
Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Methods
2.1. Study Area and Fieldwork
2.2. Molecular Analyses
2.3. Phylogenetic and Statistical Analysis
3. Results
3.1. Prevalences of Haemosporidian Parasites
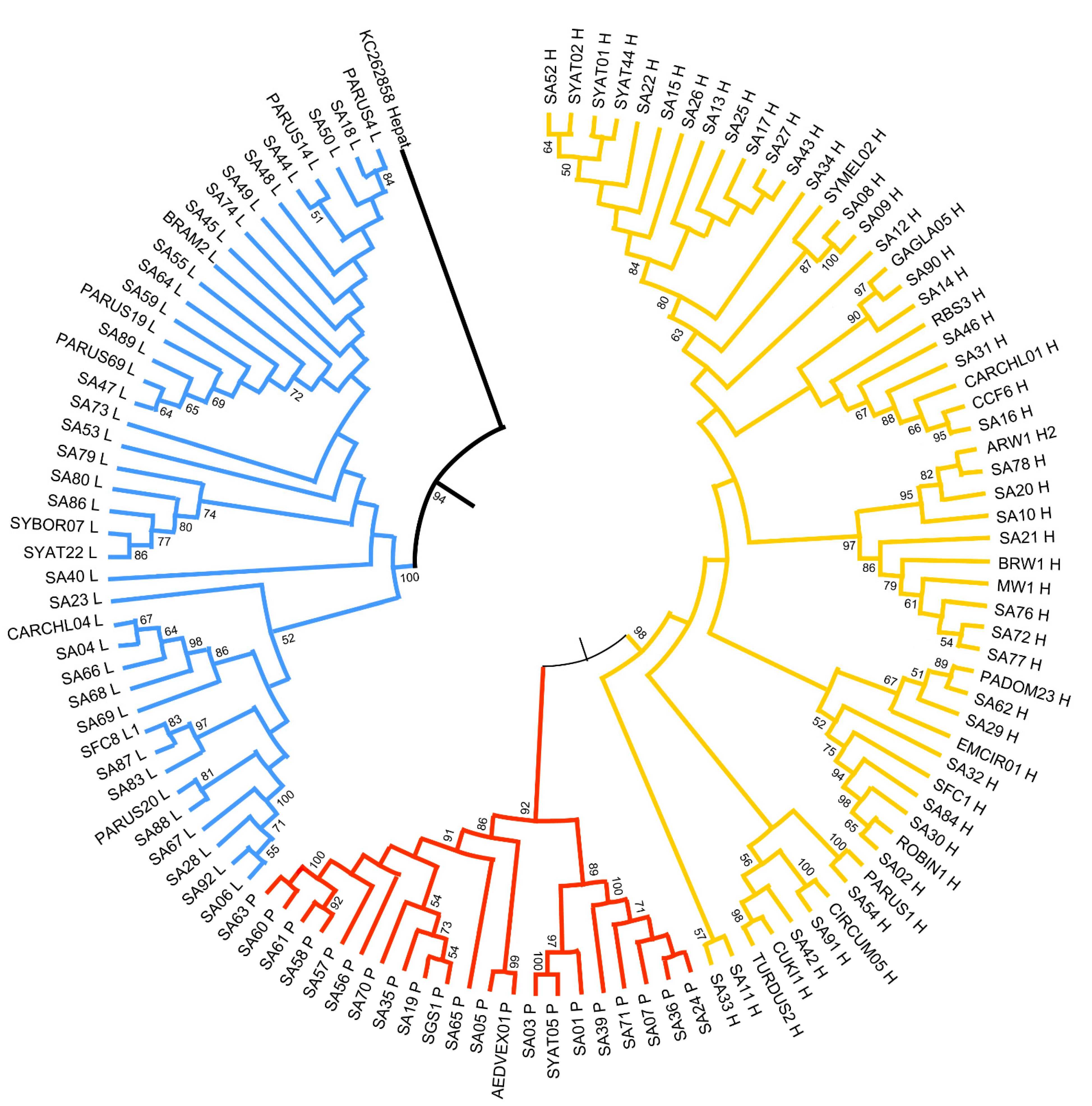
3.2. Genetic Diversity and Phylogeny of Haemosporidian Lineages
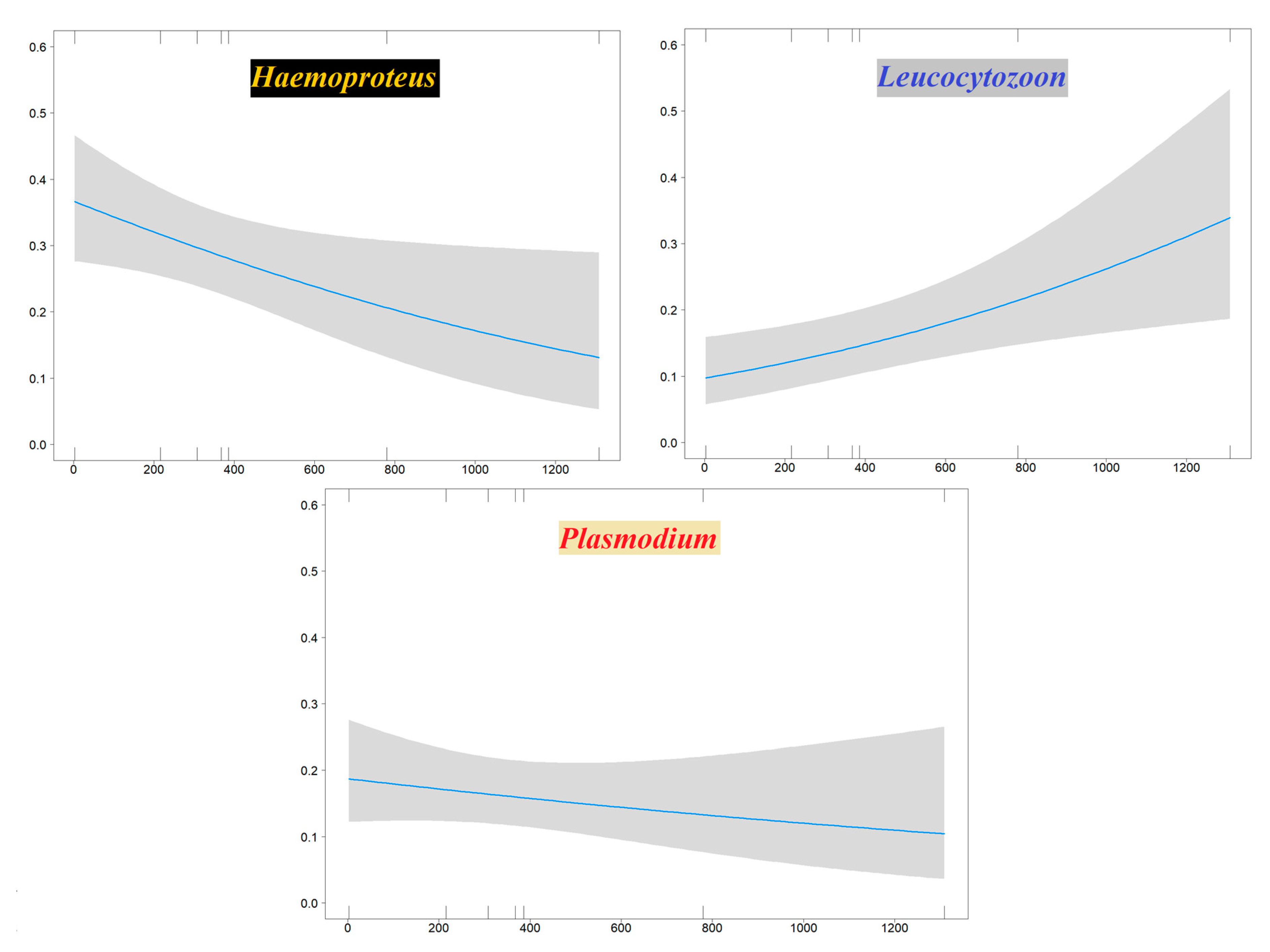
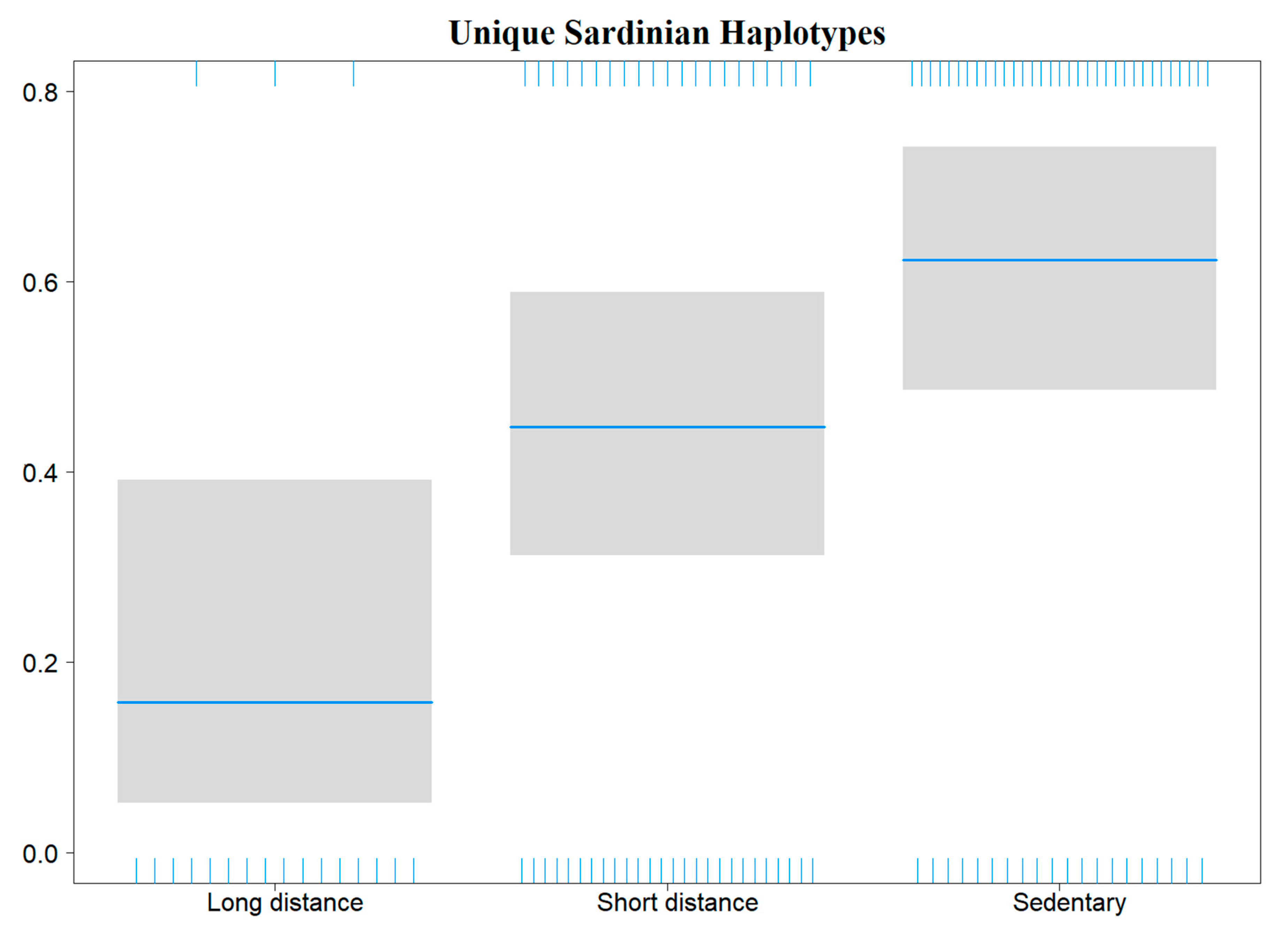
3.3. Haemosporidian Occurrence Predictors
4. Discussion
4.1. Sardinian Haemosporidian Diversity
4.2. Lineage Distributions
4.3. Genetic Diversity
4.4. Effect of Elevation and Host Species Migratory Pattern
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, K.; Träff, J.; Gustafsson, L. Importance of infection of haemosporidia blood parasites during different life history stages for long-term reproductive fitness of collared flycatchers. J. Avian Biol. 2019, 50, 1–11. [Google Scholar] [CrossRef]
- Schoener, E.R.; Banda, M.; Howe, L.; Castro, I.C.; Alley, M.R. Avian malaria in New Zealand. N. Z. Vet. J. 2014, 62, 189–198. [Google Scholar] [CrossRef]
- Van Riper, C., III; van Riper, S.G.; Lee Goff, M.; Laird, M. The Epizootiology and Ecological Significance of Malaria in Hawaiian Land Birds. Ecol. Monogr. 1986, 56, 327–344. [Google Scholar] [CrossRef]
- Magallanes, S.; García-Longoria, L.; López-Calderón, C.; Reviriego, M.; de Lope, F.; Møller, A.P.; Marzal, A. Uropygial gland volume and malaria infection are related to survival in migratory house martins. J. Avian Biol. 2017, 48, 1355–1359. [Google Scholar] [CrossRef]
- Bensch, S.; Hellgren, O.; PÉrez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Santiago-Alarcon, D.; Palinauskas, V.; Schaefer, H.M. Diptera vectors of avian Haemosporidian parasites: Untangling parasite life cycles and their taxonomy. Biol. Rev. 2012, 87, 928–964. [Google Scholar] [CrossRef] [PubMed]
- Drovetski, S.V.; Aghayan, S.A.; Mata, V.A.; Lopes, R.J.; Mode, N.A.; Harvey, J.A.; Voelker, G. Does the niche breadth or trade-off hypothesis explain the abundance-occupancy relationship in avian Haemosporidia? Mol. Ecol. 2014, 23, 3322–3329. [Google Scholar] [CrossRef]
- Harrigan, R.J.; Sedano, R.; Chasar, A.C.; Chaves, J.A.; Nguyen, J.T.; Whitaker, A.; Smith, T.B. New host and lineage diversity of avian haemosporidia in the northern Andes. Evol. Appl. 2014, 7, 799–811. [Google Scholar] [CrossRef] [Green Version]
