
Study on the Relationship between Richness and Morphological Diversity of Higher Taxa in the Darkling Beetles (Coleoptera: Tenebrionidae)

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection and Arrangement
2.2. Richness Values and Morphological Diversity
3. Results
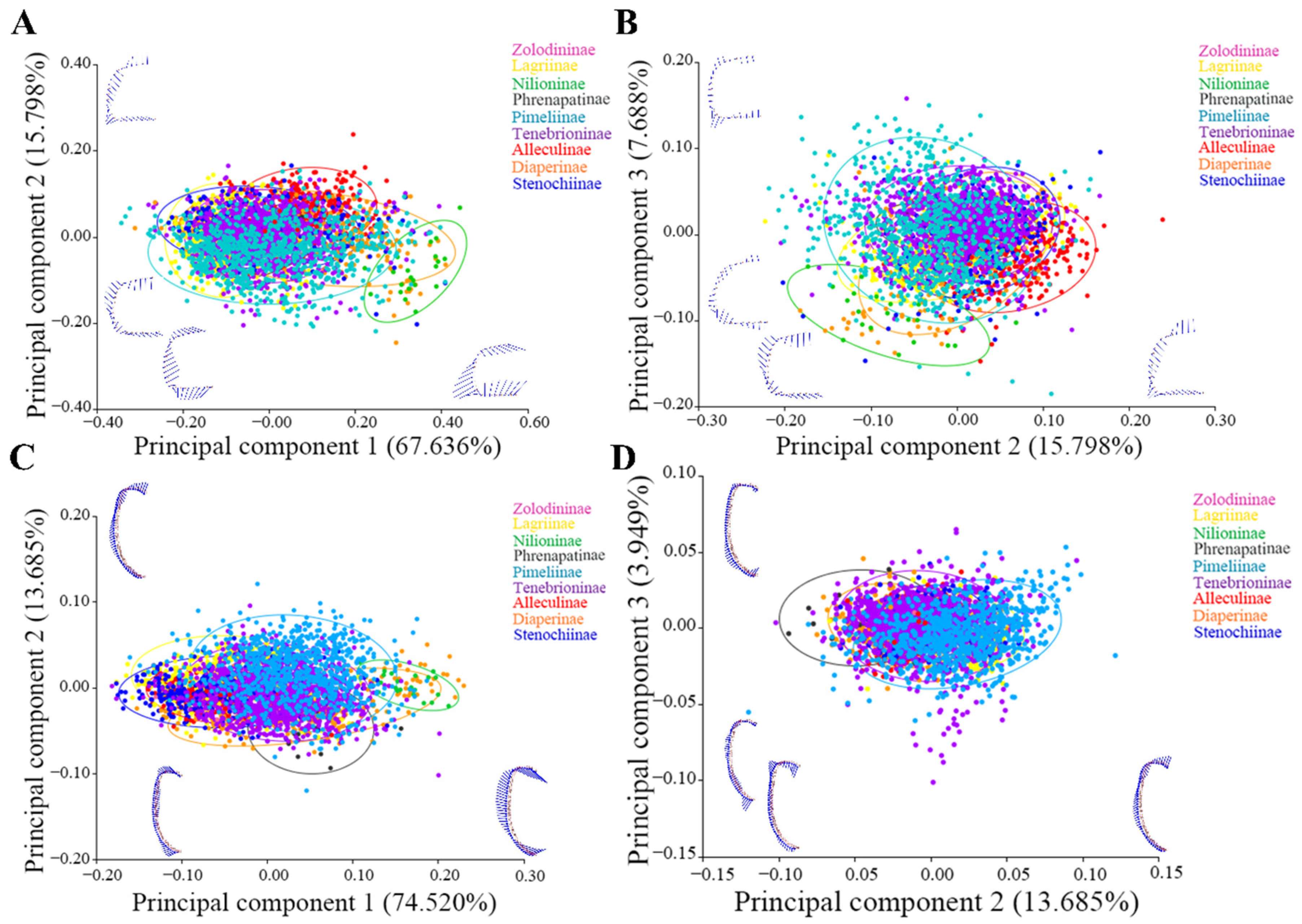
3.1. Morphological Variation of the Pronotum and Elytron
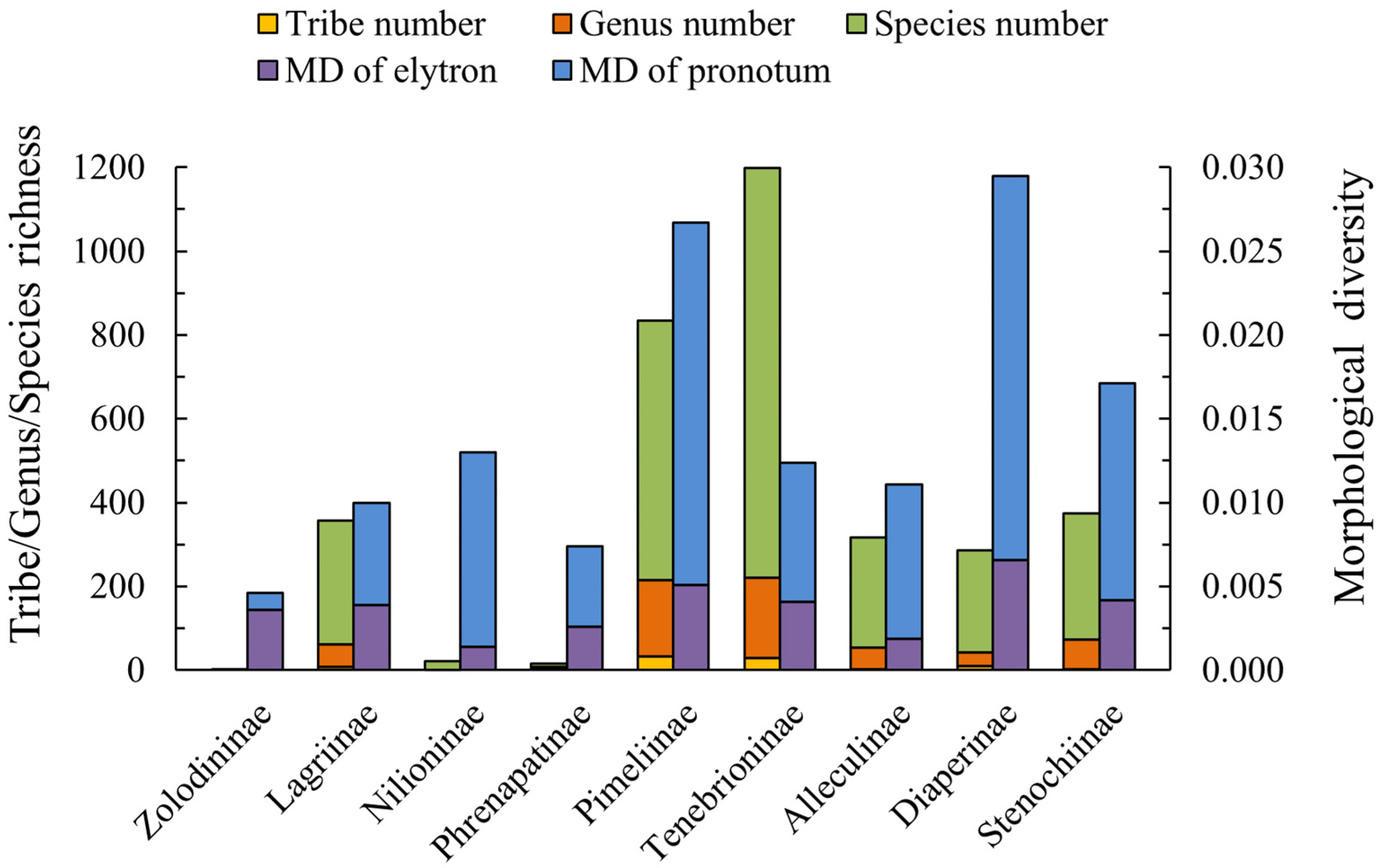
3.2. Taxa Richness and Morphological Diversity of Test Subfamilies
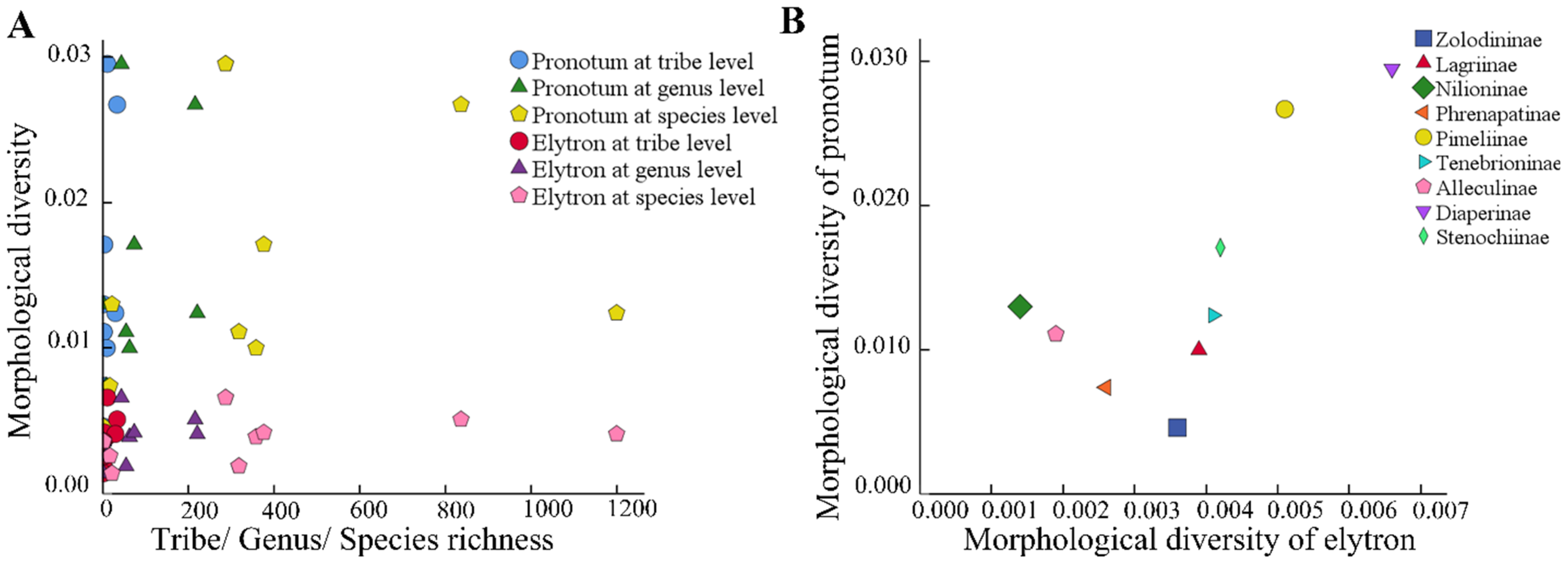
3.3. Correlation Analysis of Test Subfamilies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Z.; Colinet, G.; Zu, Y.; Wang, J.; An, L.; Li, Q.; Niu, X. Species diversity of Arabis alpina L. communities in two Pb/Zn mining areas with different smelting history in Yunnan Province, China. Chemosphere 2019, 233, 603–614. [Google Scholar] [CrossRef]
- Tong, Y.; Yang, H.; Yang, X.; Bai, M. The relationship between genus/species richness and morphological diversity among subfamilies of jewel beetles. Insects 2021, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Mohedano-Navarrete, A.; Reyes-Bonilla, H.; López-Pérez, R. Species richness and morphological diversity of the genus Porites in the Pacific Ocean. In Proceedings of the 11th International Coral Reef Symposium, Fort Lauderdale, FL, USA, 7–11 July 2008; pp. 7–11. [Google Scholar]
- Bell, J.; Barnes, D. Sponge morphological diversity: A qualitative predictor of species diversity? Aquat. Conserv. 2001, 11, 109–121. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Voultsiadou, E. Sponge diversity gradients in marine caves of the eastern Mediterranean. J. Mar. Biol. Assoc. UK 2015, 96, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.; Brandão, C. Ecosystem-wide morphological structure of leaf-litter ant communities along a tropical latitudinal gradient. PLoS ONE 2014, 9, e93049. [Google Scholar] [CrossRef]
- Neustupa, J.; Erná, K.; T’astn, J. Diversity and morphological disparity of desmid assemblages in Central European peatlands. Hydrobiologia 2008, 630, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Ruan, Y.; Wan, X.; Tong, Y.; Yang, X.; Bai, M. Geometric morphometric analysis of the pronotum and elytron in stag beetles: Insight into its diversity and evolution. ZooKeys 2019, 833, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Foote, M. Discordance and concordance between morphological and taxonomic diversity. Paleobiology 1993, 19, 185–204. [Google Scholar] [CrossRef] [Green Version]
- Mindel, B.; Neat, F.; Trueman, C.; Webb, T.; Blanchard, J. Functional, size and taxonomic diversity of fish along a depth gradient in the deep sea. Peer J 2016, 4, e2387. [Google Scholar] [CrossRef]
- Matthews, E.; Lawrence, J.; Bouchard, P.; Steiner, W.; Slipinski, A. 11.14. Tenebrionidae Latreille, 1802. In Volume 2 Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim); De Gruyter: Berlin, Germany, 2011; pp. 574–659. [Google Scholar]
- Yunlong, W. Species Diversity of Tenebrionidae (Coleoptera) in the North Luo-River Basin. Master’s Thesis, Shaanxi Normal University, Xi’an, China, 2008. [Google Scholar]
- Guodong, R.; Youzhi, Y. Tenebrionidae from the Arid and Semi-Arid Region of China; Hebei University Press: Baoding, China, 1999. [Google Scholar]
- Zelditch, M.L.; Fink, W.L.; Swiderski, D.L. Morphometrics, homology, and phylogenetics: Quantified characters as synapomorphies. Syst. Biol. 1995, 44, 179–189. [Google Scholar] [CrossRef]
- Banaszkiewicz, M. Comparative study of female genitalia in Pedinini (sensu Iwan 2004) (Coleoptera: Tenebrionidae: Pedinini), with notes on the classification. Ann. Zool. 2006, 56, 59–77. [Google Scholar]
- Kamiński, M.J.; Kanda, K.; Lumen, R.; Smith, A.; Iwan, D. Molecular phylogeny of Pedinini (Coleoptera, Tenebrionidae) and its implications for higher-level classification. Zool. J. Linn. Soc. 2019, 185, 77–97. [Google Scholar] [CrossRef]
- Juntong, L. Fauna Formation and Adaptive Evolution of the Darkling Beetles on Hainan Island. Master’s Thesis, Hebei University, Baoding, China, 2010. [Google Scholar]
- Ye, L. “Fake” Carabid beetle—Darkling beetle. Knowl. Is Power 2009, 2, 10–13. [Google Scholar]
- Fattorini, S. Conservation biogeography of Tenebrionid beetles: Insights from Italian Reserves. Diversity 2020, 12, 348. [Google Scholar] [CrossRef]
- Fattorini, S. Biogeography of the tenebrionid beetles (Coleoptera, Tenebrionidae) on the Aegean Islands (Greece). J. Biogeogr. 2002, 29, 49–67. [Google Scholar] [CrossRef]
