

Morphological and Molecular Studies of Three New Diatom Species from Mountain Streams in South Korea



Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
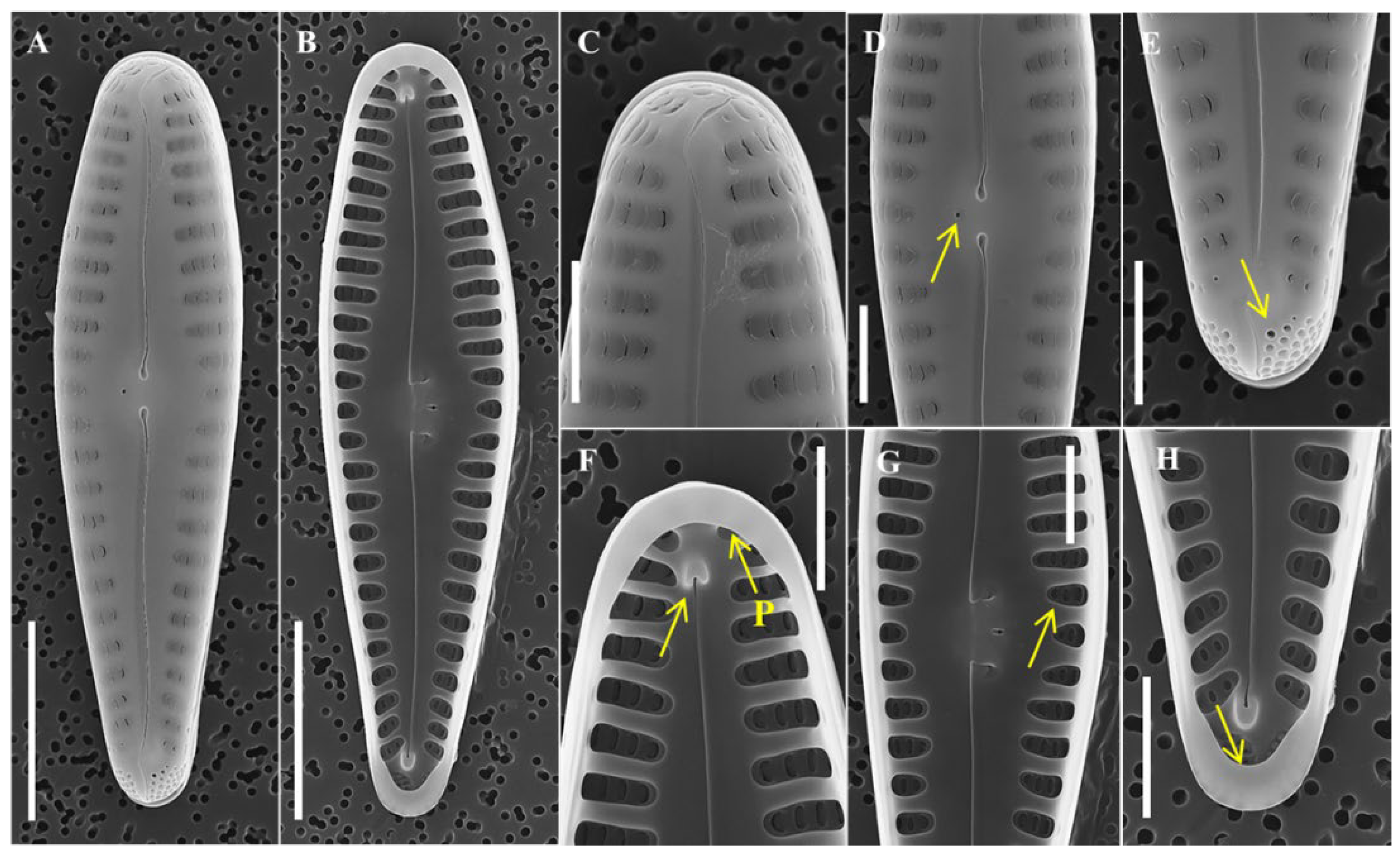
3.1. Morphological Characteristics of Hannaea librata sp. nov.
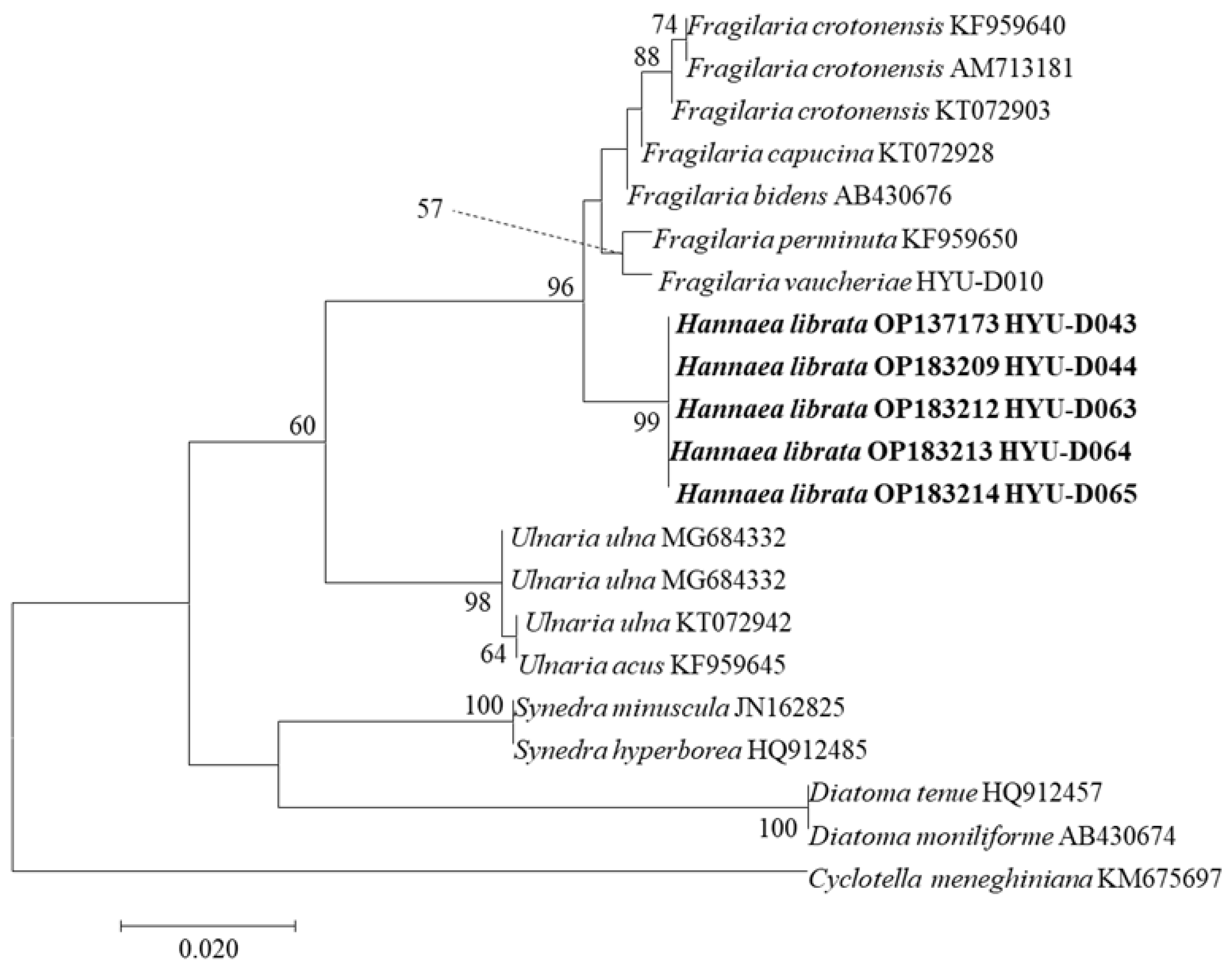
3.2. Phylogenetic Characteristics of H. librata sp. nov.
3.3. Morphological Characteristics of Gomphonema Seminulum sp. nov.
3.4. Phylogenetic Characteristics of Gomphonema Seminulum sp. nov.
3.5. Morphological Characteristics of Nitzschia inclinata sp. nov.
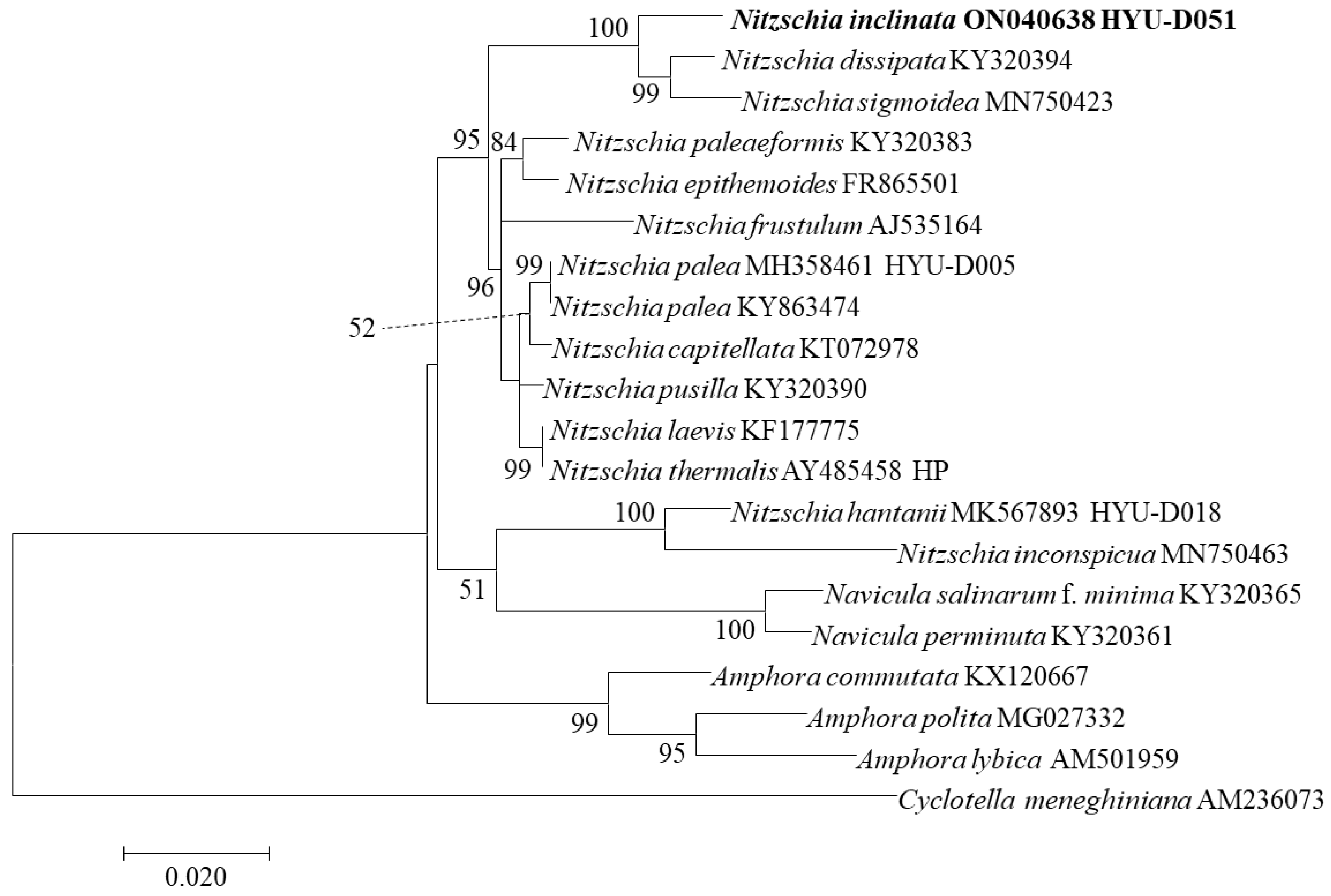
3.6. Phylogenetic Characteristics of Nitzschia inclinata sp. nov.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mann, D.G.; Vanormelingen, P. An inordinate fondness? The number, distributions, and origins of diatom species. J. Eukaryot. Microbiol. 2013, 60, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Kovács, C.; Kahlert, M.; Padisák, J. Benthic diatom communities along pH and TP gradients in Hungarian and Swedish streams. J. Appl. Phycol. 2006, 18, 105–117. [Google Scholar] [CrossRef]
- Pestryakova, L.A.; Herzschuh, U.; Gorodnichev, R.; Wetterich, S. The sensitivity of diatom taxa from Yakutian lakes (north-eastern Siberia) to electrical conductivity and other environmental variables. Polar Res. 2018, 37, 1485625. [Google Scholar] [CrossRef]
- Kim, H.K.; Cho, I.H.; Hwang, E.A.; Kim, Y.J.; Kim, B.H. Benthic diatom communities in Korean estuaries: Species appearances in relation to environmental variables. Int. J. Environ. Health Res. 2019, 16, 2681. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Lowe, R.L. Independent and interactive effects of nutrients and grazers on benthic algal community structure. Hydrobiologia 1994, 291, 201–209. [Google Scholar] [CrossRef]
- Pan, Y.; Stevenson, R.J.; Hill, B.H.; Herlihy, A.T.; Collins, G.B. Using diatoms as indicators of ecological conditions in lotic systems: A regional assessment. J. N. Am. Benthol. Soc. 1996, 15, 481–495. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; 747p. [Google Scholar]
- Green, J.C.; Leadbeater, B.S.C.; Diver, W.L. (Eds.) The origins of the diatom and its life cycle. In The Chromophyte Algae: Problems and Perspectives; Clarendon Press: Oxford, UK, 1989; pp. 305–321. [Google Scholar]
- Witkowski, A.; Sieminska, J. (Eds.) A review of the evolution of the diatoms—A total approach using molecules, morphology and geology. In The Origin and Early Evolution of the Diatoms: Fossil, Molecular and Biogeographical Approaches; Polish Academy of Sciences: Krakow, Poland, 2000; pp. 13–35. [Google Scholar]
- Cox, E.J. The identity and typification of some naviculoid diatoms (Bacillariophyta) from freshwater or brackish habitats. Phycologia 1998, 37, 162–175. [Google Scholar] [CrossRef]
- Morales, E.A.; Siver, P.A.; Trainor, F.R. Identification of diatoms (Bacillariophyceae) during ecological assessments: Comparison between light microscopy and scanning electron microscopy techniques. Proc. Acad. Nat. Sci. Phila. 2001, 151, 95–103. [Google Scholar] [CrossRef]
- Tan, L.; Wang, P.; Cho, I.H.; Hwang, E.A.; Lee, H.; Kim, B.H. Morphology and phylogenetic position of three new raphid diatoms (Bacillariophyceae) from Hangang River, South Korea. Phytotaxa 2020, 442, 153–182. [Google Scholar] [CrossRef]
- Auwera, G.V.D.; Wachter, R.D. Structure of the large subunit rDNA from a diatom, and comparison between small and large subunit ribosomal RNA for studying stramenopile evolution. J. Eukaryot. Microbiol. 1998, 45, 521–527. [Google Scholar] [CrossRef]
- Mann, D.G.; Simpson, G.E.; Sluiman, H.J.; Möller, M. rbcL gene tree of diatoms: A second large data-set for phylogenetic reconstruction. Phycologia 2001, 40, 1–2. [Google Scholar]
- Kim, H.P. A Study on Ecological Model to Estimate Water Quality in the Lake Pal’tang; Hanyang University: Seoul, Korea, 1999. [Google Scholar]