- Zehtindjiev, P.; Križanauskienė, A.; Bensch, S.; Palinauskas, V.; Asghar, M.; Dimitrov, D.; Scebba, S.; Valkiūnas, G. A New Morphologically Distinct Avian Malaria Parasite That Fails Detection by Established Polymerase Chain Reaction–Based Protocols for Amplification of the Cytochrome B Gene. J. Parasitol. 2012, 98, 657–665. [Google Scholar] [CrossRef]
- Scaglione, F.E.; Cannizzo, F.T.; Chiappino, L.; Sereno, A.; Ripepi, M.; Salamida, S.; Manuali, E.; Bollo, E. Plasmodium spp. In a captive raptor collection of a safaripark in northwest Italy. Res. Vet. Sci. 2016, 104, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Fecchio, A.; Pinheiro, R.; Felix, G.; Faria, I.P.; Pinho, J.B.; Lacorte, G.A.; Braga, E.M.; Farias, I.P.; Aleixo, A.; Tkach, V.V.; et al. Host community similarity and geography shape the diversity and distribution of haemosporidian parasites in Amazonian birds. Ecography 2018, 41, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E.; Swanson, B.L.; Fallon, S.M.; Martínez-Abraín, A.; Scheuerlein, A.; Gray, J.; Latta, S.C. Community relationships of avian malaria parasites in southern Missouri. Ecol. Monogr. 2005, 75, 543–559. [Google Scholar] [CrossRef]
- Liu, B.; Deng, Z.; Huang, W.; Dong, L.; Zhang, Y. High prevalence and narrow host range of haemosporidian parasites in Godlewski’s bunting (Emberiza godlewskii) in northern China. Parasitol. Int. 2019, 69, 121–125. [Google Scholar] [CrossRef]
- González, A.D.; Matta, N.E.; Ellis, V.A.; Miller, E.T.; Ricklefs, R.E.; Gutiérrez, H.R. Mixed species flock, nest height, and elevation partially explain avian haemoparasite prevalence in Colombia. PLoS ONE 2014, 9, e100695. [Google Scholar] [CrossRef] [Green Version]
- Fecchio, A.; Ellis, V.A.; Bell, J.A.; Andretti, C.B.; D’horta, F.M.; Silva, A.M.; Tkach, V.V.; Weckstein, J.D. Avian malaria, ecological host traits and mosquito abundance in southeastern Amazonia. Parasitology 2017, 144, 1117–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, J.A.; Voelker, G. Host associations and climate influence avian haemosporidian distributions in Benin. Int. J. Parasitol. 2019, 49, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Clark, N.J.; Drovetski, S.V.; Voelker, G. Robust geographical determinants of infection prevalence and a contrasting latitudinal diversity gradient for haemosporidian parasites in Western Palearctic birds. Mol. Ecol. 2020, 29, 3131–3143. [Google Scholar] [CrossRef] [PubMed]
- Bodawatta, K.H.; Synek, P.; Bos, N.; Garcia-del-Rey, E.; Koane, B.; Marki, P.Z.; Albrecht, T.; Lifjeld, J.; Poulsen, M.; Munclinger, P.; et al. Spatiotemporal patterns of avian host–parasite interactions in the face of biogeographical range expansions. Mol. Ecol. 2020, 29, 2431–2448. [Google Scholar] [CrossRef]