- Chapman, R.F.; Simpson, S.J.; Douglas, A.E. The Insects: Structure and Function, 5th ed.; Cambridge University Press: Cambridge, UK, 2013; pp. 149–230. [Google Scholar]
- Frantsevich, L.; Dai, Z.; Wang, W.Y.; Zhang, Y. Geometry of elytra opening and closing in some beetles (Coleoptera, Polyphaga). J. Exp. Biol. 2005, 208, 3145–3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonk, J.; Shackelford, T. Encyclopedia of Animal Cognition and Behavior; Springer International Publishing: New York, NY, USA, 2019; pp. 920–926. [Google Scholar]
- Manseguiao, M.R.; Manseguiao, S.; Demayo, C. Morphological variations on the pronotum of the Hispid beetle P. reichei from eight Mindanao populations. Adv. Environ. Biol. 2015, 9, 46–55. [Google Scholar]
- Bunalski, M.; Ghahari, H. Contribution to the study of darkling beetles (Coleoptera: Tenebrionidae) from Golestan Province, Northern Iran. Wiadomości. Entomol. 2017, 36, 25–31. [Google Scholar]
- Fattorini, S.; Dapporto, L.; Strona, G.; Borges, P.A.V. Calling for a new strategy to measure environmental (habitat) diversity in Island Biogeography: A case study of Mediterranean tenebrionids (Coleoptera: Tenebrionidae). Fragm. Entomol. 2015, 47, 1–14. [Google Scholar] [CrossRef]
- Fattorini, S.; Monaco, R.L.; Giulio, A.D.; Ulrich, W. Latitudinal trends in body length distributions of European darkling beetles (Tenebrionidae). Acta Oecol. 2013, 53, 88–94. [Google Scholar] [CrossRef]
- Fattorini, S.; Ulrich, W. Drivers of species richness in European Tenebrionidae (Coleoptera). Acta Oecol. 2012, 43, 22–28. [Google Scholar] [CrossRef]
- Fattorini, S. Testing the latitudinal gradient: A narrow-scale analysis of tenebrionid richness (Coleoptera, Tenebrionidae) in the Aegean archipelago (Greece). Ital. J. Zool. 2006, 73, 203–211. [Google Scholar] [CrossRef]
- Giraldo, A.E.; Flores, G.E. Peruvian Tenebrionidae: A review of present knowledge and biodiversity. Ann. Zool. 2016, 66, 499–513. [Google Scholar] [CrossRef]
- Kamiński, M.; Lumen, R.; Kanda, K.; Iwan, D.; Johnston, M.A.; Kergoat, G.; Bouchard, P.; Li, X.; Guo-dong, R.; Smith, A. Reevaluation of Blapimorpha and Opatrinae: Addressing a major phylogeny-classification gap in darkling beetles (Coleoptera: Tenebrionidae: Blaptinae). Syst. Entomol. 2021, 46, 140–156. [Google Scholar] [CrossRef]
- Mas-Peinado, P.; Buckley, D.; Ruiz, J.; García-París, M. Recurrent diversification patterns and taxonomic complexity in morphologically conservative ancient lineages of Pimelia (Coleoptera: Tenebrionidae). Syst. Entomol. 2018, 43, 522–548. [Google Scholar] [CrossRef]
- Soldati, L.; Condamine, F.L.; Clamens, A.L.; Kergoat, G.J. Documenting tenebrionid diversity: Progress on Blaps Fabricius (Coleoptera, Tenebrionidae, Tenebrioninae, Blaptini) systematics, with the description of five new species. Eur. J. Taxon. 2017, 282, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Junior, A.; Zucateli, S. Systematics and Phylogeny of Diaperinae (Coleoptera: Tenebrionidae); Universidade Federal de Viçosa: Viçosa, Brazil, 2020. [Google Scholar]
- Kergoat, G.J.; Soldati, L.; Clamens, A.L.; Jourdan, H.; Jabbour-Zahab, R.; Genson, G.; Bouchard, P.; Condamine, F.L. Higher level molecular phylogeny of darkling beetles (Coleoptera: Tenebrionidae). Syst. Entomol. 2014, 39, 486–499. [Google Scholar] [CrossRef]
- Aalbu, R.L.; Triplehorn, C.A.; Campbell, J.M.; Brown, K.W.; Somerby, R.E.; Thomas, D.B. Tenebrionidae Latreille. In American Beetles, Vol. 2. Polyphaga: Scarabaeoidea through Curculionoidea; Ross, H., Arnett, M.C., Thomas, P.E., Skelley, J., Howard, F., Eds.; CRC Press: New York, NY, USA, 2002; pp. 463–509. [Google Scholar]
- McCormack, J.E.; Hird, S.M.; Zellmer, A.J.; Carstens, B.C.; Brumfield, R.T. 11.14. Tenebrionidae Latreille, 1802. In Handbook of zoology. A Natural History of the Phyla of the Animal Kingdom, Vol. IV—Arthropoda: Insecta. Part 38. Coleoptera, Beetles, Vol. 2: Systematics (Part 2); Leschen, R.A.B., Beutel, R.G., John, J.F.L., Eds.; Walter de Gruyter: Berlin, Germany, 2010; pp. 574–659. [Google Scholar]
- Daly, H.V. Insect morphometrics. Annu. Rev. Entomol. 1985, 30, 415–438. [Google Scholar] [CrossRef]
- Yan, B.R.; Hua, B.Z. Geometric morphometrics and its application in the systematics and phylogenetics of insects. Entomotaxonomia 2010, 32, 313–320. [Google Scholar] [CrossRef]