- Choi, H.J.; Cho, Y.C.; Yu, S.; Song, Y.S.; Ryu, I. Analysis of Water Circulation Characteristics for Hydraulic and Water Temperature Investigation in Paldang Reservoir. Ecol. Resilient Infrastruct. 2019, 6, 12–22. [Google Scholar]
- Tsoi, W.Y.; Hadwen, W.L.; Sheldon, F. Identifying diatom indicator species of nutrient enrichment: An in situ nutrient enrichment experiment in subtropical upland streams. Ecol. Indic. 2020, 119, 106744. [Google Scholar] [CrossRef]
- Andersen, R.A. (Ed.) Algal Culturing Techniques; Elsevier: San Diego, CA, USA, 2005; p. 578. [Google Scholar]
- Beakes, G.W.; Canter, H.M.; Jaworski, G.H.M. Zoospore ultrastructure of Zygorhizidium affluens and Z. planktonicum, two chytrids parasitizing the diatom Asterionella formosa. Can. J. Bot. 1988, 66, 1054–1067. [Google Scholar] [CrossRef]
- Miao, M.; Li, Z.; Hwang, E.A.; Kim, H.K.; Lee, H.; Kim, B.H. Two New Benthic Diatoms of the Genus Achnanthidium (Bacillariophyceae) from the Hangang River, Korea. Diversity 2020, 12, 285. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, P.; Kim, H.K.; Lee, H.; Han, M.S.; Kim, B.H. Lemnicola hungarica (Bacillariophyceae) and the new monoraphid diatom Lemnicola uniseriata sp. nov. (Bacillariophyceae) from South Korea. Diatom Res. 2018, 33, 69–87. [Google Scholar] [CrossRef]
- Ki, J.S.; Han, M.S. Molecular analysis of complete ssu to lsu rdna sequence in the harmful dinoflagellate Alexandrium tamarense (Korean isolate, HY970328M). Ocean Sci. 2005, 40, 43–54. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Bruder, K.; Medlin, L. Molecular assessment of phylogenetic relationships in selected species/genera in the naviculoid diatoms (Bacillariophyta). I. The genus Placoneis. Nova Hedwig. 2007, 85, 331. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7 (2016): Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Glushchenko, A.; Kulikovskiy, M.; Maltsev, Y.; Kociolek, J.P. New Hannaea Patrick (Fragilariaceae, Bacillariophyta) species from Asia, with comments on the biogeography of the genus. Cryptogam. Algol. 2019, 40, 41–61. [Google Scholar] [CrossRef]
- Meister, F. Beiträge zur Bacillariaceenflora Japan. Arch. Hydrobiol. 1914, 9, 226–232. [Google Scholar]
- Skvortzow, B.W. A contribution to the diatoms of Baikal Lake. Proc. R. Soc. B 1928, 1, 3. [Google Scholar]
- Kobayasi, H.; Idei, M.; Mayama, S.; Nagumon, T.; Osada, K. (Eds.) Kobayasi’s Atlas of Japanese Diatoms Based on Electron Microscopy; Unchida Rokakuho: Tokyo, Japan, 2006; p. 531. [Google Scholar]
- Bixby, R.J. Morphology, Phytogeography, and Systematics of the Diatom Genus Hannaea (Bacillariophyceae); University of Michigan: Ann Arbor, MI, USA, 2001. [Google Scholar]
- Williams, D.M. Tabulariopsis, a new genus of marine araphid diatom, with notes on the taxonomy of Tabularia (Kütz.) Williams et Round. Nova Hedwig. 1988, 47, 247–254. [Google Scholar]
- Lange-Bertalot, H. Zur systematischen Bewertung der bandformigen Kolonien von Navicula und Fragilaria. Kriterien fur die Vereinigung von Synedra (subgen. Synedra) Ehrenberg mit Fragilaria Lyngbye. Nova Hedwig. 1980, 33, 723–787. [Google Scholar]
- Tuji, A.; Williams, D.M. Typification of Conferva pectinalis OF Müll. (Bacillariophyceae) and the identity of the type of an alleged synonym, Fragilaria capucina Desm. Taxon 2006, 55, 193–199. [Google Scholar] [CrossRef]
- Heudre, D.; Wetzel, C.E.; Moreau, L.; Van de Vijver, B.; Ector, L. On the identity of the rare Fragilaria subconstricta (Fragilariaceae), with Fragilaria species forming ribbon-like colonies shortly reconsidered. Plant Ecol. Evol. 2019, 152, 327–339. [Google Scholar] [CrossRef]
- Reichardt, E. Taxonomische revision des artenkomplexes um Gomphonema pumilum (Bacillariophyceae). Nova Hedwig. 1997, 99–129. [Google Scholar] [CrossRef]