- Clark, N.J.; Clegg, S.M.; Lima, M.R. A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): New insights from molecular data. Int. J. Parasitol. 2014, 44, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Illera, J.C.; López, G.; García-Padilla, L.; Moreno, Á. Factors governing the prevalence and richness of avian haemosporidian communities within and between temperate mountains. PLoS ONE 2017, 12, e0184587. [Google Scholar] [CrossRef]
- Ferraguti, M.; de la Puente, J.M.; Bensch, S.; Roiz, D.; Ruiz, S.; Viana, D.S.; Soriguer, R.C.; Figuerola, J. Ecological determinants of avian malaria infections: An integrative analysis at landscape, mosquito and vertebrate community levels. J. Anim. Ecol. 2018, 87. [Google Scholar] [CrossRef]
- Garcia-Longoria, L.; Marzal, A.; De Lope, F.; Garamszegi, L. Host-parasite interaction explains variation in the prevalence of avian haemosporidians at the community level. PLoS ONE 2019, 14, e0205624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanković, D.; Jönsson, J.; Raković, M. Diversity of avian blood parasites in wild passerines in Serbia with special reference to two new lineages. J. Ornithol. 2019, 160, 545–555. [Google Scholar] [CrossRef]
- De La Puente, J.M.; Muñoz, J.; Capelli, G.; Montarsi, F.; Soriguer, R.; Arnoldi, D.; Rizzoli, A.; Figuerola, J. Avian malaria parasites in the last supper: Identifying encounters between parasites and the invasive Asian mosquito tiger and native mosquito species in Italy. Malar. J. 2015, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Romano, A.; Nodari, R.; Bandi, C.; Caprioli, M.; Costanzo, A.; Ambrosini, R.; Rubolini, D.; Parolini, M.; Epis, S.; Saino, N. Haemosporidian parasites depress breeding success and plumage coloration in female barn swallows Hirundo rustica. J. Avian Biol. 2019, 50, 1–14. [Google Scholar] [CrossRef]
- Gutiérrez-López, R.; Gangoso, L.; De La Puente, J.M.; Fric, J.; López-López, P.; Mailleux, M.; Muñoz, J.; Touati, L.; Samraoui, B.; Figuerola, J. Low prevalence of blood parasites in a long-distance migratory raptor: The importance of host habitat. Parasites Vectors 2015, 8, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Hellgren, O.; Wood, M.J.; Waldenström, J.; Hasselquist, D.; Ottosson, U.; Stervander, M.; Bensch, S. Circannual variation in blood parasitism in a sub-Saharan migrant passerine bird, the garden warbler. J. Evol. Biol. 2013, 26, 1047–1059. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Grussu, M. Checklist of the birds of Sardinia. Aves Ichnusae 2001, 4, 2–55. [Google Scholar]