- Bouchard, P.; Bousquet, Y.; Davies, A.; Alonso-Zarazaga, M.; Lawrence, J.; Lyal, C.; Newton, A.; Reid, C.; Schmitt, M.; Slipinski, A.; et al. Family-Group Names In Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.D.; Jing, N.; Jia, L. A new species of the genus Cheirodes Gen-From Alxa plateau (Coleoptera, Tenebrionidae, Melanimin). Acta Zoot. Sin. 2011, 36, 564–567. [Google Scholar]
- Ren, G.D. Six new species of the genus Uloma Gaoligong Mountain in China (Coleoptera, Tenebrionidae). Acta Zoot. Sin. 2004, 29, 296–304. [Google Scholar] [CrossRef]
- Yibin, B. Systematics of Pimeliinae and Its Distribution in China (Coleoptera: Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2012. [Google Scholar]
- Haoyu, L. Fauna and Distribution of the Darkling Beetles in Qinghai-Xizang Plateau (Coleoptera: Tenebrionoidea). Master’s Thesis, Hebei University, Baoding, China, 2010. [Google Scholar]
- Yong, Z. Taxonomy of Lagriina from China (ColeoPtera, Lagriini). Ph.D. Thesis, Hebei University, Baoding, China, 2011. [Google Scholar]
- Long, J. Fauna and Distribution of Tenebrionid Beetles in Alxa Plateau (Coleoptera: Tenebrionoidea). Ph.D. Thesis, Hebei University, Baoding, China, 2014. [Google Scholar]
- Ping, L. A Preliminary Study on the Taxonomy of the Tribe Stenochiini from Hainan Island (Coleoptera: Tenebrionidae). Ph.D. Thesis, Yanan University, Yan’an, China, 2019. [Google Scholar]
- Xiumin, L. Phylogenetic and Historical Biogeography of Partial Genera of the Tribe Blaptini from the Qinghai-Xizang Plateau. Ph.D. Thesis, Hebei University, Baoding, China, 2020. [Google Scholar]
- Xue, L. Classification of Stored Products Darkling Beetles in China (Coleoptera: Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2020. [Google Scholar]
- Li, L.; Yunzhen, L.; Aimin, S. A redescription of Bioramix aenescens (Blair, 1923) (Coleoptera: Tenbrionidae: Platyscelidini). J. Yunnan Agric. Univ. 2015, 30, 133–136. [Google Scholar]
- Xiaoli, L. Molecular Systematic Study on Partial Darkling Beetles Based on Sequences of Mitochondrial 16S rDNA and 18S rDNA. Master’s Thesis, Hebei University, Baoding, China, 2004. [Google Scholar]
- Chuanxiong, N. Taxonomy of Cteniopinus from China (Coleoptera: Tenebrionidae: Alleculinae). Master’s Thesis, Hebei University, Baoding, China, 2020. [Google Scholar]
- Yan, W.; Xiaoning, L.; Ji, M. Pigmentation patterns on larval cephalic capsule of three desert beetles (Coleoptera: Tentyriini). Chin. J. Appl. Entomol. 2011, 48, 447–451. [Google Scholar] [CrossRef] [Green Version]
- Xiaoqing, Y.; Guodong, R. A new species of the genus Derispia Lewis from Hainan, China (Coleoptera, Tenebrionidae, Leiochrinini). Acta Zoot. Sin. 2011, 36, 571–573. [Google Scholar]
- Min, Z.; Ying, F.; Xiaoming, C.; Huanhong, J. Morphological and biological characteristics of Blaps rhynchopetera (Coleoptera: Tenebrionidae) J. Environ. Entomol. 2009, 31, 348–355. [Google Scholar]
- Xiaolin, Z.; Guodong, R. A new species of the genus Laena Dejean from Xizang, China (Coleoptera, Tenebrionidae, Lagriinae). Acta Zoot. Sin. 2011, 3, 568–570. [Google Scholar]
- Yu, Z. Faunistic and Geographic Distribution on Tenebrionid Beetles of the Ordos Plateau (Coleoptera:Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2014. [Google Scholar]
- Ming, B. Systematic Study of the Subfamily Alleculinae from China (Coleoptera: Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2004. [Google Scholar]
- Shanshan, L.; Guodong, R.; Jiashe, W. Three new species of the genus Uloma Dejean, 1821 from Wuyi Mountain in China with a new record (Coleoptera, Tenebrionidae, Ulomini). Acta Zoot. Sin. 2007, 32, 70–75. [Google Scholar]
- Aloquio, S.; Lopes-Andrade, C. Redescription of immature stages and adults of Nilio (Nilio) brunneus (Coleoptera: Tenebrionidae: Nilioninae). Zoologia 2016, 33, e20150191. [Google Scholar] [CrossRef] [Green Version]
- Ando, K.; Ruzzier, E. New species of Tenebrionidae Latreille, 1802 (Coleoptera: Tenebrionoidea) from the Philippines. Zootaxa 2016, 4175, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Runraj, C.; Vineesh, P.; Sabu, T. Darkling beetles (Coleoptera: Tenebrionidae) of forest sites and agricultural fields in the south Western Ghats (South India). JIB 2017, 5, 1–12. [Google Scholar] [CrossRef]
- Xing-long, B.; Guo-dong, R. Revision of the genus Bioramix Bates, 1879 (Coleoptera: Tenebrionidae: Platyscelidini) from China. Zootaxa 2020, 4815, 1–102. [Google Scholar] [CrossRef]