- Reichardt, E.; Lange-Bertalot, H. Taxonomische Revision des Artenkomplexes um Gomphonoma angustum—G. dichotomum—G. intricatum—G. vibrio und ähnliche Taxa (Bacillariophyceae). Nova Hedwig. 1991, 53, 519–544. [Google Scholar]
- Wojtal, A. Diatoms of the genus Gomphonema Ehr. (Bacillariophyceae) from a karstic stream in the Krakowsko-Czestochowska Upland. Acta Soc. Bot. Pol. 2003, 72, 213–220. [Google Scholar] [CrossRef]
- Agardh, C.A. (Ed.) Conspectus Criticus Diatomacearum. Part 3; Litteris Berlingianis: Lundae, Sweden, 1831; pp. 33–48. [Google Scholar]
- Fricke, F. (Ed.) Verzeichniss der in A. Schmidt’s Atlas der Diatomaceenkunde Tafel 1–240 (Serie IV) Abgebildeten und Bekannten Formen/Herausgegeben von Friedr; O.R. Reisland: Leipzig, Germany, 1902. [Google Scholar]
- Rabenhorst, L. (Ed.) Die Algen Sachsens. Resp. Mittel-Europa’s Gesammelt und Herausgegeben von Dr. L.; Exsiccata: Dresden, Germany, 1860. [Google Scholar]
- Lange-Bertalot, H. (Ed.) Indicators of Oligotrophy. 800 Taxa Representative of Three Ecologically Distinct Lake Types; Koeltz Scientific Books: Königstein, Germany, 1996; 390p. [Google Scholar]
- Knattrup, A.; Yde, M.; Lundholm, N.; Ellegaard, M. A detailed description of a Danish strain of Nitzschia sigmoidea, the type species of Nitzschia, providing a reference for future morphological and phylogenetic studies of the genus. Diatom Res. 2007, 22, 105–116. [Google Scholar] [CrossRef]
- Smith, W. (Ed.) A Synopsis of the British Diatomaceae; with Remarks on Their Structure, Function and Distribution; and Instructions for Collecting and Preserving Specimens; Paternoster Row: London, UK, 1853; Volume 1, pp. 1–89. [Google Scholar]
- Joh, G. (Ed.) Chrysophyta: Bacillariophyceae: Pennales: Raphidineae: Naviculaceae: Cymbella, Cymbopleura, Encyonema, Encyonopsis, Reimeria, Gomphonema. Freshwater diatoms IV. In Algal Flora of Korea; National Institute of Biological Resources: Incheon, Korea, 2011; Volume 3, pp. 1–70. [Google Scholar]
- Lange-Bertalot, H. (Ed.) Diatomeen im Süßwasser—Benthos von Mitteleuropa. Bestimmungsflora Kieselalgen für die Ökologische Praxis. Über 700 der Häufigsten Arten und Ihre Ökologie, Königstein; Koeltz Scientific Books: Königstein, Germany, 2013; pp. 1–908. [Google Scholar]
- In Diatoms of North America. Available online: https://diatoms.org/species/nitzschia_dissipata (accessed on 15 August 2022).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Nucleotide Sequence (5′ to 3′) | References |
|---|---|---|---|
| SSU rRNA | AT18F01 | YAC CTG GTT GAT CCT GCC AGT AG | [21] |
| AT18R02 | GTTTCAGCCTTGCGACCATACTCC | [21] | |
| AT18F02 | AGA ACG AAA GTT AAG GGA TCG AAG ACG | [21] | |
| AT18R01 | GCTTGATCCTTCTGCAGGTTCACC | [21] | |
| EulA | AAC CTG GTT GAT CCT GCC AGT | [22] | |
| EukB | GAT CCT TCT GCA GGT TCA CCT AC | [22] | |
| rbcL | F3 | GCT TAC CGT GTA GAT CCA GTT CC | [23] |
| R3 | CCT TCT AAT TTA CCA ACA ACT G | [23] |
| Hannaea librata | H. pamirensis | H. hattoriana | H. recta | Fragilaria capucina | |
|---|---|---|---|---|---|
| Length (μm) | 44–99 | 42–45 | 40–85 | 29–71 | 28–47 |
| Width (μm) | 5–5.5 | 5.5–7.0 | 6–7 | 6–7 | 3.3–4.2 |
| No. striae (/10 μm) | 12–16 | 17–18 | 13–14 | 12–14 | 14–17 |
| No. areolae (/10 μm) | 70–80 | 75–80 | n.d. | n.d. | n.d. |
| Valves | Linear | Weakly arcuate to almost linear | Lanceolate, slightly arcuate to almost linear | Lanceolate | Linear |
| Valve apices | Strongly rostrate | Capitate to rostrate apices | Capitate to rostrate apices | Rostrate | Weakly rostrate |
| Striation | Parallel | Parallel | Parallel | Parallel | |
| Ghost strain | Central area | Central area | Central area | Central area | n.d. |
| Valve face | Waved | Waved | n.d. | Waved | Alternate, parallel to slightly radiate toward the apices |
| Valve mantle | Flat | Flat | n.d. | Waved | n.d. |