- Pellegrino, I.; Negri, A.; Boano, G.; Cucco, M.; Kristensen, T.; Pertoldi, C.; Randi, E.; Salek, M.; Mucci, N. Evidence for strong genetic structure in European populations of the little owl Athene noctua. J. Avian Biol. 2015, 46, 462–1475. [Google Scholar] [CrossRef]
- Pellegrino, I.; Boatti, L.; Cucco, M.; Mignone, F.; Kristensen, T.N.; Mucci, N.; Randi, E.; Ruiz-Gonzalez, A.; Pertoldi, C. Development of SNP markers for population structure and phylogeography characterization in little owl (Athene noctua) using a genotyping- by-sequencing approach. Conserv. Genet. Resour. 2016, 8, 13. [Google Scholar] [CrossRef]
- Pons, J.-M.; Thibault, J.-C.; Aymí, R.; Grussu, M.; Muntaner, J.; Olioso, G.; Sunyer, J.R.; Touihri, M.; Fuchs, J. The role of western Mediterranean islands in the evolutionary diversification of the spotted flycatcher Muscicapa striata, a long-distance migratory passerine species. J. Avian Biol. 2016, 47, 386–398. [Google Scholar] [CrossRef]
- Zuccon, D.; Pons, J.; Boano, G.; Chiozzi, G.; Gamauf, A.; Mengoni, C.; Nespoli, D.; Olioso, G.; Pavia, M.; Pellegrino, I.; et al. Type specimens matter: New insights on the systematics, taxonomy and nomenclature of the subalpine warbler (Sylvia cantillans) complex. Zool. J. Linn. Soc. 2020, 169, 1–28. [Google Scholar] [CrossRef]
- Cucco, M.; Grenna, M.; Pellegrino, I.; Malacarne, G. Egg-sequence rather than mating preference influences female egg investment in the red-legged partridge. Ethol. Ecol. Evol. 2011, 23, 343–357. [Google Scholar] [CrossRef]
- Seutin, G.; White, B.N.; Boag, P.T. Preservation of avian blood and tissue samples for DNA analysis. Can. J. Zool. 1991, 69, 82–90. [Google Scholar] [CrossRef]
- Gill, F.; Donsker, D.; Rasmussen, P. (Eds.) IOC World Bird List (v11.1). 2021. Available online: https://www.worldbirdnames.org/ioc-lists/crossref/ (accessed on 29 September 2020). [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Browning, S.R.; Browning, B.L. Haplotype phasing: Existing methods and new developments. Nat. Rev. Genet. 2011, 12, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M.; Smith, N.; Donnelly, P. A new statistical method for haplotype reconstruction from population data. Am. J. Hum. Gen. 2001, 68, 978-–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 29 October 2020).