- Xing-long, B.; Xiu-Min, L.; Guo-dong, R. A review of the genus Oodescelis Motschulsky, 1845 (Coleoptera: Tenebrionidae: Platyscelidini) from China. Zootaxa 2019, 4656, 401–430. [Google Scholar] [CrossRef]
- Bolaño, J.A.S.; Villero, J.M.B.; Puebla, P.B.d.l.; López-Colón, J.I. Aportación a la corología del género Prionychus Solier, 1835 (Coleoptera: Tenebrionidae, Alleculinae) en Espaa y Marruecos. Boletín SEA 2010, 229–234. [Google Scholar]
- Bousquet, Y.; Thomas, D.; Bouchard, P.; Smith, A.; Aalbu, R.; Johnston, M.A.; Steiner, W. Catalogue of Tenebrionidae (Coleoptera) of North America. ZooKeys 2018, 728, 1–455. [Google Scholar] [CrossRef] [PubMed]
- Cid-Arcos, M.; Campodonico, J. Nueva especie de Psectrascelis Solier (Coleoptera: Tenebrionidae) de la Region del Maule, Chile. Rev. Chil. Entomol. 2017, 43, 55–60. [Google Scholar]
- Doyen, J.T. A new genus and four new species of Coelometopini from MesoAmerica (Coleoptera: Tenebrionidae). Coleopts Bull. 1995, 49, 8–14. [Google Scholar] [CrossRef]
- Liebherr, J. Fauna of New Zealand (Ko te Aitanga Pepeke o Aotearoa) No. 43, Carabidae (Insecta: Coleoptera): Catalogue. J. New York Entomol. Soc. 2002, 110, 260–262. [Google Scholar] [CrossRef]
- Bujalance, J.; Cárdenas, A.; Ferrer, J. The genus Tentyria (Latreille, 1802) in Doñana national park (Southern Iberian Peninsula) with descriptions of two new species (Insecta: Coleoptera: Tenebrionidae). Ann. Zool. 2016, 66, 345–355. [Google Scholar] [CrossRef]
- Ferrer, J.; Ødegaard, F. New species of darkling beetles from Central America with systematic notes (Coleoptera: Tenebrionidae). Ann. Zool. 2005, 55, 633–661. [Google Scholar]
- Ferrer, J. The types of darkling beetles (Coleoptera: Tenebrionidae) described by Thunberg (1821, 1827) in "Coleoptera Capensia" and other papers, with taxonomic comments. Boletín SEA 2009, 44, 111–129. [Google Scholar]
- Witt, T. Das Weibchen von Anaea phoebe (Druce, 1877) (Lepidoptera, Nymphalidae). Munich Entomol Ges Mitt 1968, 59, 32–34. [Google Scholar]
- Grimm, R. New and little known species of Tenebrionidae (Coleoptera) from Borneo (6). Stuttgarter Beiträge Zur Nat. A 2016, 9, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Hopp, K.J.; Ivie, M.A. First report of Tyrtaeus rufus Champion and Tyrtaeus dobsoni Hinton (Tenebrionidae: Diaperinae: Anopidiina) from Florida. Coleopts Bull. 2008, 62, 427–436. [Google Scholar] [CrossRef]
- Kamiński, M.J.; Gearner, O.M.; Kanda, K.; Swichtenberg, K.; Purchart, L.; Smith, A.D. First insights into the phylogeny of tok-tokkie beetles (Tenebrionidae: Molurina, Phanerotomeina) and examination of the status of the Psammodes vialis species-group. Zool. J. Linn. Soc. 2020, 193, 889–901. [Google Scholar] [CrossRef]
- Kamiński, M.J.; Kanda, K.; Lumen, R.; Ulmer, J.M.; Wirth, C.C.; Bouchard, P.; Aalbu, R.; Mal, N.; Smith, A.D. A catalogue of the tribe Sepidiini Eschscholtz, 1829 (Tenebrionidae, Pimeliinae) of the world Launched to accelerate biodiversity research A peer-reviewed open-access journal. ZooKeys 2019, 844, 1–121. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Ren, G. Two new species and one newly recorded species of Uloma Dejean, 1821 from Zhejiang, China (Coleoptera, Tenebrionidae, Ulomini). Zookeys 2016, 607, 103–118. [Google Scholar] [CrossRef] [Green Version]
- Masumoto, K.; Akita, K.; Lee, C.F. New Tenebrionid (Coleoptera) beetles from Taiwan. Elytra 2012, 15, 193–205. [Google Scholar]
- Matthews, E.G.; Lawrence, J.F. Trachelostenini sensu novo: Redescriptions of Trachelostenus Solier, Myrmecodema Gebien and Leaus Matthews & Lawrence, based on adults and larvae, and descriptions of three new species of Leaus (Coleoptera: Tenebrionidae). Zootaxa 2015, 4020, 289–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkl, O. Redescription of Lagria (Apteronympha) tenenbaumi Pic, 1929, with a checklist of the Western Palaearctic species of the genus Lagria F. (Coleoptera: Tenebrionidae: Lagriini). Proc. Russ. Entomol. Soc. 2006, 77, 219–225. [Google Scholar]
- Pollard, R.M. An Assessment of the Polytypic Status of the Namib Darkling Beetles Onymacris Unguicularis and Onymacris Rugatipennis. Bachelor’s Thesis, East Carolina University, Greenville, NC, USA, 2014. [Google Scholar]