| Valves of central area | Expanded to unilateral | Expanded to unilateral | Expanded to unilateral | Expanded to unilateral | Flat |
| Central area | Wide U-shape extending into both striae | Unilaterally tumid on the concave margin | Unilaterally tumid on the concave margin | Horseshoe shaped | Rectangular to rhombic |
| Girdle band | Single row of small punctuate | n.d. | n.d. | n.d. | n.d. |
| Apical pore fields | Rectangular | Rectangular | n.d. | Rectangular | n.d. |
| Spine shape | Conical | Bifurcated thorn (center)/ conical (near the pole) | n.d. | Bifurcated thorn (center)/ conical (near the pole) | Conical near the apex to triangular in the middle |
| Rimoportula | One per valve | One per valve | n.d. | One per valve | Two per valve |
| References | This study | [27] | [27,28] | [29,30] | [31,32,33,34,35] |
| Species | Accession | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Similarity | ||||||||||||||||
| 1 Hannaea librata | ON040635 | 1.000 | 1.000 | 1.000 | 1.000 | 0.989 | 0.989 | 0.989 | 0.987 | 0.987 | 0.985 | 0.974 | 0.974 | 0.969 | 0.969 | |
| 2 Hannaea librata | ON040636 | 0.000 | 1.000 | 1.000 | 1.000 | 0.989 | 0.989 | 0.989 | 0.987 | 0.987 | 0.985 | 0.974 | 0.974 | 0.969 | 0.969 | |
| 3 Hannaea librata | ON040640 | 0.000 | 0.000 | 1.000 | 1.000 | 0.989 | 0.989 | 0.989 | 0.987 | 0.987 | 0.985 | 0.974 | 0.974 | 0.969 | 0.969 | |
| 4 Hannaea librata | ON040641 | 0.000 | 0.000 | 0.000 | 1.000 | 0.989 | 0.989 | 0.989 | 0.987 | 0.987 | 0.985 | 0.974 | 0.974 | 0.969 | 0.969 | |
| 5 Hannaea librata | ON040642 | 0.000 | 0.000 | 0.000 | 0.000 | 0.989 | 0.989 | 0.989 | 0.987 | 0.987 | 0.985 | 0.974 | 0.974 | 0.969 | 0.969 | |
| 6 Fragilaria capucina | MH356257 | 0.011 | 0.011 | 0.011 | 0.011 | 0.011 | 0.999 | 1.000 | 0.996 | 0.996 | 0.994 | 0.979 | 0.979 | 0.965 | 0.965 | |
| 7 F. capucina var. mesolepta | MH997845 | 0.011 | 0.011 | 0.011 | 0.011 | 0.011 | 0.001 | 0.999 | 0.996 | 0.996 | 0.992 | 0.979 | 0.979 | 0.966 | 0.966 | |
| 8 Fragilaria bidens | AB430599 | 0.011 | 0.011 | 0.011 | 0.011 | 0.011 | 0.000 | 0.001 | 0.996 | 0.996 | 0.994 | 0.979 | 0.979 | 0.965 | 0.965 | |
| 9 Fragilaria crotonensis | AM712616 | 0.013 | 0.013 | 0.013 | 0.013 | 0.013 | 0.004 | 0.004 | 0.004 | 1.000 | 0.997 | 0.977 | 0.977 | 0.965 | 0.965 | |
| 10 Fragilaria vaucheriae | AM497733 | 0.013 | 0.013 | 0.013 | 0.013 | 0.013 | 0.004 | 0.004 | 0.004 | 0.000 | 0.997 | 0.977 | 0.977 | 0.965 | 0.965 | |
| 11 Fragilaria vaucheriae | EU260469 | 0.015 | 0.015 | 0.015 | 0.015 | 0.015 | 0.006 | 0.007 | 0.006 | 0.003 | 0.003 | 0.974 | 0.974 | 0.963 | 0.963 | |
| 12 Synedra minuscula | EF423415 | 0.027 | 0.027 | 0.027 | 0.027 | 0.027 | 0.022 | 0.022 | 0.022 | 0.024 | 0.024 | 0.026 | 1.000 | 0.959 | 0.959 | |
| 13 Synedra hyperborea | AY485464 | 0.027 | 0.027 | 0.027 | 0.027 | 0.027 | 0.022 | 0.022 | 0.022 | 0.024 | 0.024 | 0.026 | 0.001 | 0.959 | 0.959 | |
| 14 Ulnaria acus | KF959659 | 0.030 | 0.030 | 0.030 | 0.030 | 0.030 | 0.034 | 0.033 | 0.034 | 0.034 | 0.034 | 0.036 | 0.041 | 0.041 | 0.999 | |
| 15 Ulnaria ulna | MG684361 | 0.030 | 0.030 | 0.030 | 0.030 | 0.030 | 0.034 | 0.033 | 0.034 | 0.034 | 0.034 | 0.036 | 0.041 | 0.041 | 0.001 | |
| p-distance | ||||||||||||||||
| Species | Accession | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Similarity | ||||||||||||||||||
| 1 Hannaea librata | OP137173 | 1.000 | 1.000 | 1.000 | 1.000 | 0.988 | 0.980 | 0.986 | 0.982 | 0.984 | 0.984 | 0.982 | 0.914 | 0.914 | 0.947 | 0.945 | 0.947 | |
| 2 Hannaea librata | OP183209 | 0.000 | 1.000 | 1.000 | 1.000 | 0.988 | 0.980 | 0.986 | 0.982 | 0.984 | 0.984 | 0.982 | 0.914 | 0.914 | 0.947 | 0.945 | 0.947 | |
| 3 Hannaea librata | OP183212 | 0.000 | 0.000 | 1.000 | 1.000 | 0.988 | 0.980 | 0.986 | 0.982 | 0.984 | 0.984 | 0.982 | 0.914 | 0.914 | 0.947 | 0.945 | 0.947 | |