- Breheny, P.; Burchett, W. Visualization of regression models using Visreg. R J. 2017, 9, 56–71. [Google Scholar] [CrossRef]
- Gangoso, L.; Gutiérrez-López, R.; Martínez-de la Puente, J.; Figuerola, J. Louse flies of Eleonora’s falcons that also feed on their prey are evolutionary dead-end hosts for blood parasites. Mol. Ecol. 2019, 1812–1825. [Google Scholar] [CrossRef] [Green Version]
- Himmel, T.; Harl, J.; Pfanner, S.; Nedorost, N.; Nowotny, N.; Weissenböck, H. Haemosporidioses in wild Eurasian blackbirds (Turdus merula) and song thrushes (T. philomelos): An in situ hybridization study with emphasis on exo-erythrocytic parasite burden. Malar. J. 2020, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mata, V.A.; da Silva, L.P.; Lopes, R.J.; Drovetski, S.V. The Strait of Gibraltar poses an effective barrier to host-specialised but not to host-generalised lineages of avian Haemosporidia. Int. J. Parasitol. 2015, 45, 711–719. [Google Scholar] [CrossRef]
- Fernández, M.; Rojo, M.Á.; Casanueva, P.; Carrión, S.; Hernández, M.Á.; Campos, F. High prevalence of haemosporidians in Reed Warbler Acrocephalus scirpaceus and Sedge Warbler Acrocephalus schoenobaenus in Spain. J. Ornithol. 2010, 151, 27–32. [Google Scholar] [CrossRef]
- Ventim, R.; Tenreiro, P.; Grade, N.; Encarnação, P.; Araújo, M.; Mendes, L.; Pérez-Tris, J.; Ramos, J.A. Characterization of haemosporidian infections in warblers and sparrows at south-western European reed beds. J. Ornithol. 2012, 153, 505–512. [Google Scholar] [CrossRef]
- Ciloglu, A.; Ergen, A.G.; Inci, A.; Dik, B.; Duzlu, O.; Onder, Z.; Yetismis, G.; Bensch, S.; Valkiūnas, G.; Yildirim, A. Prevalence and genetic diversity of avian haemosporidian parasites at an intersection point of bird migration routes: Sultan Marshes National Park, Turkey. Acta Trop. 2020, 210. [Google Scholar] [CrossRef] [PubMed]
- Jovani, R.; Tella, J.L. Parasite prevalence and sample size: Misconceptions and solutions. Trends Parasitol. 2006, 22, 214–218. [Google Scholar] [CrossRef]
- Illera, J.C.; Fernández-Álvarez, Á.; Hernández-Flores, C.N.; Foronda, P. Unforeseen biogeographical patterns in a multiple parasite system in Macaronesia. J. Biogeogr. 2015, 42, 1858–1870. [Google Scholar] [CrossRef]
- Ellis, V.A.; Huang, X.; Westerdahl, H.; Jönsson, J.; Hasselquist, D.; Neto, J.M.; Nilsson, J.Å.; Nilsson, J.; Hegemann, A.; Hellgren, O.; et al. Explaining prevalence, diversity and host specificity in a community of avian haemosporidian parasites. Oikos 2020, 129, 1314–1329. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, A.; Ramírez, Á.; Richardson, D.S.; Pérez-Tris, J. Evolution of parasite island syndromes without long-term host population isolation: Parasite dynamics in Macaronesian blackcaps Sylvia atricapilla. Glob. Ecol. Biogeogr. 2013, 22, 1272–1281. [Google Scholar] [CrossRef]
- Dimitrov, D.; Zehtindjiev, P.; Bensch, S. Genetic diversity of avian blood parasites in SE Europe: Cytochrome b lineages of the genera Plasmodium and Haemoproteus (Haemosporida) from Bulgaria. Acta Parasitol. 2010, 55, 201–209. [Google Scholar] [CrossRef]
- Hellgren, O.; Waldenström, J.; Peréz-Tris, J.; Szöll Ösi, E.; Hasselquist, D.; Krizanauskiene, A.; Ottosson, U.; Bensch, S. Detecting shifts of transmission areas in avian blood parasites—A phylogenetic approach. Mol. Ecol. 2007, 16, 1281–1290. [Google Scholar] [CrossRef]
- Hellgren, O.; Kutzer, M.; Bensch, S.; Valkiūnas, G.; Palinauskas, V. Identification and characterization of the merozoite surface protein 1 (msp1) gene in a host-generalist avian malaria parasite, Plasmodium relictum (lineages SGS1 and GRW4) with the use of blood transcriptome. Malar. J. 2013, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- López, G.; Muñoz, J.; Soriguer, R.; Figuerola, J. Increased Endoparasite Infection in Late-Arriving Individuals of a Trans-Saharan Passerine Migrant Bird. PLoS ONE 2013, 8, e61236. [Google Scholar] [CrossRef] [Green Version]