- Ruzzier, E.; Martínez-Muoz, C.A. First record of the invasive Lagria villosa (Fabricius, 1781) (Coleoptera: Tenebrionidae: Lagriinae) in Europe. Zootaxa 2021, 4908, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Schawaller, W. The Oriental species of Platydema Laporte & Brullé, part 2, with descriptions of 11 new species (Coleoptera: Tenebrionidae: Diaperinae). Stuttg. Beiträge Zur Nat. A 2012, 5, 243–255. [Google Scholar]
- Schawaller, W. New species of the genus Laena Dejean (Coleoptera: Tenebrionidae) from southeastern Asia (2). Ann. Zool. 2014, 64, 287–298. [Google Scholar] [CrossRef]
- Schawaller, W. New species of the genus Menimus Sharp (Coleoptera: Tenebrionidae: Gnathidiini) from Peninsular Malaysia and adjacent southern Thailand 1. Stuttg. Beiträge Zur Nat. A 2016, 9, 207–2016. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; Cifuentes-Ruiz, P. Revision of Diceroderes Solier (Coleoptera: Tenebrionidae: Toxicini: Eudysantina), with descriptions of four new species. Coleopts Bull. 2015, 69, 55–72. [Google Scholar] [CrossRef]
- Smith, A.D.; Wilson, J.S.; Cognato, A.I. The evolution of Batesian mimicry within the North American Asidini (Coleoptera: Tenebrionidae). Cladistics 2015, 31, 441–454. [Google Scholar] [CrossRef]
- Steiner, W.E.; Swearingen, J.M. New secords of three non-native darkling beetles (Coleoptera: Tenebrionidae) established in California and Nevada, USA. Coleopts Bull. 2015, 69, 22–26. [Google Scholar] [CrossRef]
- Triplehorn, C.A. A review of the genus Phaleria Latreille from the Western Hemisphere (Coleoptera: Tenebrionidae: Phaleriinae). Coleopts Bull. 1991, 45, 258–270. [Google Scholar] [CrossRef]
- Kamiński, M. Afrotropical Melambiina: Four New Species of Zadenos (Tenebrionidae: Pedinini), with Notes on Related Taxa. Ann. Zool. 2015, 65, 531–550. [Google Scholar] [CrossRef]
- Wenting, A. Systematics of the Tribe Platyopini in China (Coleoptera: Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2010. [Google Scholar]
- Ming, B.; Guodong, R. Four new species of the genus Cteniopinus Seidlite, 1896 from China (Coleoptera, Tenebrionidae, Alleculinae). Acta Zoot. Sin. 2003, 28, 682–687. [Google Scholar] [CrossRef]
- Xinglong, B. Systematic Taxonomy of Platyscelidini from China (Coleoptera: Tenebrionidae). Ph.D. Thesis, Hebei University, Baoding, China, 2020. [Google Scholar]
- Junsheng, S. Diversity and Particularity of the Darkling Beetles of the Everest Nature Reserve. Master’s Thesis, Hebei University, Baoding, China, 2016. [Google Scholar]
- Junsheng, S.; Guodong, R. Two New Species of the Genus Ascelosodis From Chinese Himalaya (Coleoptera, Tenebrionidae). J. Hebei Univ. 2016, 36, 175–179. [Google Scholar]
- Guodong, R. Two new species of the family Tenebrionidae from desert of northwest China (Coleoptera:Tenebrionidae Tenebrio). Zool. Res. 1992, 13, 330–332. [Google Scholar]
- Chao, G. Taxonomy of Part Diaperini from China (Coleoptera:Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2007. [Google Scholar]
- Xiujuan, Y. Systematic Study on the Tribe Opatrini in China (Coleoptera:Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2003. [Google Scholar]
- Huiran, H. A systematic Study on the Genus Promethis from China (Coleoptera:Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2006. [Google Scholar]
- Guodong, R. Three New Species of Genus Melanesthes Lacord. from China (Coleoptera,Tenebrionidae). Acta Entomol. Sin. 1993, 36, 486–489. [Google Scholar]
- Shanshan, L. Study on the Taxonomy of the Genus Uloma from China (Coleoptera: Tenebrionidae:Ulomini). Master’s Thesis, Hebei University, Baoding, China, 2006. [Google Scholar]
- Shanshan, L.; Guodong, R. Two new species of the genus Uloma Dejean,1821 from China (Coleoptera, Tenebrionidae, Ulomini). Acta Zoot. Sin. 2008, 33, 498–501. [Google Scholar]
- Shanshan, L.; Guodong, R. Taxonomic of the genus Uloma Dejean (Coleoptera, Tenebriondae, Ulomini) from Yunnan, China. Acta Zoot. Sin. 2013, 38, 559–565. [Google Scholar]
- Lei, M. Study on Taxonomy of the Tribe Platyscelidini from China (Coleoptera:Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2005. [Google Scholar]
- Lei, M.; Guodong, R. A systematic study on the genus Myatis Bates from China and adjacent areas (Coleoptera, Tenebrionidae). Acta Zoot. Sin. 2005, 30, 104–110. [Google Scholar]
- Akita, K.; Masumoto, K. New or little-known Tenebrionid beetles (Coleoptera, Tenebrionidae) from Japan (11) descriptions of seven new species,up-grading of a subspecies to the species rank, and a new distributional record of a species. Elytra 2011, 1, 275–294. [Google Scholar]