| 4 Hannaea librata | OP183213 | 0.000 | 0.000 | 0.000 | 1.000 | 0.988 | 0.980 | 0.986 | 0.982 | 0.984 | 0.984 | 0.982 | 0.914 | 0.914 | 0.947 | 0.945 | 0.947 | |
| 5 Hannaea librata | OP183214 | 0.000 | 0.000 | 0.000 | 0.000 | 0.988 | 0.980 | 0.986 | 0.982 | 0.984 | 0.984 | 0.982 | 0.914 | 0.914 | 0.947 | 0.945 | 0.947 | |
| 6 Fragilaria capucina | KT072928 | 0.012 | 0.012 | 0.012 | 0.012 | 0.012 | 0.986 | 0.998 | 0.994 | 0.996 | 0.990 | 0.988 | 0.921 | 0.921 | 0.945 | 0.943 | 0.945 | |
| 7 Fragilaria capucina | KC736594 | 0.020 | 0.020 | 0.020 | 0.020 | 0.020 | 0.014 | 0.988 | 0.984 | 0.986 | 0.988 | 0.986 | 0.921 | 0.921 | 0.945 | 0.943 | 0.945 | |
| 8 Fragilaria bidens | AB430676 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.002 | 0.012 | 0.992 | 0.994 | 0.992 | 0.990 | 0.923 | 0.923 | 0.947 | 0.945 | 0.947 | |
| 9 Fragilaria crotonensis | KF959640 | 0.018 | 0.018 | 0.018 | 0.018 | 0.018 | 0.006 | 0.016 | 0.008 | 0.998 | 0.988 | 0.986 | 0.917 | 0.917 | 0.941 | 0.938 | 0.941 | |
| 10 Fragilaria crotonensis | KT072903 | 0.016 | 0.016 | 0.016 | 0.016 | 0.016 | 0.004 | 0.014 | 0.006 | 0.002 | 0.990 | 0.988 | 0.919 | 0.919 | 0.943 | 0.941 | 0.943 | |
| 11 Fragilaria vaucheriae | HYUD010 | 0.016 | 0.016 | 0.016 | 0.016 | 0.016 | 0.010 | 0.012 | 0.008 | 0.012 | 0.010 | 0.994 | 0.923 | 0.923 | 0.947 | 0.945 | 0.947 | |
| 12 Fragilaria perminuta | KF959650 | 0.018 | 0.018 | 0.018 | 0.018 | 0.018 | 0.012 | 0.014 | 0.010 | 0.014 | 0.012 | 0.006 | 0.921 | 0.921 | 0.945 | 0.943 | 0.945 | |
| 13 Synedra minuscula | JN162825 | 0.081 | 0.081 | 0.081 | 0.081 | 0.081 | 0.075 | 0.075 | 0.073 | 0.079 | 0.077 | 0.073 | 0.075 | 1.000 | 0.934 | 0.936 | 0.934 | |
| 14 Synedra hyperborea | HQ912485 | 0.081 | 0.081 | 0.081 | 0.081 | 0.081 | 0.075 | 0.075 | 0.073 | 0.079 | 0.077 | 0.073 | 0.075 | 0.000 | 0.934 | 0.936 | 0.934 | |
| 15 Ulnaria acus | KF959645 | 0.051 | 0.051 | 0.051 | 0.051 | 0.051 | 0.053 | 0.053 | 0.051 | 0.057 | 0.055 | 0.051 | 0.053 | 0.063 | 0.063 | 0.998 | 1.000 | |
| 16 Ulnaria ulna | MG684332 | 0.053 | 0.053 | 0.053 | 0.053 | 0.053 | 0.055 | 0.055 | 0.053 | 0.059 | 0.057 | 0.053 | 0.055 | 0.061 | 0.061 | 0.002 | 0.998 | |
| 17 Ulnaria ulna | KT072942 | 0.051 | 0.051 | 0.051 | 0.051 | 0.051 | 0.053 | 0.053 | 0.051 | 0.057 | 0.055 | 0.051 | 0.053 | 0.063 | 0.063 | 0.000 | 0.002 | |
| p-distance | ||||||||||||||||||
| Gomphonema seminulum | G. bourbonense | G. pumilum | G. angustum | G. clevei | |
|---|---|---|---|---|---|
| Length (μm) | 15–20 | 9.4–28 | 17–37 | 13–130 | 11.5–34 |
| Width (μm) | 4–5 | 3.3–4.7 | 5–8 | 3–12 | 3.5–7 |
| No. striae (/10 μm) | 13–15 | 10.5–13 | 11–14 | 11–15 | 10–15 |
| No. areolae (/1 μm) | 4–5 | n.d. | n.d. | n.d. | 3–4 |
| Striae | Parallel to slightly radiate near the head pole, strongly radiate toward the foot pole | Slightly radiate, almost parallel | Slight radiate to strongly radiate toward the apices | Slightly radiate, parallel, low density of central area | Slightly radiate |
| Areola | Broadly C-shaped, occluded by raised flaps, 2–4 lines from the axis are flap open toward the center, and the other lines are open in the opposite direction | Occluded by raised flaps, C-shaped, slit-like | C-shaped, 1–3 lines from the axis are flap open toward the center, and the other lines are open in the opposite direction | n.d. | C-shaped, occluded by raised flaps, 2–3 lines from the axis are flap open toward the center, and the other lines are open in the opposite direction |
| Axial area | Wide lanceolate | Anguste lanceolate | Small | Wide rectangle | Wide lanceolate |
| Stigma | One per valve, circular | One per valve, circular | One per valve | One per valve | One per valve, circular with a thickened margin |
| Valves | Narrow lanceolate, slightly asymmetric about the axis | Linear and slightly elliptical | Narrow elliptic lanceolate, tapering more strongly toward the head pole | Narrow lanceolate–ovata | Rhombic clavate, elliptical clavate |