- Ayadi, T.; Selmi, S.; Hammouda, A.; Reis, S.; Boulinier, T.; Loiseau, C. Diversity, prevalence and host specificity of avian parasites in southern Tunisian oases. Parasitology 2018, 145, 971–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, M.A.; Matta, N.E.; Valkiünas, G.; Parker, P.G.; Mello, B.; Stanley, C.E.; Lentino, M.; Garcia-Amado, M.A.; Cranfield, M.; Pond, S.L.K.; et al. Mode and rate of evolution of haemosporidian mitochondrial genomes: Timing the radiation of avian parasites. Mol. Biol. Evol. 2018, 35, 383–403. [Google Scholar] [CrossRef] [Green Version]
- Outlaw, D.C.; Ricklefs, R.E. Comparative Gene Evolution in Haemosporidian (Apicomplexa) Parasites of Birds and Mammals. Mol. Biol. Evol. 2010, 27, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Grill, A.; Casula, P.; Lecis, R.; Menken, S. Endemism in Sardinia. In Phylogeography of Southern European Refugia: Evolutionary Perspectives on the Origins and Conservation of European Biodiversity; Weiss, S., Ferrand, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 273–296. ISBN 978-1-4020-4904-0. [Google Scholar]
- Gupta, P.; Vishnudas, C.K.; Ramakrishnan, U.; Robin, V.V.; Dharmarajan, G. Geographical and host species barriers differentially affect generalist and specialist parasite community structure in a tropical sky-island archipelago. Proc. R. Soc. B Biol. Sci. 2019, 286. [Google Scholar] [CrossRef]
- Hsü, K.J.; Montadert, L.; Bernouilli, D.; Cita, M.B.; Erikson, A.; Garrison, R.E.; Kidd, R.B.; Melieres, F.; Muller, C.; Wright, R. History of the Mediterranean salinity crisis. Nature 1978, 267, 399–403. [Google Scholar] [CrossRef]
- Tognotti, E. Program to eradicate malaria in Sardinia, 1946–1950. Emerg. Infect. Dis. 2009, 15, 1460–1466. [Google Scholar] [CrossRef] [PubMed]
- Hocking, B.; Richards, W. Biology and Control of Labrador Black Flies (Diptera: Simuliidae). Bull. Entomol. Res. 1952, 43, 237–257. [Google Scholar] [CrossRef]
- Imamura, H.; Downing, T.; van den Broeck, F.; Sanders, M.J.; Rijal, S.; Sundar, S.; Mannaert, A.; Vanaerschot, M.; Berg, M.; de Muylder, G.; et al. Evolutionary genomics of epidemic visceral leishmaniasis in the Indian subcontinent. Elife 2016, 5, 1–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, S.; Melo, M.; Covas, R.; Doutrelant, C.; Pereira, H.; de Lima, R.; Loiseau, C. Influence of land use and host species on parasite richness, prevalence and co-infection patterns. Int. J. Parasitol. 2021, 51, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Talbalaghi, A.; Pessino, M.; Agosta, P.; Bo, T.; Ignjatović-Ćupina, A. Overview of Possible Simuliidae Related Problems in the Alessandria District (Piedmont, Italy). Suppl. Acta Entomol. Serbica 2006, 77–81. [Google Scholar]
- Margoni, S.; Maiolini, B. Distribuzione altitudinale di Simulidi in torrenti alpini d’alta quota (Trentino, Italia). Stud. Trent. Sci. Nat. Acta Biol. 2003, 80, 63–64. [Google Scholar]
- Ya’Cob, Z.; Takaoka, H.; Pramual, P.; Low, V.L.; Sofian-Azirun, M. Distribution pattern of black fly (Diptera: Simuliidae) assemblages along an altitudinal gradient in Peninsular Malaysia. Parasites Vectors 2016, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Van Rooyen, J.; Lalubin, F.; Glaizot, O.; Christe, P. Avian haemosporidian persistence and co-infection in great tits at the individual level. Malar. J. 2013, 12, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latta, S.C.; Ricklefs, R.E. Prevalence patterns of avian haemosporida on hispaniola. J. Avian Biol. 2010, 41, 25–33. [Google Scholar] [CrossRef]