- Guodong, R.; Qiqi, W. Study on the Taxonomy of the Genus Toxicum from China (Coleoptera: Tenebrionidae:Toxicini). Acta Zoot. Sin. 2007, 32, 689–699. [Google Scholar]
- Guodong, R.; Chao, G. Study on the taxonomy of the genus Cerpria from China (Coleoptera: Tenebrionidae). Acta Zoot. Sin. 2007, 32, 200–207. [Google Scholar]
- Guodong, R.; Caixia, Y. Taxonomy of the genus Ainu Lewis, 1 894(Coleoptera: Tenebrionidae)from China, with description of a new specie. Entomotaxonomia 2015, 37, 129–133. [Google Scholar]
- Guodong, R.; Benyong, M. Resoure of darkling beetles from Yunnan and the species problems of Blaps in developing and ultilizing. In Proceedings of the National Symposium on Entomological Fauna Classification and Diversity, Kunming, China, 23–25 June 2005; pp. 176–182. [Google Scholar]
- Guodong, R.; Ximeng, W. Three new species of The Genus Adesmia Fisch. from China (Coleoptera: Tenebriondae, Adesmini). J. Ningxia Agric. Coll. 1993, 14, 1–8. [Google Scholar]
- Guodong, R.; Ximeng, W.; Feng, M. Two new species and one new record of Platyope F.-W. from China (Coleoptera: Tenebriondae). J. Ningxia Agric. Coll. 1993, 14, 44–49. [Google Scholar]
- Guodong, R.; Youzhi, Y.; Feng, M.; Hong, J. Study on the genus Melanesthes lacord. and description of its three new species and five new recordeds species (Coleoptera: Tenebriondae) in China. J. Ningxia Agric. Coll. 1993, 14, 24–33. [Google Scholar]
- Aimin, S. Taxonomic and Faunistic Studies on the family Tenebrionidae from Xizang (Coleoptera:Tenebrionoidae). Master’s Thesis, Hebei University, Baoding, China, 2006. [Google Scholar]
- Aimin, S.; Guodong, R. A new species of Itagonia Reitter,1887(Coleoptera: Tenebrionidae) from Xizang. J. China West Norm. Univ. 2007, 28, 176–178. [Google Scholar]
- Fengyan, W. Faunistic and Distribution Pattern Studies on Tenebrionidae of the North China Region (Coleoptera: Tenebrionidea). Master’s Thesis, Hebei University, Baoding, China, 2008. [Google Scholar]
- Zhonghua, W.; Guodong, R. Taxonomy of the genus Luprops from China, with description of the larva of Luprops horni (Coleoptera:Tenebrionidae). Entomotaxonomia 2019, 41, 299–312. [Google Scholar]
- Qiqi, W. Fauna and Biogeography of the Toxicini from China (Coleoptera, Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2009. [Google Scholar]
- Qiqi, W.; Guodong, R. A taxonomic study of the genus Cryphaeus Klug (Coleoptera,Tenebrionidae,Toxicini) from China with descriptions of four new species. Acta Entomol. Sin. 2008, 51, 1065–1076. [Google Scholar]
- Wei, W.; Zhaotian, F.; Renxin, H. Three new species of Platyope (Coleoptera: Tenebrionidae) from Xinjiang, China. Entomotaxonomia 2005, 27, 283–288. [Google Scholar]
- Juan, X. Taxonomic Study on the Genus Bioramix Bates from China (Coleoptera: Tenebrionoidea: Platyscelidini). Master’s Thesis, China West Normal University, Nanchong, China, 2017. [Google Scholar]
- Xiaoqing, Y. A taxonomic Study on Tribe Leiochrinini from China (Coleoptera: Tenebrionidae: Diaperinae). Master’s Thesis, Hebei University, Baoding, China, 2012. [Google Scholar]
- Caixia, Y. A preliminary Study on the Taxonomy of the Tribe Strongyliini from China (Coleoptera:Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2005. [Google Scholar]
- Caixia, Y.; Guodong, R. Study of the Genus Arboreal Darkling Beetles of the Strongylium from China (Coleoptera, Tenebrionidae). Acta Zoot. Sin. 2005, 30, 399–406. [Google Scholar]
- Caixia, Y.; Guodong, R. A new record genus of Strongyliini (Coleoptera, Tenebrionidae) from China, with two new species and two new names. Acta Zoot. Sin. 2006, 31, 851–854. [Google Scholar]
- Chengli, Z. Faunistic and Evolution on Tenebrionidae of the Arid and Semi-Arid Region of China. Master’s Thesis, Hebei University, Baoding, China, 2010. [Google Scholar]
- Qing, Z. Taxonomy of the Genus Eucyrtus and Its Five Related Genera from China (Coleoptera, Tenebrionidae, Stenochiinae). Master’s Thesis, Hebei University, Baoding, China, 2013. [Google Scholar]
- Xiaolin, Z. Taxonomic of Laena and Hypolaenopsis from China (Coleoptera, Tenebrionidae, Lagriinaeagriinae). Master’s Thesis, Hebei University, Baoding, China, 2012. [Google Scholar]
- Xichao, Z. Taxonomic and Faunistic Studies on the Family Tenebrionidae from Qinghai (Coleoptera: Tenebrionidae). Master’s Thesis, Hebei University, Baoding, China, 2014. [Google Scholar]
- BugGuide. Available online: https://bugguide.net/node/view/152/ (accessed on 23 April 2021).
