Diversity in Gopher Tortoise Burrows" Diversity 13, no. 1: 7. https://doi.org/10.3390/d13010007
APA StyleEpperson, D. M., Allen, C. R., & Hogan, K. F. E. (2021). Red Imported Fire Ants Reduce Invertebrate Abundance, Richness, and Diversity in Gopher Tortoise Burrows. Diversity, 13(1), 7. https://doi.org/10.3390/d13010007