- Doussang, D.; González-Acuña, D.; Torres-Fuentes, L.G.; Lougheed, S.C.; Clemente-Carvalho, R.B.; Greene, K.C.; Vianna, J.A. Spatial distribution, prevalence and diversity of haemosporidians in the rufous-collared sparrow, Zonotrichia capensis. Parasites Vectors 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Mukhopadhyay, E.; Hazra, S.; Banerjee, D. Effect of Environment and Elevation on Seasonal Prevalence of Culicoides in West Bengal. Legum. Res. Int. J. 2020, 54, 1125–1135. [Google Scholar]
- Tschuor, A.C.; Kaufmann, C.; Schaffner, F.; Mathis, A. Vorkommen von gnitzen (Culicoides spp.) in drei höhenlagen einer alpinen region der schweiz. Schweiz. Arch. Tierheilkd. 2009, 151, 215–221. [Google Scholar] [CrossRef]
- Cosgrove, C.L.; Wood, M.J.; Day, K.P.; Sheldon, B.C. Seasonal variation in Plasmodium prevalence in a population of blue tits Cyanistes caeruleus. J. Anim. Ecol. 2008, 77, 540–548. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Migratory Pattern | n | prev. | Infections | Haplotypes Detected | ||||||
| Species | + | − | H | L | P | |||||
| Caprimulgidae | 1 | 0% | − | 1 | − | − | − | |||
| Caprimulgus europaeus | LD | 1 | 0% | − | 1 | − | − | − | ||
| Picidae | 4 | 0% | − | 4 | − | − | − | |||
| Dendrocopos major | S | 4 | 0% | − | 4 | − | − | − | ||
| Falconidae | 1 | 0% | − | 1 | − | − | − | |||
| Falco tinnunculus | S | 1 | 0% | − | 1 | − | − | − | ||
| Lanidae | 2 | 100% | 2 | − | 2 | − | − | |||
| Lanius senator | LD | 2 | 100% | 2 | − | 2 | − | − | RBS3_H | |
| Corvidae | 7 | 100% | 7 | − | 7 | − | − | |||
| Garrulus glandarius | S | 7 | 100% | 7 | − | 7 | − | − | CIRCUM05_H, GAGLA05_H, SA14_H, SA32_H, SA33_H, SA90_H, SA91_H | |
| Paridae | 29 | 55.2% | 16 | 13 | 3 | 10 | 9 | |||
| Periparus ater | S | 4 | 50% | 2 | 2 | − | 2 | 1 | PARUS20_L, SA88_L SA89_L, SGS1_P | |
| Cyanistes caeruleus | S | 13 | 38.5% | 5 | 8 | 2 | 5 | − | PARUS1_H, PARUS14_L, PARUS4_L, SA18_L, SA44_L, SA45_L, SA53_L, SA54_H | |
| Parus major | S | 12 | 75% | 9 | 3 | 1 | 3 | 8 | PARUS1_H, PARUS19_L, SA07_P, SA24_P, SA36_L, SA39_P, SA40_L, SA48_L, SA49_L, SA50_L, SA70_P, SA71_P, SA73_L, SA74_L, SGS1_P | |
| Alaudidae | 1 | 0% | − | 1 | − | − | 0 | |||
| Lullula arborea | SD | 1 | 0% | − | 1 | − | − | 0 | ||
| Hirundinidae | 1 | 0% | − | 1 | − | − | 0 | |||
| Hirundo rustica | LD | 1 | 0% | − | 1 | − | − | 0 | ||
| Cettiidae | 4 | 25% | 1 | 3 | − | − | 1 | |||
| Cettia cetti | S | 4 | 25% | 1 | 3 | − | − | 1 | SGS1_P | |
| Phylloscopidae | 1 | 0% | − | 1 | − | − | 0 | |||
| Phylloscopus collybita | SD | 1 | 0% | − | 1 | − | − | 0 | ||
| Acrocephalidae | 22 | 63.6% | 14 | 8 | 14 | − | − | |||
| Acrocephalus scirpaceus | LD | 22 | 63.6% | 14 | 8 | 14 | − | − | BRW1_H, ARW1_H, MW1_H, SA10_H, SA20_H, SA21_H, SA72_H, SA76_H, SA77_H, SA78_H | |
| Sylviidae | 39 | 59.0% | 23 | 16 | 16 | 5 | 3 | |||
| Sylvia atricapilla | SD | 15 | 86.6% | 13 | 2 | 11 | 2 | − | SA13_H, SA15_H, SA17_H, SA22_H, SA25_H, SA26_H, SA27_H, SA42_H, SA43_H, SA52_H, SA79_L, SA80_L, SA86_L, SA87_L, SYAT01_H, SYAT02_H, SYAT22_L, SYAT44_H, SYBOR07_L | |
| Curruca melanocephala | S | 15 | 53.3% | 8 | 7 | 5 | 1 | 3 | SYAT22_L, SYMEL02_H, SA08_H, SA09_H, SA34_H,SA35_P, SA83_L, SGS1_P | |
| Curruca subalpina | LD | 6 | 33.3% | 2 | 4 | − | 2 | − | SYAT22_L, SFC8_L | |
| Curruca sarda | SD | 3 | 0% | − | 3 | − | − | − | ||
| Regulidae | ||||||||||
| Regulus ignicapilla | SD | 2 | 0% | − | 2 | − | − | 2 | ||
| Sturnidae | 5 | 40% | 2 | 3 | − | − | 2 | |||
| Sturnus unicolor | S | 5 | 40% | 2 | 3 | − | − | 2 | SA03_P, SYAT05_P | |