- BOLD Systems (Barcode of Life Data System). Available online: http://v3.boldsystems.org/ or https://www.boldsystems.org/ (accessed on 28 May 2021).
- Flickriver. Available online: https://www.flickriver.com/ (accessed on 23 July 2021).
- BioLib.cz (Biological Library). Available online: https://www.biolib.cz/ (accessed on 2 June 2021).
- iNaturalist. Available online: https://inaturalist.ca/ (accessed on 1 July 2021).
- Virtual-beetle. Available online: http://www.virtual-beetle.com (accessed on 7 June 2021).
- MNHN (Museum National d’Histoire naturelle, Paris, France). Available online: https://science.mnhn.fr/ (accessed on 7 June 2021).
- TENEBRIONIDAE DE CHILE. Available online: http://www.coleoptera-neotropical.org/paginas/2_PAISES/Chile/teneb_ch.html (accessed on 14 May 2021).
- SCAN (Symbiota Collections of Arthropods Network). Available online: https://scan-bugs.org/portal/ (accessed on 3 July 2021).
- Zoological Institute of Russian Academy of Sciences. Available online: https://www.zin.ru/animalia/coleoptera/eng/atl_te.htm (accessed on 18 July 2021).
- Insecta.pro. Available online: http://insecta.pro/ (accessed on 18 July 2021).
- Museum Royal Belgian Institute of Natural Sciences. Available online: https://virtualcollections.naturalsciences.be/ (accessed on 17 July 2021).
- Ukrainian Biodiversity Information Network. Available online: https://ukrbin.com/ (accessed on 2 July 2021).
- Beetles of Africa. Available online: http://www.beetlesofafrica.com/ (accessed on 18 July 2021).
- TENEBRIONIDAE Latreille. 1802. Available online: http://www.tenebrionidae.net/ (accessed on 18 July 2021).
- Southern Alps Photography. Available online: https://www.southernalpsphotography.com/ (accessed on 1 July 2021).
- Wikispecies. Available online: https://species.wikimedia.org/wiki/Main_Page (accessed on 17 March 2021).
- Annals of the Transvaal Museum—BioNames. Available online: http://bionames.org/archive/issn/0255-0172/7/1.pdf (accessed on 11 June 2021).
- Bai, M.; Yang, X.K.; Jing, L.I.; Wang, W.C. Geometric Morphometrics, a super scientific computing tool in morphology comparison. Chin. Sci. Bull. 2014. [Google Scholar] [CrossRef]
- Rohlf, F.J. tpsDig, Digitize Landmarks and Outlines; Version 2.05; Department of Ecology and Evolution, State University of New York: New York, NY, USA, 2005. [Google Scholar]
- Rohlf, F.J. tps-SMALL. In Version 1.20 [Software and Manual]; Department of Ecology and Evolution, State University of New York: New York, USA, 2003. [Google Scholar]
- Goodall, C. Procrustes methods in the statistical analysis of shape. J. R. Stat. Soc. B 1991, 53, 285–321. [Google Scholar] [CrossRef]
- Macleod, N. PalaeoMath: Part 18-Form & Shape Models. Palaeontology 2009, 101, 72. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Green, S.B.; Salkind, N.J. Using SPSS for Windows and Macintosh, 7th ed.; Pearson: London, UK, 2013. [Google Scholar]
- Pretorius, E.; Scholtz, C.H. Geometric morphometries and the analysis of higher taxa: A case study based on the metendosternite of the Scarabaeoidea (Coleoptera). Biol. J. Linn. Soc. 2001, 74, 35–50. [Google Scholar] [CrossRef]
- Bai, M.; Jarvis, K.; Wang, S.-Y.; Song, K.-Q.; Wang, Y.-P.; Wang, Z.-L.; Li, W.-Z.; Wang, W.; Yang, X.-K. A second new species of ice crawlers from China (Insecta: Grylloblattodea), with thorax evolution and the prediction of potential distribution. PLoS ONE 2010, 5, e12850. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Foote, M. Morphological approaches to measuring biodiversity. Trends. Ecol. Evol. 1997, 12, 277–281. [Google Scholar] [CrossRef]
- DeLong, C.M.; Bragg, R.; Simmons, J.A. Evidence for spatial representation of object shape by echolocating bats (Eptesicus fuscus). J. Acoust. Soc. Am. 2008, 123, 4582–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.S. Pattern and process in competition. Adv. Ecol. Res. 1967, 4, 1–74. [Google Scholar] [CrossRef]
- Burns, J.; Strauss, S. More closely related species are more ecologically similar in an experimental test. Proc. Natl. Acad. Sci. USA 2011, 108, 5302–5307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, I.; Denno, R. Interspecific interactions in phytophagous insects revisited: A quantitative assessment of competition theory. Ecol. Lett. 2007, 10, 977–994. [Google Scholar] [CrossRef]
- Jenssen, T.A. Shift in the structural habitat of Anolis opalinus due to congeneric competition. Ecology 1973, 54, 863–869. [Google Scholar] [CrossRef]
- Boer, P.J. Exclusion or coexistence and the taxonomic or ecological relationship between species. Neth. J. Zool. 1979, 30, 278–306. [Google Scholar] [CrossRef]
- Fattorini, S.; Mantoni, C.; Di Biase, L.; Strona, G.; Pace, L.; Biondi, M. Elevational patterns of generic diversity in the tenebrionid beetles (Coleoptera Tenebrionidae) of latium (Central Italy). Diversity 2020, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Rundle, H.D.; Nosil, P. Ecological speciation. Ecol. Lett. 2005, 8, 336–352. [Google Scholar] [CrossRef]
- Schluter, D. Ecology and the origin of species. Trends Ecol. Evol. 2001, 16, 372–380. [Google Scholar] [CrossRef]
- Schluter, D. Evidence for ecological speciation and its alternative. Science 2009, 323, 737–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, R.L.; Sweeney, M.J.; Badyaev, A.V. Morphological diversity and ecological similarity: Versatility of muscular and skeletal morphologies enables ecological convergence in shrews. Funct. Ecol. 2010, 24, 556–565. [Google Scholar] [CrossRef]