| Valve apices | Bluntly rounded | Obtuse, wide circular | bluntly rounded, slightly protracted apex | Wide circular | Obtuse (head pole is wider than foot pole) |
| Raphe | Extends along the entire valvar face, slightly undulate | Parallel | Extends along the entire valvar face | Extends along the entire valvar face | Undulate (external), relatively linear (internal) |
| Central raphe ending | Elliptical (external), bent to stigma, hooks in opposite directions (internal) | Elliptical(external), bent to stigma, hooks in opposite directions (internal) | Elliptical (external), sharp hook shape (internal) | Hook shape | Elliptical (external), bending to stigma, hook shape (internal) |
| Polar raphe ending | deflects in the opposite direction of the stigma (external), formed helictoglossa (internal) | n.d. | Deflects in the opposite direction of the stigma (external), formed helictoglossa (internal) | n.d. | Curved toward the stigma (external), formed small helictoglossa (internal) |
| Reference | This study | [36] | [36,37,38] | [37,38,39] | [30,40] |
| Species | Accession | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Similarity | ||||||||||||||||
| 1 Gomphonema seminulum | ON040637 | 0.992 | 0.992 | 0.992 | 0.991 | 0.992 | 0.992 | 0.992 | 0.991 | 0.991 | 0.976 | 0.976 | 0.974 | 0.974 | 0.969 | |
| 2 Gomphonema bourbonense | KC736621 | 0.007 | 1.000 | 1.000 | 0.999 | 0.990 | 0.992 | 0.992 | 0.991 | 0.992 | 0.974 | 0.974 | 0.972 | 0.972 | 0.968 | |
| 3 Gomphonema bourbonense | KC736620 | 0.007 | 0.000 | 1.000 | 0.999 | 0.990 | 0.992 | 0.992 | 0.991 | 0.992 | 0.974 | 0.974 | 0.972 | 0.972 | 0.968 | |
| 4 Gomphonema bourbonense | JN790281 | 0.007 | 0.000 | 0.000 | 0.999 | 0.990 | 0.992 | 0.992 | 0.991 | 0.992 | 0.974 | 0.974 | 0.972 | 0.972 | 0.968 | |
| 5 Gomphonema bourbonense | KT072957 | 0.009 | 0.001 | 0.001 | 0.001 | 0.989 | 0.990 | 0.990 | 0.990 | 0.991 | 0.972 | 0.972 | 0.972 | 0.972 | 0.967 | |
| 6 Gomphonema pumilum | KC736629 | 0.008 | 0.010 | 0.010 | 0.010 | 0.012 | 0.992 | 0.992 | 0.992 | 0.992 | 0.973 | 0.973 | 0.972 | 0.972 | 0.967 | |
| 7 Gomphonema acuminatum | JN790280 | 0.008 | 0.008 | 0.008 | 0.008 | 0.009 | 0.008 | 1.000 | 0.999 | 0.999 | 0.978 | 0.978 | 0.977 | 0.977 | 0.969 | |
| 8 Gomphonema acuminatum | AM502019 | 0.008 | 0.008 | 0.008 | 0.008 | 0.009 | 0.008 | 0.000 | 0.999 | 0.999 | 0.978 | 0.978 | 0.977 | 0.977 | 0.969 | |
| 9 Gomphonema subclavatum | KJ011666 | 0.009 | 0.009 | 0.009 | 0.009 | 0.010 | 0.009 | 0.001 | 0.001 | 0.999 | 0.977 | 0.977 | 0.976 | 0.976 | 0.969 | |
| 10 Gomphonema truncatum | AM501956 | 0.009 | 0.007 | 0.007 | 0.007 | 0.009 | 0.009 | 0.001 | 0.001 | 0.001 | 0.977 | 0.977 | 0.976 | 0.976 | 0.970 | |
| 11 Gomphonema parvulum | KC736625 | 0.024 | 0.026 | 0.026 | 0.026 | 0.027 | 0.027 | 0.022 | 0.022 | 0.022 | 0.022 | 1.000 | 0.997 | 0.998 | 0.985 | |
| 12 Gomphonema parvulum | KT072961 | 0.024 | 0.026 | 0.026 | 0.026 | 0.027 | 0.027 | 0.022 | 0.022 | 0.022 | 0.022 | 0.000 | 0.997 | 0.998 | 0.985 | |
| 13 Gomphonema dichotomum | KJ011655 | 0.025 | 0.027 | 0.027 | 0.027 | 0.028 | 0.028 | 0.023 | 0.023 | 0.024 | 0.024 | 0.003 | 0.003 | 0.998 | 0.985 | |
| 14 Gomphonema affine | MH997843 | 0.025 | 0.027 | 0.027 | 0.027 | 0.028 | 0.028 | 0.023 | 0.023 | 0.024 | 0.024 | 0.002 | 0.002 | 0.002 | 0.987 | |
| 15 Gomphonema clevei | KT072959 | 0.031 | 0.031 | 0.031 | 0.031 | 0.032 | 0.033 | 0.030 | 0.030 | 0.031 | 0.029 | 0.015 | 0.015 | 0.015 | 0.013 | |
| p-distance | ||||||||||||||||
| Species | Accession | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Similarity | |||||||||||||
| 1 Gomphonema seminulum | OP183210 | 0.955 | 0.949 | 0.962 | 0.962 | 0.960 | 0.953 | 0.942 | 0.942 | 0.948 | 0.948 | 0.927 | |
| 2 Gomphonema bourbonense | KC736595 | 0.044 | 0.947 | 0.962 | 0.958 | 0.953 | 0.957 | 0.948 | 0.948 | 0.942 | 0.942 | 0.926 | |