| Turdidae | 15 | 73.3 | 11 | 4 | 2 | − | 9 | |||
| Turdus merula | SD | 14 | 71.4% | 10 | 4 | 1 | − | 9 | SA01_P, SYAT05_P, TURDUS2_H | |
| Turdus viscivorus | SD | 1 | 100% | 1 | − | 1 | − | − | CUKI1_H | |
| Muscicapidae | 22 | 54.5% | 12 | 10 | 10 | 1 | 2 | |||
| Muscicapa tyrrhenica | LD | 1 | 100% | 1 | − | 1 | − | − | SFC1_H | |
| Erithacus rubecula | SD | 16 | 62.5% | 10 | 6 | 9 | − | 2 | ROBIN1_H, SA02_H, SA19_P, SA30_H, SA84_H, SGS1_P | |
| Saxicola rubicola | SD | 5 | 20% | 1 | 4 | − | 1 | − | SFC8_L | |
| Passeridae | 27 | 44.4% | 12 | 15 | 2 | 6 | 7 | |||
| Passer hispaniolensis | S | 27 | 44.4% | 12 | 15 | 2 | 6 | 7 | PARUS69_L, PADOM23_H, SA06_L, SA28_L, SA47_L, SA55_L, SA56_P, SA57_P, SA58_P, SA59_L, SA60_P, SA61_P, SA62_H, SA63_P,SA64_L, SA65_P, SGS1_P | |
| Fringillidae | 20 | 70.0% | 14 | 6 | 6 | 8 | − | |||
| Fringilla coelebs | SD | 6 | 83.3% | 5 | 1 | 4 | 1 | − | BRAM2_L, CCF6_H, SA16_H, SA29_H, SA31_H, SA46_H | |
| Chloris chloris | SD | 3 | 66.7% | 2 | 1 | 1 | 1 | − | CARCHL01_H, CARCHL04_L | |
| Linaria cannabina | SD | 6 | 83.3% | 5 | 1 | − | 5 | − | CARCHL04_L, SA04_L, SA06_L, SA66_L, SA67_L,SA68_L, SA69_L, SA92_L | |
| Carduelis carduelis | SD | 1 | 0% | − | 1 | − | − | − | ||
| Carduelis corsicana | S | 4 | 50% | 2 | 2 | 1 | 1 | − | CARCHL04_L, SA11_H, | |
| Emberizidae | 13 | 38.5% | 5 | 8 | 2 | 2 | 2 | |||
| Emberiza calandra | S | 7 | 14.3% | 1 | 6 | − | − | 1 | SA05_P | |
| Emberiza cirlus | S | 6 | 66.7% | 4 | 2 | 2 | 2 | 1 | AEDVEX01_P, PARUS69_L, EMCIR01_H,, SA12_H, SA23_L | |
| Locality | Elevation m a.s.l. | Environment | n | prev. | H | L | P |
|---|---|---|---|---|---|---|---|
| Cabras | 3 m | Agricultural mosaic | 15 | 33.3% | 4 | - | 5 |
| Marceddì | 3 m | Reed-dominated Wetland | 33 | 51.5% | 23 | 2 | 2 |
| Luogosanto | 216 m | Woodland | 45 | 48.9% | 18 | 9 | 7 |
| Priatu | 308 m | Woodland | 52 | 69.2% | 30 | 9 | 13 |
| Telti | 370 m | Agricultural mosaic | 32 | 65.6% | 7 | 4 | 6 |
| Monte Albo | 780 m | Montane shrublands | 17 | 58.8% | 1 | 8 | 1 |
| Monte Limbara | 1309 m | Montane shrublands | 22 | 36.4% | 4 | 7 | 1 |
| Haemoproteus Occurrence | β ± ES | Z | p |
| intercept | −0.523 ± 0.211 | −2.477 | 0.013 |
| Sample Site Elevation | −0.001 ± 0.0005 | −2.141 | 0.032 |
| Leucocytozoon Occurrence | β ± ES | Z | p |
| intercept | −2.231 ± 0.288 | −7.741 | <0.001 |
| Sample Site Elevation | 0.001 ± 0.0004 | 2.713 | 0.007 |
| Plasmodium Occurrence | β ± ES | Z | p |
| intercept | −1.468 ± 0.258 | −5.689 | <0.001 |
| Sample Site Elevation | −0.001 ± 0.001 | −0.940 | 0.347 |
| NovelHaplotypes Occurrence | β ± ES | Z | p |
| Long−distance | −1.674 ± 0.629 | −2.661 | 0.008 |
| Short−distance | 1.460 ± 0.694 | 2.104 | 0.035 |
| Sedentary | 2.175 ± 0.690 | 3.152 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pellegrino, I.; Ilahiane, L.; Boano, G.; Cucco, M.; Pavia, M.; Prestridge, H.L.; Voelker, G. Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean. Diversity 2021, 13, 75. https://doi.org/10.3390/d13020075
Pellegrino I, Ilahiane L, Boano G, Cucco M, Pavia M, Prestridge HL, Voelker G. Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean. Diversity. 2021; 13(2):75. https://doi.org/10.3390/d13020075
Chicago/Turabian StylePellegrino, Irene, Luca Ilahiane, Giovanni Boano, Marco Cucco, Marco Pavia, Heather L. Prestridge, and Gary Voelker. 2021. "Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean" Diversity 13, no. 2: 75. https://doi.org/10.3390/d13020075
APA StylePellegrino, I., Ilahiane, L., Boano, G., Cucco, M., Pavia, M., Prestridge, H. L., & Voelker, G. (2021). Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean. Diversity, 13(2), 75. https://doi.org/10.3390/d13020075