- Gould, S. A developmental constraint in Cerion, with comments on the definition and interpretation of constraint in evolution. Evolution 1989, 43, 516–539. [Google Scholar] [CrossRef]
- Wainwright, P. Functional versus morphological diversity in macroevolution. Annu. Rev. Ecol. Evol. S 2007, 38, 381–401. [Google Scholar] [CrossRef]
- Guodong, R.; Guohong, W. Study on the role of darkling beetles’ sub-elytralcavity in sandy areas. Entomol. Knowl. 1990, 27, 160–162. [Google Scholar]
- Ren, G.D.; Yu, Y.Z.; Ma, F. Desert envIronment and adaptatIon of darklIng beetles. J. Ningxia Agric. Coll. 1993, 14, 85–92. [Google Scholar]
- Xianlei, S. Morphological Characteristics of HEALING-elytron and Its Relationship with Habitat Selection of the Darkling Beetles. Master’s Thesis, Hebei University, Baoding, China, 2019. [Google Scholar]
- Foote, M. The evolution of morphological diversity. Annu. Rev. Ecol. Evolut. Syst. 1997, 28, 129–152. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Guodong, R. Species diversity and faunal distribution of darkling beetles (Coleoptera, Tenebrionidae) on the Ordos Platean. J. Mong. Agric. Univ. 2014, 35, 19–25. [Google Scholar]
- Humphreys, A.; Barraclough, T. The evolutionary reality of higher taxa in mammals. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, T.S. The origin of higher taxa: Macroevolutionary processes, and the case of the mammals. Acta Zool. 2007, 88, 3–22. [Google Scholar] [CrossRef]
- OOliveira, S.; Ortega, J.C.; Ribas, L.; Lopes, V.; Bini, L. Higher taxa are sufficient to represent biodiversity patterns. Ecol. Indic. 2020, 111, 105994. [Google Scholar] [CrossRef]
- Timms, L.; Bowden, J.; Summerville, K.; Buddle, C. Does species-level resolution matter? Taxonomic sufficiency in terrestrial arthropod biodiversity studies. Insect Conserv. Divers. 2013, 6, 453–462. [Google Scholar] [CrossRef]
- Taravati, S.; Darvish, J.; Mirshamsi, O. Geometric morphometric study of twospecies of the psammophilous genusErodiontes (Coleoptera: Tenebrionidae) from the Lute desert, Central Iran. IJAB 2009, 5, 81–89. [Google Scholar] [CrossRef]
- Cruz, L.; Torres, M.A.; Barrion, A.; Joshi, R.; Sebastian, L.; Demayo, C. Geometric morphometric analysis of the head, pronotum and genitalia of the rice black bug associated with selected rice types. Egypt. Acad. J. Biol. Sci. A Entomol. 2011, 4, 21–31. [Google Scholar] [CrossRef]
- Robin, W. Functional morphology of insect wings. Annu. Rev. Entomol. 1992, 37, 113–140. [Google Scholar] [CrossRef]
- Evans, M.E.G. Locomotion in the Coleoptera Adephaga, especially Carabidae. J. Zool. 2010, 181, 189–226. [Google Scholar] [CrossRef]
- Snodgrass, R.E. Principles of Insect Morphology; Cornell University Press: New York, NY, USA, 1935. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Groups | Sampling Number of Tribes | Described Number of Tribes | Sampling Number of Genera | Described Number of Genera | Sampling Number of Species |
|---|---|---|---|---|---|
| Zolodininae | * | * | 2 | 3 | 2 |
| Lagriinae | 9 | 9 | 62 | 270 | 357 |
| Nilioninae | * | * | 1 | 1 | 21 |
| Phrenapatinae | 3 | 3 | 8 | 20 | 16 |
| Pimeliinae | 33 | 39 | 215 | 401 | 835 |
| Tenebrioninae | 29 | 29 | 220 | 955 | 1198 |
| Alleculinae | 2 | 2 | 54 | 140 | 317 |
| Diaperinae | 10 | 11 | 43 | 120 | 286 |
| Stenochiinae | 3 | 3 | 73 | 390 | 375 |
| Total | 89 | 96 | 678 | 2300 | 3407 |
| Test Groups | MD of Pronotum | MD of Elytron |
|---|---|---|
| Zolodininae | 0.0046 | 0.0036 |
| Lagriinae | 0.0100 | 0.0039 |
| Nilioninae | 0.0130 | 0.0014 |
| Phrenapatinae | 0.0074 | 0.0026 |
| Pimeliinae | 0.0267 | 0.0051 |
| Tenebrioninae | 0.0124 | 0.0041 |
| Alleculinae | 0.0111 | 0.0019 |
| Diaperinae | 0.0295 | 0.0066 |
| Stenochiinae | 0.0171 | 0.0042 |
| Tribes Richness | Genera Richness | Species Richness | ||||
|---|---|---|---|---|---|---|
| r | p | r | p | r | p | |
| MD of pronotum | 0.521 | 0.075 | 0.350 | 0.178 | 0.500 | 0.085 |
| MD of elytron | 0.782 | 0.006 | 0.600 | 0.044 | 0.571 | 0.077 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, L.; Tong, Y.; Zhao, Y.; Sun, Z.; Wang, X.; Ma, F.; Bai, M. Study on the Relationship between Richness and Morphological Diversity of Higher Taxa in the Darkling Beetles (Coleoptera: Tenebrionidae). Diversity 2022, 14, 60. https://doi.org/10.3390/d14010060
Cheng L, Tong Y, Zhao Y, Sun Z, Wang X, Ma F, Bai M. Study on the Relationship between Richness and Morphological Diversity of Higher Taxa in the Darkling Beetles (Coleoptera: Tenebrionidae). Diversity. 2022; 14(1):60. https://doi.org/10.3390/d14010060
Chicago/Turabian StyleCheng, Liangxue, Yijie Tong, Yuchen Zhao, Zhibin Sun, Xinpu Wang, Fangzhou Ma, and Ming Bai. 2022. "Study on the Relationship between Richness and Morphological Diversity of Higher Taxa in the Darkling Beetles (Coleoptera: Tenebrionidae)" Diversity 14, no. 1: 60. https://doi.org/10.3390/d14010060
APA StyleCheng, L., Tong, Y., Zhao, Y., Sun, Z., Wang, X., Ma, F., & Bai, M. (2022). Study on the Relationship between Richness and Morphological Diversity of Higher Taxa in the Darkling Beetles (Coleoptera: Tenebrionidae). Diversity, 14(1), 60. https://doi.org/10.3390/d14010060