| 3 Gomphonema pumilum | KC736599 | 0.049 | 0.050 | 0.957 | 0.957 | 0.957 | 0.957 | 0.946 | 0.946 | 0.944 | 0.944 | 0.933 | |
| 4 Gomphonema acuminatum | LR742550 | 0.037 | 0.042 | 0.037 | 0.997 | 0.990 | 0.983 | 0.953 | 0.953 | 0.953 | 0.953 | 0.933 | |
| 5 Gomphonema acuminatum | LR742553 | 0.037 | 0.042 | 0.040 | 0.003 | 0.993 | 0.986 | 0.953 | 0.953 | 0.953 | 0.953 | 0.933 | |
| 6 Gomphonema subclavatum | LR742556 | 0.039 | 0.042 | 0.045 | 0.010 | 0.007 | 0.986 | 0.950 | 0.950 | 0.950 | 0.950 | 0.929 | |
| 7 Gomphonema truncatum | LR742547 | 0.045 | 0.042 | 0.042 | 0.017 | 0.013 | 0.013 | 0.950 | 0.950 | 0.949 | 0.949 | 0.933 | |
| 8 Gomphonema parvulum | KY661571 | 0.055 | 0.052 | 0.050 | 0.045 | 0.045 | 0.049 | 0.049 | 1.000 | 0.985 | 0.985 | 0.964 | |
| 9 Gomphonema parvulum | KY661554 | 0.055 | 0.052 | 0.050 | 0.045 | 0.045 | 0.049 | 0.049 | 0.000 | 0.985 | 0.985 | 0.964 | |
| 10 Gomphonema affine | AM710469 | 0.050 | 0.054 | 0.055 | 0.045 | 0.045 | 0.049 | 0.049 | 0.015 | 0.015 | 1.000 | 0.964 | |
| 11 Gomphonema affine | MK576042 | 0.050 | 0.054 | 0.055 | 0.045 | 0.045 | 0.049 | 0.049 | 0.015 | 0.015 | 0.000 | 0.964 | |
| 12 Gomphonema clevei | JQ354682 | 0.069 | 0.064 | 0.070 | 0.064 | 0.064 | 0.067 | 0.064 | 0.035 | 0.035 | 0.035 | 0.035 | |
| p-distance | |||||||||||||
| Nitzschia inclinata | N. dissipata | N. oligotraphenta | N. sigmoidea | N. angularis | |
|---|---|---|---|---|---|
| Length (μm) | 32–34.5 | 12.5–85 | 30–45 | 346–359 | 60–200 |
| Width (μm) | 3.5–5 | 3.5–7 | 3–4 | 9–13 | 6–15 |
| No. striae (/10 μm) | 50 | 39–50 | 46–48 | 24–26 | 31–32 |
| No. areolae (/10 μm) | 60–80 | 69 | n.d. | 40–50 | n.d. |
| No. fibulae (/10 μm) | 7–10 | 5–11.5 | 8.5–11.5 | 6–9 | 2.5–5 |
| Fibulae | One-sided valve, irregularly distributed | One side of valve, irregularly distributed | One-sided valve | One side of valve margin, irregularly distributed | Center of valve longitudinally striated |
| Striation | Parallel | Parallel | Parallel | Parallel | Parallel |
| Helictoglossa | Very apex of the valve (raphe ends internally formed) | n.d. | n.d. | Very apex of the valve (raphe ends internally formed) | Very apex of the valve |
| Valves | Narrow linear | Narrow lanceolate | Linear lanceolate | Linear | Rhomboidal |
| Valve apices | Capitate, bended | subrostrate | Distinctly capitulum | Tapering | n.d. |
| Raphe | Eccentric | Slightly eccentric | Eccentric | Highly eccentric, one side of valve margin | Almost central of valve face |
| Terminal fissure | Broad sweeping curve over the valve apex | Hook shape, sometimes widened or bifurcate | n.d. | Hook-shaped | Hook-shaped |
| Girdle view | Rectangular | n.d. | n.d. | Sigmoid | n.d. |
| Girdle band | Double rows of small punctuate | Double rows of small punctuate | n.d. | Double rows of small punctuate | n.d. |
| Reference | This study | [41,45,46,47] | [42,46,47] | [43] | [44] |
| Species | Accession | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Similarity | |||||||||||||||
| 1 Nitzschia inclinata | ON040638 | 0.984 | 0.981 | 0.978 | 0.976 | 0.976 | 0.975 | 0.975 | 0.974 | 0.975 | 0.975 | 0.963 | 0.953 | 0.937 | |
| 2 Nitzschia dissipata | KY320394 | 0.016 | 0.988 | 0.977 | 0.976 | 0.976 | 0.974 | 0.974 | 0.976 | 0.976 | 0.974 | 0.964 | 0.958 | 0.943 | |
| 3 Nitzschia sigmoidea | MN750423 | 0.019 | 0.012 | 0.975 | 0.972 | 0.972 | 0.974 | 0.974 | 0.971 | 0.972 | 0.970 | 0.959 | 0.953 | 0.938 | |
| 4 Nitzschia pusilla | KY320390 | 0.022 | 0.022 | 0.025 | 0.993 | 0.993 | 0.994 | 0.994 | 0.989 | 0.989 | 0.993 | 0.981 | 0.960 | 0.948 | |
| 5 Nitzschia palea | MH358461 | 0.024 | 0.024 | 0.027 | 0.006 | 1.000 | 0.995 | 0.995 | 0.991 | 0.990 | 0.995 | 0.982 | 0.962 | 0.951 | |
| 6 Nitzschia palea | KY863474 | 0.024 | 0.024 | 0.027 | 0.006 | 0.000 | 0.995 | 0.995 | 0.991 | 0.990 | 0.995 | 0.982 | 0.962 | 0.951 | |
| 7 Nitzschia laevis | KF177775 | 0.025 | 0.025 | 0.026 | 0.006 | 0.005 | 0.005 | 1.000 | 0.990 | 0.991 | 0.993 | 0.982 | 0.960 | 0.948 | |
| 8 Nitzschia thermalis | AY485458 | 0.025 | 0.025 | 0.026 | 0.006 | 0.005 | 0.005 | 0.000 | 0.990 | 0.991 | 0.993 | 0.982 | 0.960 | 0.948 | |
| 9 Nitzschia epithemoides | FR865501 | 0.025 | 0.024 | 0.028 | 0.011 | 0.009 | 0.009 | 0.010 | 0.010 | 0.993 | 0.990 | 0.979 | 0.958 | 0.949 | |
| 10 Nitzschia paleaeformis | KY320383 | 0.025 | 0.024 | 0.027 | 0.011 | 0.010 | 0.010 | 0.009 | 0.009 | 0.007 | 0.988 | 0.979 | 0.961 | 0.948 | |
| 11 Nitzschia capitellata | KT072978 | 0.025 | 0.025 | 0.029 | 0.006 | 0.005 | 0.005 | 0.006 | 0.006 | 0.010 | 0.012 | 0.980 | 0.961 | 0.948 | |
| 12 Nitzschia frustulum | AJ535164 | 0.038 | 0.037 | 0.041 | 0.020 | 0.019 | 0.019 | 0.019 | 0.019 | 0.022 | 0.022 | 0.021 | 0.949 | 0.936 | |
| 13 Nitzschia hantanii | MK567893 | 0.045 | 0.041 | 0.045 | 0.039 | 0.037 | 0.037 | 0.039 | 0.039 | 0.040 | 0.038 | 0.038 | 0.051 | 0.968 | |
| 14 Nitzschia inconspicua | MN750463 | 0.061 | 0.055 | 0.059 | 0.051 | 0.048 | 0.048 | 0.051 | 0.051 | 0.049 | 0.050 | 0.050 | 0.063 | 0.031 | |
| p-distance | |||||||||||||||
| Species | Accession | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Similarity | |||||||||||||
| 1 Nitzschia inclinata | OP183211 | 0.956 | 0.973 | 0.967 | 0.890 | 0.886 | 0.882 | 0.882 | 0.900 | 0.886 | 0.884 | 0.873 | |
| 2 Nitzschia dissipata | KY320332 | 0.043 | 0.948 | 0.956 | 0.867 | 0.871 | 0.871 | 0.871 | 0.900 | 0.873 | 0.871 | 0.860 | |
| 3 Nitzschia sigmoidea | FN557033 | 0.027 | 0.050 | 0.965 | 0.892 | 0.882 | 0.877 | 0.877 | 0.902 | 0.886 | 0.879 | 0.875 | |
| 4 Nitzschia cf. sigmoidea | KM999113 | 0.032 | 0.043 | 0.034 | 0.890 | 0.890 | 0.881 | 0.881 | 0.912 | 0.896 | 0.888 | 0.871 | |
| 5 Nitzschia capitellata | KT072924 | 0.102 | 0.122 | 0.101 | 0.102 | 0.976 | 0.965 | 0.965 | 0.912 | 0.926 | 0.928 | 0.916 | |
| 6 Nitzschia palea | KC736609 | 0.106 | 0.118 | 0.110 | 0.102 | 0.023 | 0.989 | 0.989 | 0.914 | 0.912 | 0.925 | 0.914 | |
| 7 Nitzschia palea | KF959639 | 0.110 | 0.118 | 0.113 | 0.110 | 0.034 | 0.011 | 1.000 | 0.906 | 0.904 | 0.917 | 0.906 | |
| 8 Nitzschia palea | KJ542464 | 0.110 | 0.118 | 0.113 | 0.110 | 0.034 | 0.011 | 0.000 | 0.906 | 0.904 | 0.917 | 0.906 | |
| 9 Nitzschia paleaeformis | KY320322 | 0.093 | 0.093 | 0.092 | 0.083 | 0.083 | 0.081 | 0.088 | 0.088 | 0.930 | 0.940 | 0.928 | |
| 10 Nitzschia hantanii | MK576040 | 0.106 | 0.117 | 0.106 | 0.097 | 0.070 | 0.083 | 0.090 | 0.090 | 0.066 | 0.961 | 0.950 | |
| 11 Nitzschia inconspicua | KC736607 | 0.108 | 0.118 | 0.111 | 0.104 | 0.068 | 0.072 | 0.079 | 0.079 | 0.057 | 0.038 | 0.969 | |
| 12 Nitzschia frustulum | HF675070 | 0.117 | 0.127 | 0.115 | 0.118 | 0.079 | 0.081 | 0.088 | 0.088 | 0.068 | 0.048 | 0.031 | |
| p-distance | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, E.-A.; Kim, H.-K.; Cho, I.-H.; Yi, C.; Kim, B.-H. Morphological and Molecular Studies of Three New Diatom Species from Mountain Streams in South Korea. Diversity 2022, 14, 790. https://doi.org/10.3390/d14100790
Hwang E-A, Kim H-K, Cho I-H, Yi C, Kim B-H. Morphological and Molecular Studies of Three New Diatom Species from Mountain Streams in South Korea. Diversity. 2022; 14(10):790. https://doi.org/10.3390/d14100790
Chicago/Turabian StyleHwang, Eun-A, Ha-Kyung Kim, In-Hwan Cho, Chen Yi, and Baik-Ho Kim. 2022. "Morphological and Molecular Studies of Three New Diatom Species from Mountain Streams in South Korea" Diversity 14, no. 10: 790. https://doi.org/10.3390/d14100790
APA StyleHwang, E. -A., Kim, H. -K., Cho, I. -H., Yi, C., & Kim, B. -H. (2022). Morphological and Molecular Studies of Three New Diatom Species from Mountain Streams in South Korea. Diversity, 14(10), 790. https://doi.org/10.3390/d14100790